高原肺水肿大鼠肺组织差异表达microRNA的筛选、生物学功能分析及验证
2020-03-19刘三立蔡未刘子泉侯世科樊毫军丁辉
刘三立,蔡未,刘子泉,侯世科,樊毫军,丁辉
1中国人民武装警察部队后勤学院,天津300162;2中国人民解放军98985部队卫生连;3天津大学灾难医学研究院
高原肺水肿是发生在健康人群快速进入海拔2 500 m以上高度时常出现的威胁生命的急性并发症[1, 2]。高原肺水肿起病急,进展快,如救治不及时,可在较短时间内发展至呼吸衰竭甚至死亡,病死率36.36%~90%[3],是严重威胁高原人群健康和生命的急性重症高原病。个体所处的海拔高度、上升速率、预适应能力及其易感性是导致高原肺水肿发生的主要因素[4]。张西南等[5]发现,385例急性高原病患者中重型高原病175例,死亡5例,病死率1.3%。我国高原面积大,常驻人口多,如何预防高原肺水肿的发生及减轻其严重程度是高原医学领域亟待解决的重大课题。
microRNA(miRNA)是一组21~25 nt长的非编码蛋白质的短序列RNA,能通过碱基配对与mRNA分子的3′UTR相结合,降解mRNA或抑制靶基因的翻译,从而对基因进行转录后表达的调控。2007年,Kulshreshtha 等[6]首次报道了缺氧与miRNA的关系,采用基因芯片技术筛选出了27个缺氧相关miRNA。Alam 等[7]首次研究了miRNA在高原肺水肿病理生理方面发挥的重要作用。大量缺氧介导的差异表达miRNA参与了与高原肺水肿发病机制密切相关的诸如离子通道、肺动脉高压、血管功能和应激反应等的调控过程,表明高原肺水肿受miRNA遗传调控作用的影响。2018年12月~2019年6月,本研究在低氧暴露不同时间条件下,建立稳定大鼠高原肺水肿模型,运用基因芯片技术筛选出差异表达高原肺水肿大鼠与健康大鼠肺组织差异表达miRNA,并进行靶基因预测及miRNA对靶基因的调控网络分析和生物学信息分析,揭示这些差异表达miRNA与高原肺水肿的内在联系,为探索高原肺水肿发病机制提供一种新的思路。
1 材料与方法
1.1 动物、试剂、仪器 SPF级成年雄性SD大鼠24只(购自军事医学科学院动物中心),体质量180~200 g;芯片类型:miRNA 4.0 Rat(美国Affymetrix公司):GeneChip WT Terminal Labeling and Controls Kit、Ambion WT Expression Kit、GeneChip Hybridization, Wash, and Stain Kit;芯片设备、软件(美国Affymetrix公司):基因芯片洗脱工作站GeneChip Fluidics Station 450、基因芯片杂交炉GeneChip Hybridization Oven 645、基因芯片扫描仪GeneChip Scanner 3000 7G、GeneChip GCOS软件Affymetrix GeneChip Operating Software;TRIzol RNA提取试剂(美国invitrogen公司),反转录试剂盒(日本TaKaRa公司),SYBR Green预混液(日本TaKaRa公司),IL-6、TNF-α、IL-10 ELISA检测试剂盒(武汉华美生物工程有限公司),BCA总蛋白试剂盒(北京索莱宝公司),荧光定量PCR仪(美国Bio-RAD公司),超微量紫外分光光度计(德国implen公司),DMI3000B生物倒置显微镜(德国Leica公司)。
1.2 高原肺水肿大鼠模型建立及鉴定 将大鼠随机分为4组:对照组和低氧24、48、72 h组,每组6只。低氧组使用数字化动物实验舱(武警后勤学院)模拟6 000 m低压低氧环境,运行时间依据每次造模时间设置,设置舱内温度为(10±2)℃,湿度保持在(60±10)%,舱内汞压 54.5 kPa,氧浓度 9.76%,氧分压9.9 kPa,泵切换时间720 min,高度上升/下降速率为10 m/s,启动制冷开启模式。对照组在SPF环境下饲养72 h。造模完成后,各组大鼠腹腔注射2%戊巴比妥(2 mL/0.1 kg)麻醉,取仰卧位固定。经会阴上方腹部正中做纵行切口,充分暴露腹腔后,腹主动脉抽取2 mL动脉血。