生物钟基因Bmal1通过氧化应激信号通路调节胰岛β细胞凋亡的研究
2020-01-10叶绿许伟红薛静张婷朱亮杜燕吴华香
叶绿 许伟红 薛静 张婷 朱亮 杜燕 吴华香
生物钟存在于机体内几乎所有的细胞中,调节机体的行为和生理过程,包括能量代谢、内分泌激素的昼夜节律[1]。生物钟包括正向调节作用的circadian locomotor output cycles kaput(Clock)、brain and muscle Arnt-like protein-1(Bmal1)和负向调节作用的 Period1-3(Per1-3)、Cryptochrome1-2(Cry1-2)。这些生物钟基因相互作用,在转录和翻译水平构成自主的反馈环路[2]。
生物钟功能紊乱与代谢综合征、糖尿病、心血管疾病密切相关。研究表明,睡眠障碍、轮班工作导致糖尿病发病率增加[3]。人群生物钟基因的多态性和突变与心血管疾病、糖尿病和肥胖相关[4-6]。Per2基因缺失导致小鼠肥胖[7],Clock和Bmal1基因缺失引起小鼠血糖升高[8-9]。但目前多数动物模型为全身性基因敲除,除外环境和摄食行为等影响后,生物钟基因异常导致β细胞功能障碍的机制不明确。有研究发现,特异性敲除β细胞Bmal1基因比全身性敲除Bmal1基因的小鼠,胰岛素分泌下降、糖耐量受损和血糖升高更明显。特异性敲除小鼠β细胞Clock和Per2基因未导致胰岛功能障碍[10]。这些研究证实Bmal1对维持胰岛β细胞正常功能的重要作用。
β细胞功能障碍是糖尿病发病的中心环节,细胞凋亡在胰岛功能障碍起重要作用[11-13]。目前生物钟基因对细胞凋亡的调控机制尚不明确。本研究应用RNA干扰,沉默胰岛β细胞Bmal1基因表达,观察Bmal1基因缺失对β细胞的影响及其机制,现报道如下。
1 材料和方法
1.1 细胞、主要仪器和试剂 大鼠胰岛β细胞瘤株(insulinoma cell line,INS-1,武汉普诺赛生命科技有限公司,货号:CL-0368)。全自动酶标仪(美国Thermo scientific公司,型号:Multiskan MK3),实时荧光定量PCR仪(美国 Applied Biosystems公司,型号:QuantStudio6),流式细胞仪(美国Beckman公司,型号:CytoFLEX),荧光显微镜(日本Olympus公司,型号:BX53),增强型ATP检测试剂盒(上海碧云天公司,批号:S0027),ROS检测试剂盒(上海碧云天公司,批号:S0033),RNA抽提试剂盒(美国Ambion公司,批号:15596-026),实时荧光定量PCR试剂盒(美国Vazyme公司,批号:Q111-02),TUNEL检测试剂盒(美国Roche Applied Science公司,批号:12156792910)。
1.2 实验方法
1.2.1 细胞培养 胰岛β细胞培养于含5%CO2的37℃培养箱中,培养条件为含10%胎牛血清(美国Gibco公司,批号:10091-148)、2mmol/L左旋谷氨酰胺(美国Gibco公司,批号:25030-081)、青霉素-链霉素溶液(美国 Gibco 公司,批号:15070-063)、10 mmol/L HEPES(美国 Gibco公司,批号:15630-080)、50μmol/L 巯基乙醇(美国 Gibco公司,批号:21985023)、11.2mmol/L 葡萄糖、1mmol/L丙酮酸钠的RPMI-1640培养液(美国Gibco公司,批号:11875-093)。
