猪miR-124靶向IQGAP2调节巨噬细胞内沙门氏菌的增殖
2020-03-24陈旺邓榆殷俊官州金凯石博妹黄廷华姚敏
陈旺 邓榆 殷俊 官州 金凯 石博妹 黄廷华 姚敏

摘要:【目的】明確猪miR-124与其靶基因IQGAP2间的表达调控关系,以及miR-124表达水平与猪巨噬细胞内沙门氏菌数量的关联,为揭示沙门氏菌在感染细胞内存活与增殖的机制提供理论依据。【方法】通过荧光素酶报告基因系统验证miR-124与IQGAP2基因的作用位点;再以GM-CSF诱导的猪巨噬细胞和鼠伤寒沙门氏菌(ATCC 14028)为试验材料,通过实时荧光定量PCR和流式细胞术测定沙门氏菌感染猪巨噬细胞中miR-124和IQGAP2基因的表达及巨噬细胞内沙门氏菌的增殖情况。【结果】miR-124结合位点野生型载体转染的荧光报告信号显著低于miR-124结合位点突变载体(P<0.05,下同),但共转染anti-miR-124序列后能显著增强miR-124结合位点野生型载体的荧光报告信号。经沙门氏菌感染后,猪巨噬细胞中的miR-124表达被激活,感染12、24和48 h后的相对表达量均显著高于沙门氏菌感染前(0 h),而IQGAP2基因表达水平呈显著下调趋势;在沙门氏菌感染猪巨噬细胞内,miR-124表达水平与IQGAP2基因表达水平呈明显负相关(r=-0.92)。miR-124高表达组细胞内的沙门氏菌数量显著高于正常巨噬细胞,但miR-124敲低表达组细胞内的沙门氏菌数量显著低于正常巨噬细胞;IQGAP2基因敲低表达组细胞内的沙门氏菌数量显著高于正常巨噬细胞;此外,miR-124高表达+IQGAP2基因敲低表达组细胞内的沙门氏菌数量与IQGAP2基因敲低表达处理组相比无显著差异(P>0.05),但显著高于miR-124高表达组细胞。【结论】沙门氏菌感染猪巨噬细胞中的miR-124表达水平与IQGAP2基因表达水平及胞内沙门氏菌数量呈负相关,即沙门氏菌可通过上调miR-124表达靶向抑制IQGAP2基因表达,从而调节其在猪巨噬细胞内的增殖。
关键词: 猪沙门氏菌;miR-124;IQGAP2基因;胞内增殖;流式细胞术;荧光素酶报告基因系统
中图分类号: S852.61 文献标志码: A 文章编号:2095-1191(2020)12-3066-07
Abstract:【Objective】To explore the regulation relationship between miR-124 and its target gene IQGAP2, study the relationship between the expression level of miR-124 and the number of Salmonella in pig macrophages,so as to provide a theoretical basis for revealing the mechanism of survival and proliferation of Salmonella in infected cells. 【Method】The binding sitebetween miR-124 and IQGAP2 was verified by luciferase reporter gene system assay. Then, the GM-CSF induced macrophages and Salmonella typhimurium(ATCC 14028) were used as research materials. The expression of miR-124 and IQGAP2 gene and the proliferation of Salmonella in macrophages were determined by real-time quantitative PCR and flow cytometry. 【Result】The fluorescence signal of wild-type vector transfected by miR-124 binding site was significantly lower than that of miR-124 binding site mutant vector(P<0.05, the same below), and the co-transfection of anti-miR-124 sequence could significantly enhance the fluorescence signal of wild-type vector. In the process of Salmonella infection, the expression of miR-124 was activated,the relative expression levels at 12, 24 and 48 h after infection were significantly higher than those before infection (0 h). The expression of IQGAP2 was significantly down-regulated. In Salmonella infected macrophages, the expression levels of miR-124 were negatively correlated with IQGAP2 (r=-0.92). The Salmonella counts in miR-124 high expression group were significantly higher than the control. The Salmonella counts in miR-124 knock down group were significantly lower than the control macrophages. The Salmonella counts in miR-124 knock-down group were significantly higher than the control macrophagesgroup. The numberof Salmonella counts in miR-124 high expression+IQGAP2 gene knockdown expression group were not significantly different compared to the IQGAP2 knockdown group(P>0.05), but significantly higher than the miR-124 high expression group cells. 【Conclusion】The expression levels of miR-124 and IQGAP2 in Salmonella infected porcine macrophages are negatively correlated with intercellular Salmonella counts. Salmonella inhibits IQGAP2 gene expression by up-regulating miR-124 expression targeting, thereby regulating its proliferation in porcine macrophages.