采集的血样本4 ℃、3 000 r/min、离心后取血清保存于-80 ℃冰箱,采用ELISA法检测IL-6、TNF-α、IL-10。打开胸腔结扎右主支气管,气管插管通过左主支气管向左肺缓缓注入生理盐水4 mL,缓慢回抽生理盐水再重新注入,重复冲洗2~3次,将收集到的肺泡灌洗液(BALF)800 g,4 ℃离心10 min,保留上清液,采用ELISA法检测IL-6、TNF-α、IL-10。分离右肺,右上叶用于测量肺组织含水量,右中叶用4%多聚甲醛固定用于肺组织HE染色,右下叶-80 ℃低温冰箱中保存,进行后续miRNA芯片分析。低氧24、48、72 h组及对照组血清IL-6水平分别为(68.20±9.66)、(99.83±13.10)、(88.39±14.71)、(37.03±14.22)pg/μL;TNF-α水平分别为(218.19±25.76)、(311.08±34.91)、(281.68±32.92)、(173.95±24.04)pg/μL;IL-10水平分别为(39.34±4.51)、(54.05±7.57)、(64.70±8.22)、(23.98±5.91)pg/μL。低氧24、48、72 h组及对照组BALF中IL-6水平分别为(105.25±9.38)、(145.39±17.25)、(129.83±14.16)、(79.86±15.29)pg/μL;TNF-α水平分别为(303.94±27.79)、(370.14±34.00)、(365.74±19.58)、(260.61±29.92)pg/μL;IL-10水平分别为(53.88±5.40)、(66.47±6.48)、(79.21±9.82)、(38.37±5.34)pg/μL。低氧24、48、72 h组及对照组总蛋白量分别为(0.36±0.10)、(0.39±0.13)、(0.38±0.03)、(0.20±0.05)mg/μL。低氧24、48、72 h组及对照组肺含水量分别为0.79±0.01、0.80±0.02、0.80±0.01、0.78±0.02。与对照组相比,低氧24、48、72 h组血清和BALF中IL-6、IL-10、TNF-α水平升高,BALF中总蛋白含量增多(P均<0.01)。与对照组相比,低氧48、72 h组肺组织含水量增加(P均<0.05)。病理染色结果示,各低氧组表现出明显的肺泡及间质水肿、肺泡间隔增厚伴红细胞浸润、炎性渗出。以上结果提示高原肺水肿模型成功建立。
1.3 高原肺水肿大鼠肺组织差异表达miRNA筛选、靶基因预测及其相互作用分析 利用miRNA 4.0-Rat芯片(美国Affymetrix公司)对肺组织miRNA进行筛选。取1 μg 肺组织总RNA进行样本RNA质量检测。总RNA加尾后,加入FlashTag Biotin HSR Ligation Mix(美国Affymetrix公司)至已加尾混合液中按照厂家说明进行生物素标记和纯化,用GeneChip Hybridization Oven 645(美国Affymetrix公司)与Affymetrix miRNA 4.0 microarray(美国Affymetrix公司)杂交,用GeneChip Fluidics Station 450(美国Affymetrix公司)进行芯片洗脱,然后用GeneChip Scanner 3000 7G(美国Affymetrix公司)扫描芯片。原始数据由GeneChip GCOS(美国Affymetrix公司)分析处理,差异表达miRNA比较,P<0.05, FC>2.0被认为是差异基因。采用miRanda和TargetScan 两个数据库对得到的差异miRNA进行靶基因预测,最后对两种预测的结果取交集以增加结果的可靠性。
1.4 大鼠肺组织差异表达miRNA基因本体功能(GO)及信号通路分析 将以上得到的差异表达miRNA靶基因基于 GO数据库进行基因功能注释,KEGG数据库进行信号通路注释,显著性筛选的标准,P<0.05。
1.5 高原肺水肿大鼠肺组织差异表达miRNA检测 在连续上调和下调的差异表达miRNA中选择相对表达强度上调较为明显和相对表达强度下调较明显者进行qPCR。使用TRIzol试剂提取样本组织总RNA。RNA在260 nm处通过吸光度定量。使用反转录试剂盒进行逆转录反应。qPCR仪进行实时PCR反应。采用U6作为内参,以2-ΔΔCt表示目的基因表达水平。