1.2.2 实验分组 正常对照组(含5.5mmol/L葡萄糖,NC组)、高糖组(含33.3mmol/L葡萄糖)、正常糖浓度+无关RNA转染组(含5.5mmol/L葡萄糖,NC+siRNA组)、正常糖浓度+Bmal1基因沉默组(NC+Bmal1-/-组)、高糖+无关RNA转染组(含33.3mmol/L葡萄糖,高糖+siRNA组)、高糖+Bmal1基因沉默组(高糖+Bmal1-/-组)。
1.2.3 细胞转染及沉默Bmal1基因 取对数生长期,生长状态良好的INS-1β细胞,每孔2×105个接种于细胞培养6孔板,在37℃、5%CO2培养箱中培养过夜。细胞密度达到70%时进行转染,在转染前2h换成无血清1640培养基。取5μl LipofectamineTM2000(美国Invitrogen 公司,批号:11668-027)加至 100μl opti-MEM 中,室温下静置5min;混合LipofectamineTM2000和核酸的稀释液,室温下静置20min。每个培养孔加混合液200μl。细胞于37℃、CO2培养箱中培养,6h后换为正常培养基,37℃、5%CO2培养箱中培养过夜。
1.2.4 三磷酸腺苷(adenosine triphosphate,ATP)浓度测定 去除培养液,每孔加入200μl裂解液裂解细胞。裂解后4℃12 000r/min离心5min,收集上清液。加100μlATP工作液至检测孔内,室温静置5min。在检测孔内加入20μl样品或标准品,迅速混匀,2s后用化学发光仪测定。
1.2.5 流式细胞仪检测细胞活性氧 (reactive oxygen species,ROS)水平 使用0.25%胰酶消化细胞,终止消化后,1 500r/min离心5min收集细胞,去上清液。用PBS洗涤细胞2次,1 500r/min离心5min。根据DCFH-DA细胞ROS检测试剂盒的说明进行操作:加入1ml DCFH;37℃培养箱孵育20min,每隔3min混匀1次;用无血清培养基洗涤细胞3次。随后用流式细胞仪进行检测。
1.2.6 TUNEL法检测细胞凋亡 将爬有细胞的玻片用4%多聚甲醛溶液室温固定25min,用PBS洗涤3次,每次5min。随后,细胞爬片浸入0.1%TritonX-100溶液中10min进行冰上操作,PBS浸洗2次,每次5min。然后加入50μl TUNEL反应混合液,37℃避光孵育60min。随后用PBS洗涤3次,每次5min。加入DAPI避光孵育5min,PBST洗涤4次,每次5min。使用抗荧光淬灭封片液封片,荧光显微镜观察及采集图像。
1.2.7 实时荧光定量PCR(qRT-PCR)检测 根据RNA抽提试剂盒说明书提取细胞RNA,应用微量分光光度计测定波长为260、280nm的光密度值,计算RNA的纯度和浓度。将RNA反转录为cDNA。目的基因Bmal1、去乙酰化酶sirtuin 1(Sirt1)以及管家基因GAPDH的引物序列由杭州擎科生物公司合成。PCR反应条件为95°C预变性10min;95℃ 变性 30s,60°C 退火延伸 30s,共 40个循环。2-ΔΔCt法计算Bmal1、Sirt1基因、管家基因 GAPDH mRNA表达。
1.3 统计学处理 采用 SPSS16.0统计软件。测得计量资料采用±se表 示,多组间比较用单因素方差分析,两两比较采用LSD-t检验。P<0.05为差异有统计学意义。