Key words: porcine Salmonella; miR-124; IQGAP2 gene; intracellular proliferation; flow cytometry; luciferase reporter gene system
Foundation item: National Natural Science Foundation of China(31902231,31402055); Innovation and Entrepreneurship Program of Yangtze University (2018057)
0 引言
【研究意义】沙门氏菌(Salmonella)感染猪群后通常形成急性一过性反应和长期携带2种状态(Alban and St?rk,2005;Bonardi,2017),即携带沙门氏菌生猪是沙门氏菌传播感染的重要媒介(Guan and Holley,2003;李帆等,2018)。沙门氏菌是一种常见的人畜共患病原菌(施开创等,2018),在我国的动物源性食物中毒事件中有70%~80%是由沙门氏菌污染猪肉产品而引起(杨怀珍等,2016);沙门氏菌在感染细胞(主要为吞噬细胞)内存活并实现胞内增殖是其形成长期携带及传播致病的重要原因(Lathrop et al.,2015)。因此,研究沙门氏菌在感染细胞内的存活和增殖发生机理及揭示相关基因在该过程中的调控作用,可为猪沙门氏菌感染的抗病育种提供分子标记,对防控生猪养殖生产中的沙门氏菌感染具有重要意义。【前人研究进展】本课题组前期通过高通量测序分析发现,在沙门氏菌感染仔猪外周血中miR-124、miR-16、miR-155和miR-143等29个miRNA呈显著差异表达,其中miR-124在沙门氏菌感染仔猪外周血中呈显著上调表达(5.86倍)(Huang et al.,2019),暗示miR-124是沙门氏菌感染过程中的重要调节因子。miR-124在免疫器官和免疫细胞,如外周血单核细胞、骨髓、淋巴结和胸腺组织中高表达,故推测其广泛参与机体的免疫调节过程(Smerkova et al.,2015)。miR-124可被脂多糖(LPS)和牛分枝杆菌卡介苗诱导,其转录水平受TLR信号适配分子MyD88调控(Mehta and Baltimore,2016;Sun et al.,2016)。激活后的miR-124通过阻碍TLR6、MyD88、TNF-α、USP2、USP14、P65、TRAF6及STAT3而反作用于(抑制)TLR信号通路(Sun et al.,2013;Ma et al.,2014;Qiu et al.,2015),其在基因组中存在多个拷贝,分别为miR-124-1、miR-124-2和miR-124-3,虽然三者的前体序列有所不同,但其成熟序列在人类、小鼠和猪中完全一致,说明miR-124在不同物种体内发挥着相同的生物学功能。沙门氏菌感染仔猪外周血miRNA/mRNA表达谱数据联合分析发现,在miR-124作用的110个候选靶基因中,沙门氏菌感染信号通路相关基因IQGAP2下调5倍,但同家族的IQGAP1和IQGAP3基因均未见显著差异表达(Huang et al.,2019)。IQGAP2基因通过与CDC42-GTP相结合并调节CDC42的活化状态(Brill et al.,1996),活化的CDC42进一步激活MAPK信号通路(Chen et al.,1996;Hobbie et al.,1997;Patel and Galán,2006),进而调节宿主的免疫功能(Garrett et al.,2000;Rodriguez-Escudero et al.,2011;Lathrop et al.,2015)。【本研究切入点】miR-124是沙门氏菌感染过程中的重要调节因子,是沙门氏菌在宿主体内存活、建立携带状态及感染致病的关键因素,但至今有关miR-124对下游靶基因IQGAP2的调节作用及其对细胞内沙门氏菌增殖情况的影响机制尚未明确。【拟解决的关键问题】采用实时荧光定量PCR、流式细胞术及荧光素酶报告检测系统对沙门氏菌感染猪巨噬细胞中miR-124和IQGAP2基因的表达及巨噬细胞内沙门氏菌的增殖情况进行探讨,为揭示沙门氏菌在感染细胞内的存活与增殖机制提供理论依据。