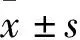
2 结果
2.1 高原肺水肿大鼠肺组织差异表达miRNA 以FC>2.0为准,筛选出的差异表达miRNA有14种,其中上调的有5种,分别为rno-miR-324-5p、rno-miR-6215、rno-miR-3473、rno-miR-344g、rno-miR-541-5p;下调的有9种,分别为rno-miR-3544、rno-miR-322-3p、rno-miR-203a-3p、rno-miR-342-5p、rno-miR-3556b、rno-miR-532-5p、rno-miR-101a-5p、rno-miR-361-3p、rno-miR-187-3p。在这些差异表达miRNA中,随着时间推移,表达量持续上调和下调的miRNA有5种。其中上调的有2种,分别为rno-miR-344g和rno-miR-541-5p,四组之间rno-miR-541-5p比较,低氧72 h组<低氧24 h组、低氧48 h组、对照组(P均<0.05),而低氧48 h、低氧24 h组与对照组之间比较,P均>0.05;四组之间rno-miR-344g比较,P>0.05。下调的有3种,分别为rno-miR-3556b、rno-miR-101a-5p、rno-miR-361-3p,四组rno-miR-3556b比较:低氧48、72 h组<对照组(P均<0.05);四组rno-miR-101a-5p、rno-miR-361-3p比较:低氧48、72 h组<低氧24 h组、对照组(P均<0.05)。
2.2 高原肺水肿大鼠肺组织差异表达miRNA靶基因预测结果及其对靶基因的调控网络 低氧组与常氧组差异表达miRNA靶基因在miRanda和TargetScan数据库中的交集较多。差异miRNA调控的靶基因很多,如:rno-miR-344g(76个)、rno-miR-541-5p(111个)、rno-miR-3556b(5个)、rno-miR-361-3p(198个)、rno-miR-101a-5p(48个)、rno-miR-187-3p(19个)、rno-miR-203a-3p(113个)、rno-miR-322-3p(36个)、rno-miR-324-5p(77个)、rno-miR-342-5p(32个)、rno-miR-3473(50个)、rno-miR-3544(39个)、rno-miR-532-5p(58个)、rno-miR-6215(15个)。同时,这些miRNA之间存在多个相互关联的靶基因。其中,rno-miR-203a-3p与rno-miR-541-5p的交互靶基因有1个(Nrp1),与rno-miR-324-5p的交互靶基因有1个(Med22),与rno-miR-322-3p的交互靶基因有1个(Plekhh2),与rno-miR-3544的交互靶基因有1个(Parg);rno-miR-101a-5p与rno-miR-541-5p的交互靶基因有1个(Rora),与rno-miR-361-3p的交互靶基因有2个(Ldoc1l、Pnoc),与rno-miR-187-3p的交互靶基因有1个(Naa38);rno-miR-541-5p与rno-miR-361-3p的交互靶基因有1个(Dcx),与rno-miR-342-5p的交互靶基因有2个(Lrch1、Slc1a2),与rno-miR-532-5p的交互靶基因有1个(C1orf21);rno-miR-361-3p与rno-miR-324-5p的交互靶基因有2个(Scn2b、Lama4),与rno-miR-322-3p的交互靶基因有2个(Otud3、Zc3h13),与rno-miR-3544的交互靶基因有2个(Tle3、Ube2d4),与rno-miR-3473的交互靶基因有1个(Zbtb24),与rno-miR-344g的交互靶基因有1个(Sbno1); rno-miR-324-5p与miR-3473的交互靶基因有1个(Hoxb3);rno-miR-3473与rno-miR-532-5p的交互靶基因有1个(Znf652);rno-miR-532-5p与rno-miR-344g的交互靶基因有1个(Bsnd);rno-miR-344g与rno-miR-6215的交互靶基因有1个(Pmfbp1)。
2.3 高原肺水肿大鼠肺组织差异表达miRNA的基因本体功能及信号通路 高原肺水肿大鼠肺组织差异表达miRNA参与的生物学过程有66种,主要涉及核酸的调控如RNA聚合酶Ⅱ启动子转录的负调控(GO:0000122)、RNA聚合酶Ⅱ启动子转录的调控(GO:0006357)等,蛋白泛素化如蛋白K63有关泛素化(GO:0070534)、蛋白K6有关泛素化(GO:0070534)等,信号通路如Wnt信号通路(GO:0016055)、胞内受体信号通路(GO:0030522)等,血管生理如血管发育(GO:0001568)、血管新生(GO:0001525)等以及生物进程如大脑发育(GO:0007420)、神经系统发育(GO:0007399)等。在细胞的组成成分层面有28种,主要涉及细胞内组成如核浆(GO:0005654)、胞质(GO:0005829)等,细胞器如蛋白复合体(GO:0043234)、多核糖体(GO:0005844)等,细胞膜如基膜(GO:0005605)、膜组成部分(GO:0005887),细胞功能如神经元投射(GO:0043005)、细胞黏附连接(GO:0005913)等。在基因分子功能层面有30种,主要涉及与生物分子有关的结合如蛋白结合(GO:0019901)、转录因子结合(GO:0033613)等,生物分子的活性如泛素蛋白转移活性(GO:0004842)、氯离子通道活性(GO:0005254),分子功能(GO:0003674)等。差异表达miRNA涉及的信号通路有11种,主要有PI3K-Akt信号通路(pathway ID:rno04151)、癌症通路(pathway ID:rno05200)、细胞外基质受体相互作用通路(pathway ID:rno04512)、内质网蛋白加工通路(pathway ID:rno04141)、泛素介导的蛋白水解作用通路(pathway ID:rno04120)等。
2.4 四组关键差异miRNA 比较 在5种连续上调和下调的高原肺水肿大鼠肺组织差异表达miRNA中选择相对表达强度上调较为明显的rno-miR-541-5p,和相对表达强度下调较为明显的rno-miR-101a-5p进行荧光定量qPCR验证。低氧72 h组、低氧48 h组、低氧24 h组、对照组rno-miR-541-5p分别为1.72±0.18、1.43±0.36、1.46±0.14、1,rno-miR-101a-5p分别为0.64±0.23、0.46±0.25、0.91±0.23、1。与对照组相比,rno-miR-541-5p在低氧暴露各组相对表达量显著升高(P均<0.05);与对照组相比,rno-miR-101a-5p在低氧暴露组相对表达量下降,其中低氧48 h组差异有统计学意义(P<0.05),低氧24 h组与低氧48 h组比较,P<0.05。与miRNA芯片结论一致。
3 讨论
当个体在适应高原环境的过程中,机体的基因表达会发生一系列变化,以平衡缺氧引起的应激,其调控元件如miRNA参与了高原肺水肿的发展过程。文献报道了多种miRNA参与调控了急性肺损伤的发生,并可能成为分子治疗的潜在靶标。Zhang等[8]研究发现,通过药物靶向miR-381介导的NLRP3的表达,能够减轻LPS诱导的急性肺损伤。Zhou等[9]研究发现,miR-126-3p的过表达能够抑制炎症蛋白HMGB1和血管通透性因子VEGF-A,增加紧密连接蛋白的表达。Zhang等[10]证实了miR-96和miR-330均可通过与AQP5的3′-非翻译区(UTR)结合来调节AQP5的表达,从而减轻LPS诱导的急性肺水肿。有研究[11, 12]发现,miR-144和miR-155被认为是两种相互拮抗的miRNA,miR-144可减弱肺血管中ROCK1/MLC通路的激活,起到抗炎作用,而在miR-155基因敲除的小鼠中,Ang-2- Tie-2-MLC信号通路活性降低,炎症相关蛋白被下调,表明miR-155对LPS诱导的急性肺损伤具有促进作用。相关研究表明,miR-16、miR-20b,miR-22、miR-206和miR-17/92在低氧暴露中表达下调,抑制离子通道并增加肺动脉压,从而导致血管功能障碍和细胞完整性丧失。而miR-23b,miR-26a和miR-155抑制TGF信号传导并导致肺动脉压升高,miR-210能够抑制线粒体功能,这些生理功能与高原肺水肿密切相关,表明高原肺水肿是一种器官特异性(肺)和环境应激诱导的疾病[7]。