1 材料与方法
1. 1 试验材料
胎牛血清(10100154)和细胞培养液RPMI 1640(22400105)购自美国Gibco公司;哺乳动物外周血单核细胞分离液Percoll(P1644)购自Sigma-Aldrich公司;重组猪GM-CSF(ab233683)购自英国Abcam公司;总RNA提取试剂盒(R1200)购自北京索莱宝科技有限公司;miR-124定量检测试剂盒(480901_mir)购自美国ABI公司;兔抗人IQGAP2多克隆抗体(PA5-95484)和PE标记的羊抗兔IgG(31864)购自ThermoFisher公司;FITC标记的羊抗人CD14抗体(ABIN2478467)购自北京四正柏生物科技有限公司;动物细胞转染试剂TransFastTM(E2431)购自美国Promega公司;IQGAP2基因siRNA序列、miR-124序列、anti-miR-124序列及實时荧光定量PCR扩增引物(表1)均委托生工生物工程(上海)股份有限公司合成;psiCHECKTM-1、pGL3-Control、鼠伤寒沙门氏菌(ATCC 14028)和Raw264.7细胞由长江大学动物科学学院分子生物学实验室保存提供。
1. 2 miR-124高表达或敲低表达巨噬细胞的建立
以无菌生理盐水对仔猪外周抗凝血进行1∶1稀释,再使用哺乳动物外周血单核细胞分离液Percoll分离外周血单核细胞,并转移至细胞6孔培养板中,经贴壁、洗涤后,以GM-CSF(20 ng/mL)和IL-4(10 ng/mL)刺激培养6 d,以获得充分分化的巨噬细胞。将IQGAP2 siRNA(10 nmol/L)、miR-124 mimics(10 nmol/L)和anti-miR-124(10 nmol/L)以TransFastTM转染试剂包裹后分别转染分化的巨噬细胞,传染24 h后使用实时荧光定量PCR检测细胞中IQGAP2基因和miR-124的转录水平,以证实各组细胞建模是否成功。将对数生长期的沙门氏菌与构建的各组模型细胞按1∶1比例互作侵染,分别于感染0、12、24和48 h时收集细胞样品用于提取总RNA,设3次重复。采用TRIzol法提取总RNA,利用Stem-Loop TaqMan检测总RNA中的miR-124表达情况,以SYBR-Green PCR Master Mix试剂盒检测总RNA中的IQGAP2基因表达水平。用4%甲醛重悬细胞并在室温(20~25 ℃)下固定15 min,1×PBS离心洗涤。向预冷的细胞中缓慢加入预冷的100%甲醇,至甲醇终浓度为90%,温和涡旋以通透细胞。BD-PharmingenTM染色缓冲液以抗CD14-FITC抗体进行表面表型染色30 min,然后将通透处理的细胞重悬于BD PharmingenTM染色缓冲液中,加入anti-IQGAP2抗体,室温下孵育20 min。以100 ?L稀释的羊抗兔IgG偶联PE二抗再次重悬细胞,并室温孵育30 min,最后通过BD-LSRFortessaTM流式细胞分析仪进行分析。每组细胞处理均设3次重复,流式细胞术数据提交至流库数据库(Spidlen et al.,2012),编号FR-FCM-Z2FQ。
1. 3 胞内沙门氏菌增殖模型
参考Huang等(2018,2019)的方法以沙门氏菌侵染巨噬细胞。正常巨噬细胞、miR-124高表达及敲低表达巨噬细胞、IQGAP2基因敲低表达巨噬细胞、miR-124高表达+IQGAP2基因敲低表达巨噬细胞及miR-124敲低表达+IQGAP2基因敲低表达巨噬细胞经PBS洗涤和重悬后,分别按1×106/孔的密度接种至细胞6孔培养板中,同时按1∶1比例将对数生长期的沙门氏菌接种至各组巨噬细胞中,细胞培养板500 r/min离心10 min,使细胞和细菌聚集于培养底面,在37 ℃、5% CO2培养箱中孵育2 h,PBS漂洗3次,加入Gentamicin培养基(100 ?g/mL)继续培养2 h,再使用无抗生素培养基继续培养一定时间后收集细胞样品。自沙门氏菌与巨噬细胞互作时开始计时,分别收集4、8和12 h时的细胞样品,各3份。采用1% Tritone裂解破碎巨噬细胞,抽提DNA,使用实时荧光定量PCR检测沙门氏菌数量(Vinayaka et al.,2019)。
1. 4 双萤光素酶报告分析
将含有miR-124结合位点(5189~5195 nt,XM_021084462)的IQGAP2基因3'-UTR序列(XM_0210 84462,4761~5860 nt)野生型或突变(5192C/G)片段克隆至psiCHECKTM-1载体质粒海肾萤光素酶ORF下游,分别命名为psiCHECKTM-1-IQGAP2 3'-UTR(Wild-type)和psiCHECKTM-1-IQGAP2 3'-UTR(Mutant)。将RAW264.7细胞以每孔0.5×106个细胞接种至细胞6孔培养板中,当有60%细胞融合时采用TransFastTM转染试剂包裹miR-124(或anti-miR-124)、psiCHECKTM-1-IQGAP2 3'-UTR(Wild-type)[或psiCHECKTM-1-IQGAP2 3'-UTR(Mutant)],转染Raw264.7细胞。试验共设4组:(1)Mutant;(2)Mutant+miR-124;(3)Wilde type;(4)Wilde type+anti-miR-124。所有細胞均与转染效率对照质粒pGL3-Control共转染。每转染4 h,即采用双萤光素酶报告分析系统测量萤光素酶活性1次(Huang et al.,2018)。每组重复3次。
1. 5 统计分析
试验数据采用SPSS 18.0进行统计分析,其中,两组间比较采用 t 检验,多组间比较采用单因素方差分析(One-way ANOVA)。
2 结果与分析
2. 1 双萤光素酶报告解析miR-124对IQGAP2基因的调控作用
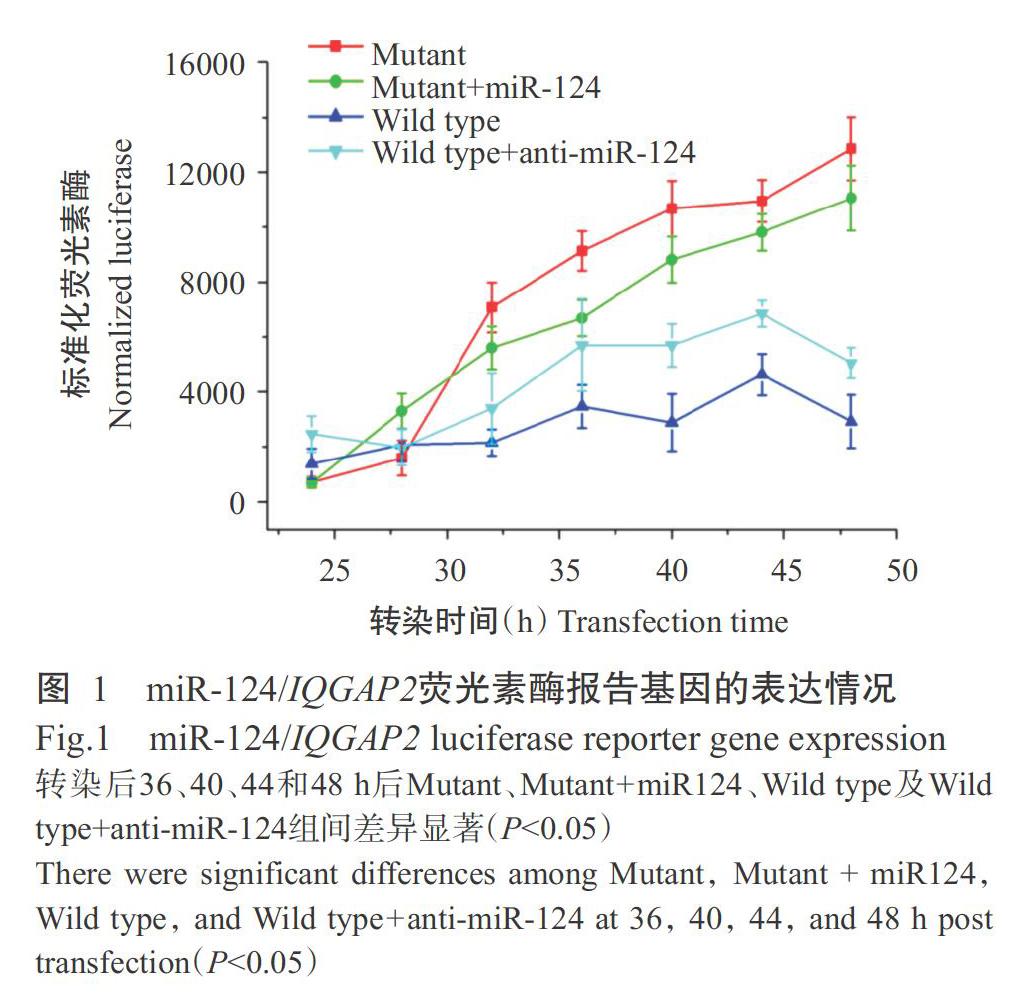
将IQGAP2基因3'-UTR区插入荧光素酶基因3'-UTR区,构建靶基因3'-UTR区的荧光素酶报告基因载体,以构建好的荧光素酶报告基因载体转染Raw264.7细胞,通过检测荧光素酶的表达情况分析IQGAP2基因3'-UTR区是否含有miR-124靶位点;同时构建miR-124结合位点人工点突变的报告基因载体。通过比较野生型和突变型报告基因的荧光素酶活性(图1)发现,转染36~48 h后miR-124结合位点野生型载体转染的荧光报告信号显著低于miR-124结合位点突变载体(P<0.05,下同),但共转染anti-miR-124序列后能显著增强miR-124结合位点野生型载体的荧光报告信号;miR-124结合位点突变载体转染及anti-miR-124序列与miR-124结合位点突变载体共转染均显著增强报告基因表达,说明miR-124能作用于IQGAP2基因3'-UTR区而抑制报告基因表达。
2. 2 沙门氏菌感染猪巨噬细胞中miR-124和IQGAP2基因的转录水平
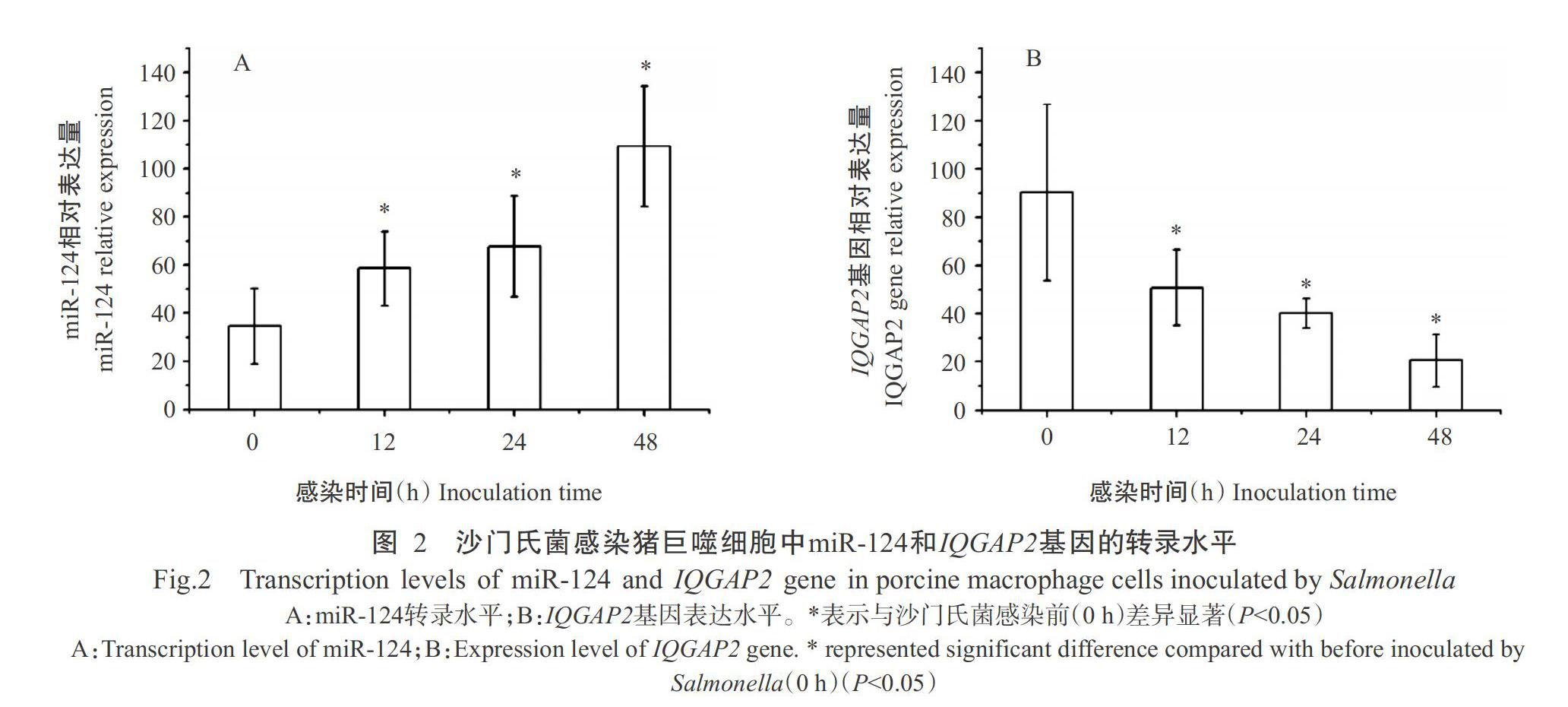