本研究在建立低氧暴露24、48、72 h模型的基础上,运用基因芯片技术,以FC>2.0为标准,选取了14种差异表达miRNA,其中上调的有5种,下调的有9种。miRNA对基因的调控网络发现,上述miRNA存在多种交互基因如Scn2b、Lama4、Hoxb3、Zc3h13等,预测它们可能共同参与调控了高原肺水肿的发生发展,其具体机制有待进一步研究。进一步筛选分析发现,与对照组相比,在低氧24、48、72 h组表达量连续上调或下调的miRNA有5种,其中上调的miRNA有rno-miR-344g和rno-miR-541-5p,下调的miRNA有rno-miR-3556b、rno-miR-101a-5p、rno-miR-361-3p。说明上述5种miRNA在低氧暴露的同时,随着低氧时间的延长,相对表达呈现出明显的时间相关性,其在低氧中的上调或下调影响着特定靶基因表达的增加或降低,可能发挥着诸如改变细胞整合和屏障功能、血管渗透性等高原肺水肿的病理生理因素等[7]。有文献报道miR-541-5p与肺纤维化的发生有关,其通过在蛋白翻译水平上调节PDE1A的表达而成为肺成纤维细胞的关键因子,同时,我们通过基因调控网络分析发现RORα是rno-miR-101a-5p参与血管新生的靶基因之一,这两种在本研究中上调和下调较为明显的miRNA 是否也通过特定机制发挥对高原肺水肿的调控作用,有待进一步研究[13~15]。
低氧是引起高原肺水肿的重要因素,高原引发肺水肿的发生机制主要有:①低氧引起肺动脉强烈收缩、肺血管损伤、通透性增加导致液体渗入肺泡增多,肺水肿形成;②低氧诱导免疫因子和炎症因子增多,促进肺动脉血管通透性增加;③低氧抑制Na+-K+-ATP酶活性,引起肺泡上皮细胞钠离子和水等清除障碍,导致肺组织水潴留[16, 17]。有文献报道,在人肺微血管内皮细胞中,通过激活PI3K/Akt信号通路可以改善内皮细胞屏障功能障碍。邓旺等[18, 19]研究发现,激活PI3K /Akt 信号通路能上调ENaC的表达,清除急性肺损伤时肺内多余的水肿液,从而减轻肺组织损伤,表明PI3K/Akt信号通路在调节肺水肿方面发挥着重要作用。Villar等[20]在大鼠COPD模型中发现抑制Wnt信号通路可减轻肺组织损伤,改善肺功能并重新建立肺泡上皮细胞表面标志物的表达,说明Wnt信号通路影响着肺泡-毛细血管屏障的通透性。在本研究中,我们通过信号通路的显著性分析发现miRNA对Wnt、PI3K/Akt信号通路也具有相关调控作用,说明筛选出的相关miRNA可能通过调控特定信号通路对高原肺水肿的产生起到正性或负性调节作用。此外,通过对14种miRNA的靶基因进行关于细胞组分、分子功能、生物过程的富集分析,我们发现miR-101a-5p、miR-203a-3p、rno-miR-361-3p、rno-miR-541-5p、rno-miR-324-5p、rno-miR-344g、rno-miR-532-5p、rno-miR-3473等具有调控血管新生的功能,而血管新生可以诱导肺毛细血管通透性的增加导致肺水肿的发生[21, 22]。结合本研究对miRNA的生物信息学分析,我们推断高原低氧下差异表达的miRNA参与了对高原肺水肿的调控过程。
综上所述,在高原肺水肿大鼠肺组织中差异表达miRNA有14种,其中持续上调的有rno-miR-344g、rno-miR-541-5p,下调的有rno-miR-3556b、rno-miR-101a-5p、rno-miR-361-3p。这些miRNA之间存在许多交互靶基因,并与其靶基因形成复杂的调控网络。主要参与核酸的调控等生物学过程,影响细胞内组成如核浆、胞质等,涉及生物分子有关的结合,相关的信号通路主要有PI3K-Akt信号通路等。高原肺水肿大鼠肺组织中差异表达miRNA及其靶基因参与了诸如血管新生、血管发育、细胞黏附连接、转录因子结合、PI3K-Akt信号通路等与高原肺水肿形成机制紧密相关的细胞分子生物学过程,表明这些差异表达miRNA可能参与调控了高原肺水肿发生的可能机制,这为进一步探索高原肺水肿发病机制及其预防治疗提供了新思路。