经沙门氏菌感染后,猪巨噬细胞中的miR-124表达水平明显升高(图2-A),感染12、24和48 h后的相对表达量均显著高于沙门氏菌感染前(0 h);IQGAP2基因表达水平则明显下调(图2-B),感染12、24和48 h的相对表达量显著低于沙门氏菌感染前(0 h)。在沙门氏菌感染猪巨噬细胞内,miR-124表达水平与IQGAP2基因表达水平呈明显负相关(r=-0.92)。
2. 3 沙门氏菌感染猪巨噬细胞中IQGAP2蛋白的流式细胞术检测结果
经沙门氏菌感染后,猪巨噬细胞中的IQGAP2蛋白表达水平呈明显下调趋势。在沙门氏菌感染前(0 h)约有61.1%的IQGAP2阳性细胞,沙门氏菌感染12 h后约有43.6%的IQGAP2阳性细胞,感染24 h后约有38.2%的IQGAP2阳性细胞,感染48 h后仅有23.0%的IQGAP2阳性细胞。即沙门氏菌感染12和24 h后的IQGAP2阳性细胞约是沙门氏菌感染前(0 h)的2/3,感染48 h后的IQGAP2阳性细胞约为沙门氏菌感染前(0 h)的1/3。流式细胞术检测结果见图3。
2. 4 沙门氏菌感染猪巨噬细胞内沙门氏菌数量的增殖规律
在沙门氏菌感染4 h后, miR-124高表达组细胞内的沙门氏菌数量明显高于正常巨噬细胞(约1.5倍),IQGAP2基因敲低表达组细胞内的沙门氏菌数量则是正常巨噬细胞的3.5倍。至沙门氏菌感染8 h后,miR-124高表达组和IQGAP2基因敲低表达组细胞内的沙门氏菌数量持续上升,miR-124高表达组细胞内的沙门氏菌数量明显高于正常巨噬细胞(约1.8倍),IQGAP2基因敲低表达组细胞内的沙门氏菌数量是正常巨噬细胞的4.0倍。在沙门氏菌感染12 h后,各处理组的胞内沙门氏菌数量均较感染8 h时有所上升,miR-124高表达组细胞内的沙门氏菌数量明显高于正常巨噬细胞(约2.0倍),IQGAP2基因敲低表达组细胞内的沙门氏菌数量是正常巨噬细胞的4.5倍。各感染时间点,miR-124敲低表达组胞内沙门氏菌数量均显著低于正常巨噬细胞;miR-124高表达+IQGAP2基因敲低表达组细胞内的沙门氏菌数量与IQGAP2基因敲低表达处理组相比无显著差异(P>0.05,下同),但显著高于miR-124高表达组细胞;miR-124敲低表达+IQGAP2基因敲低表达组细胞内的沙门氏菌数量与IQGAP2基因敲低表达处理组相比也无显著差异,但显著高于miR-124敲低表达组细胞、低于miR-124高表达+IQGAP2基因敲低表达组细胞。
3 讨论
沙门氏菌通过粪口途径进入动物体内,部分酸耐受菌体从胃进入小肠,穿过小肠黏膜层而作用于黏膜下层的M细胞(Prouty et al.,2004),再转运至肠系膜淋巴结(Chen et al.,2015)。沙门氏菌进入动物体内后,主要通过以下3条途径与机体免疫系统相互作用:(1)来源于沙门氏菌的LPS在MD2和CD14辅助下能被TLR4受体识别,TLR4识别沙门氏菌后可激活巨噬细胞并作用于TLR4适配分子MyD88,从而激活下游TLR4信号通路和NFκB信号通路(Mastroeni et al.,2009;Deng et al.,2016);(2)沙门氏菌通过III型分泌系统将菌体蛋白分泌到动物细胞内,直接(SopE和SopE2)或间接(SopB)作用于CDC42,并进一步激活MAPK信号通路(Chen et al.,1996;Hobbie et al.,1997;Patel and Galán,2006);(3)沙门氏菌基因组DNA可被动物细胞的外源DNA识别蛋白(DAI、AIM2和RIG-I)所识别,进而激活细胞质DNA感知信号通路(Qiu et al.,2015;Mehta and Baltimore,2016)。已有的研究虽然从不同角度阐明沙门氏菌对动物机体免疫系统的激活机制,但仍然无法合理解释沙门氏菌免疫逃避甚至携带排菌的现象。
miR-124在动物免疫器官和免疫细胞,如外周血单核细胞、骨髓、淋巴结和胸腺中高表达,并广泛参与机体免疫调节过程(Smerkova et al.,2015)。本研究结果表明,CSF诱导猪巨噬细胞经沙门氏菌感染后,miR-124呈上调表达,其上调表达机制可能与前人的相关研究结论(Mehta and Baltimore,2016;Sun et al.,2016)一致,即受沙门氏菌LPS-TLR信号的激活。IQGAP2基因的表达受甲基化和转录后水平等多种因素影响(Jin et al.,2008;Deng et al.,2016;Pelossof et al.,2016)。本研究的熒光素酶报告分析系统检测结果显示,IQGAP2基因3'-UTR区存在miR-124作用靶位点,一定程度上揭示了沙门氏菌感染仔猪巨噬细胞中miR-124与IQGAP2基因表达的负调控关系。miR-124高表达及IQGAP2基因敲低表达组猪巨噬细胞内沙门氏菌数量显著高于miR-124敲低表达组细胞,暗示miR-124可影响猪巨噬细胞内的沙门氏菌数量,其机制可能是通过作用于IQGAP2信号而影响宿主细胞对抗原的吞噬和加工能力,从而更有利于胞内沙门氏菌的存活与增殖。因此,miR-124/IQGAP2路径可能是沙门氏菌在宿主体内存活、增殖及建立携带状态的重要因素,是沙门氏菌激活宿主免疫的刹车系统,但具体作用机制尚需进一步研究证实。
4 结论
沙门氏菌感染猪巨噬细胞中的miR-124表达水平与IQGAP2基因表达水平及胞内沙门氏菌数量呈负相关,即沙门氏菌可通过上调miR-124表达靶向抑制IQGAP2基因表达,从而调节其在猪巨噬细胞内的增殖。
参考文献:
李帆,罗行炜,刘建华,梁军,贺丹丹,潘玉善,苑丽,胡功政. 2018. 猪源沙门氏菌多药外排泵oqxAB和氟苯尼考耐药基因floR的检测分析[J]. 江西农业学报,30(11):82-85. [Li F,Luo X W,Liu J H,Liang J,He D D,Pan Y S,Yuan L,Hu G Z. 2018. Detection and analysis of multi-drug efflux pump oqxAB and florfenicol-tolerant gene floR in swine-derived Salmonella typhimurium[J]. Acta Agriculturae Jiangxi,30(11):82-85.]
施开创,尹彦文,温丽霞,屈素洁,王海清,胡杰. 2018. 黏菌素耐药基因mcr-1 TaqMan-MGB荧光定量PCR检测方法的建立[J]. 南方农业学报,49(7):1447-1452. [Shi K C,Yin Y W,Wen L X,Qu S J,Wang H Q,Hu J. 2018. Establishment of TaqMan-MGB fluorescent quantitative PCR for detection of colistin resistance gene mcr-1[J]. Journal of Southern Agriculture,49(7):1447-1452.]
杨怀珍,牟亚,罗薇. 2016. 食源性沙门氏菌的研究进展[J]. 黑龙江畜牧兽医,(7):69-71. [Yang H Z,Mou Y,Luo W. 2016. Research progress of food-borne Salmonella[J]. Heilongjiang Animal Science and Veterinary Medicine,(7):69-71.]
Alban L,St?rk K D C. 2005. Where should the effort be put to reduce the Salmonella prevalence in the slaughtered swine carcass effectively?[J]. Preventive Veterinary Medi-cine,68(1):63-79.
Bonardi S. 2017. Salmonella in the pork production chain and its impact on human health in the European Union[J]. Epidemiology and Infection,145(8):1513-1526.
Brill S,Li S,Lyman C W,Church D M,Wasmuth J J,Weissbach L,Bernards A,Snijders A J. 1996. The Ras GTPase-activating-protein-related human protein IQGAP2 harbors a potential actin binding domain and interacts with calmo-dulin and Rho family GTPases[J]. Molecular and Cellular Biology,16(9):4869-4878.
Chen J,Tian J,Tang X Y,Rui K,Ma J,Mao C M,Liu Y Z,Lu L W,Xu H X,Wang S J. 2015. miR-346 regulates CD4+CXCR5+ T cells in the pathogenesis of Graves? disease[J]. Endocrine,49(3):752-760.
Chen L M,Hobbie S,Galán J E. 1996. Requirement of CDC42 for Salmonella-induced cytoskeletal and nuclear responses[J]. Science,274(5295):2115-2118.
Deng Z,Wang L J,Hou H L,Zhou J C,Li X. 2016. Epigenetic regulation of IQGAP2 promotes ovarian cancer progression via activating Wnt/beta-catenin signaling[J]. International Journal of Oncology,48(1):153-160.
Garrett W S,Chen L M,Kroschewski R,Ebersold M,Turley S,Trombetta S,Galán J E,Mellman I. 2000. Developmental control of endocytosis in dendritic cells by Cdc42[J]. Cell,102(3):325-334.
Guan T Y,Holley R A. 2003. Pathogen survival in swine manure environments and transmission of human enteric illness—A review[J]. Journal of Environmental Quality,32(2):383-392.
Hobbie S,Chen L M,Davis R J,Galán J E. 1997. Involvement of mitogen-activated protein kinase pathways in the nuclear responses and cytokine production induced by Salmonella typhimurium in cultured intestinal epithelial cells[J]. Journal of Immunology,159(11):5550-5559.
Huang T H,Huang X L,Chen W,Yin J,Shi B M,Wang F F,Feng W Z,Yao M. 2019. microRNA responses associa-ted with Salmonella enterica serovar typhimurium challenge in peripheral blood:Effects of miR-146a and IFN-gamma in regulation of fecal bacteria shedding counts in pig[J]. BMC Veterinary Research,15(1):195. doi:10.1186/ s12917-019-1951-4.
Huang T H,Huang X L,Yao M. 2018. miR-143 inhibits intracellular Salmonella growth by targeting ATP6V1A in macrophage cells in pig[J]. Research in Veterinary Science,117:138-143.
Jin S H,Akiyama Y,Fukamachi H,Yanagihara K,Akashi T,Yuasa Y. 2008. IQGAP2 inactivation through aberrant promoter methylation and promotion of invasion in gastric cancer cells[J]. International Journal of Cancer,122(5):1040-1046.
Lathrop S K,Binder K A,Starr T,Cooper K G,Chong A,Carmody A B,Steele-Mortimer O. 2015. Replication of Salmonella enterica serovar Typhimurium in human monocyte-derived macrophages[J]. Infection and Immunity,83(7):2661-2671.
Ma C Y,Li Y,Li M,Deng G C,Wu X L,Zeng J,Hao X J,Wang X P,Liu J,Cho W C S,Liu X M,Wang Y J. 2014. microRNA-124 negatively regulates TLR signaling in alveolar macrophages in response to mycobacterial infection[J]. Molecular Immunology,62(1):150-158.
Mastroeni P,Grant A,Restif O,Maskell D. 2009. A dynamic view of the spread and intracellular distribution of Salmonella enterica[J]. Nature Reviews. Immunology,7(1):73-80.
Mehta A,Baltimore D. 2016. microRNAs as regulatory elements in immune system logic[J]. Nature Reviews. Immunology,16(5):279-294.
Patel J C,Galán J E. 2006. Differential activation and function of Rho GTPases during Salmonella-host cell interactions[J]. The Journal of Cell Biology,175(3):453-463.
Pelossof R,Chow O S,Fairchild L,Smith J J,Setty M,Chen C T,Chen Z B,Egawa F,Avila K,Leslie C S,Garcia-Aguilar J. 2016. Integrated genomic profiling identifies microRNA-92a regulation of IQGAP2 in locally advanced rectal cancer[J]. Genes,Chromosomes & Cancer,55(4):311-321.
Prouty A M,Brodsky I E,Falkow S,Gunn J S. 2004. Bile-salt-mediated induction of antimicrobial and bile resistance in Salmonella typhimurium[J]. Microbiology(Rea-ding),150(Pt 4):775-783.
Qiu S W,Feng Y M,LeSage G,Zhang Y,Stuart C,He L,Li Y,Caudle Y,Peng Y,Yin D L. 2015. Chronic morphine-induced microRNA-124 promotes microglial immunosuppression by modulating P65 and TRAF6[J]. Journal of Immunology,194(3):1021-1030.
Rodriguez-Escudero I,Ferrer N L,Rotger R,Cid V J,Molina M. 2011. Interaction of the Salmonella typhimurium effector protein SopB with host cell Cdc42 is involved in intracellular replication[J]. Molecular Microbiology,80(5):1220-1240.
Smerkova K,Hudcova K,Vlahova V,Vaculovicova M,Peka-rik V,Masarik M,Adam V,Kizek R. 2015. Label-free and amplification-free miR-124 detection in human cells[J]. International Journal of Oncology,46(2):871-877.
Spidlen J,Breuer K,Rosenberg C,Kotecha N,Brinkman R R. 2012. FlowRepository:A resource of annotated flow cytometry datasets associated with peer-reviewed publications[J]. Cytometry (Part A),81(9):727-731.
Sun Y,Li Q,Gui H,Xu D P,Yang Y L,Su D F,Liu X. 2013. microRNA-124 mediates the cholinergic anti-inflammatory action through inhibiting the production of pro-inflammatory cytokines[J]. Cell Research,23(11):1270-1283.
Sun Y,Qin Z,Li Q,Wan J J,Cheng M H,Wang P Y,Su D F,Yu J G,Liu X. 2016. microRNA-124 negatively regulates LPS-induced TNF-alpha production in mouse macrophages by decreasing protein stability[J]. Acta Pharmacologica Sincia,37(7):889-897.
Vinayaka A C,Ngo T A,Kant K,Engelsmann P,Dave V P,Shahbazi M A,Wolff A,Bang D D. 2019. Rapid detection of Salmonella enterica in food samples by a novel approach with combination of sample concentration and direct PCR[J]. Biosensors & Bioelectronics,129:224-230.
(責任编辑 兰宗宝)