2 结果
2.1 Bmal1沉默后对ATP生成的影响 见图1。
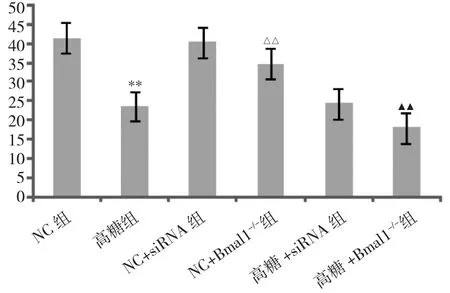
图1 Bmal1沉默后对ATP生成的影响(与NC组比较,**P<0.01;与 NC+siRNA 组比较,△△P<0.01;与高糖 +siRNA组比较,▲▲P<0.01)
由图1可见,与NC+siRNA组比较,NC+Bmal1-/-组ATP 含量下降(P<0.01);与 NC 组比较,高糖组 ATP 含量明显下降(P<0.01);与高糖+siRNA 组比较,高糖+Bmal1-/-组 ATP 明显下降(P<0.01)。
2.2 Bmal1沉默后对ROS生成的影响 见图2。

图2 Bmal1沉默后对ROS生成的影响(与NC组比较,**P<0.01;与NC+siRNA组比较,△△P<0.01;与高糖+siRNA组比较,▲▲P<0.01)
由图2可见,与NC+siRNA组比较,NC+Bmal1-/-组ROS 水平升高,(P<0.01);与 NC 组比较,高糖组 ROS生成增加(P<0.01);与高糖+siRNA组比较,高糖+Bmal1-/-组 ROS 增加更明显(P<0.01)。
2.3 Bmal1基因对Sirt1mRNA表达的影响 见图3。
由图3可见,与NC+siRNA组比较,NC+Bmal1-/-组Sirt1 mRNA 表达下降(P<0.01);与 NC 组比较,高糖组Sirt1 mRNA表达下降(P<0.01);与高糖+siRNA 组相比,高糖+Bmal1-/-组Sirt1 mRNA表达有下降趋势,下降幅度为 20.7%,但差异无统计学意义(P >0.05)。
2.4 Bmal1基因对细胞凋亡的影响 NC组、高糖组、NC+siRNA组、NC+Bmal1-/-组、高糖+siRNA组、高糖+Bmal1-/-组 的 凋 亡 率 分 别 为 4.31%、10.26%、4.86% 、6.72%、13.21%、25.36%。与 NC+siRNA 组比较,NC+Bmal1-/-组凋亡率无明显变化(P >0.05);与 NC 组比较,高糖组细胞凋亡增加(P<0.05);与高糖+siRNA组比较,高糖+Bmal1-/-组细胞凋亡率明显增加(P<0.01),见图 4(插页)。
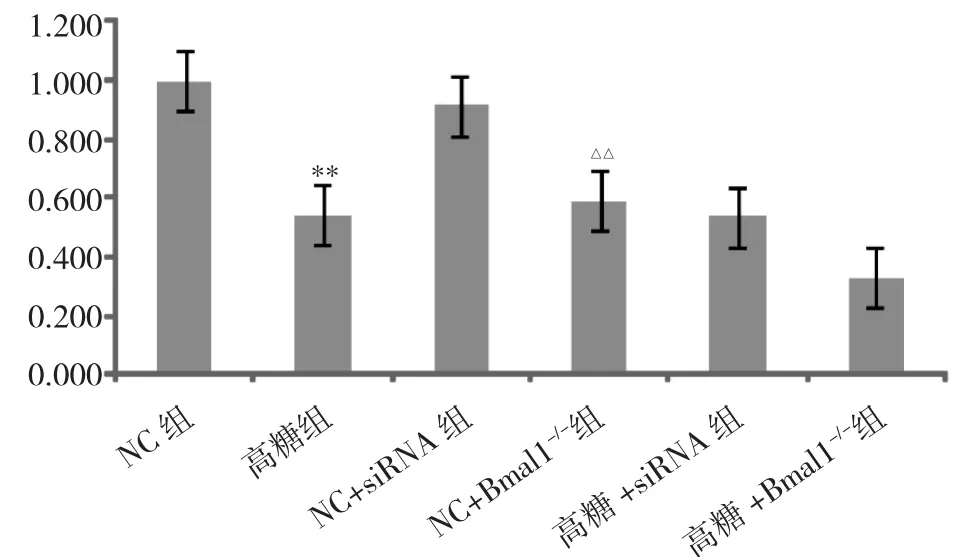
图3 Bmal1基因对Sirt1mRNA表达的影响(与NC组比较,**P<0.01;与 NC+siRNA 组比较,△△P<0.01)
3 讨论
糖尿病严重危害人类健康,β细胞功能障碍在糖尿病发病中起关键作用,其主要表现为胰岛素分泌不足和分泌节律异常。研究表明,胰岛素分泌节律主要由生物钟调控[14-16],因此,生物钟可能通过调控胰岛β细胞功能影响糖尿病的发生、发展。有关胰岛β细胞生物钟基因的研究数据较少。有学者研究发现,特异性敲除胰岛β细胞Bmal1基因的小鼠比Bmal1-/-小鼠的胰岛素分泌下降更明显,尽管Bmal1-/-β细胞大小和增殖不受影响,其葡萄糖刺激的胰岛素分泌下降近60%[10]。这些资料提示生物钟基因Bmal1参与糖尿病发病,对维持β细胞正常功能有重要作用。
细胞凋亡在胰岛β功能障碍起重要作用[11-13]。受凋亡信号刺激时,内源性ROS过量生成、细胞色素C及凋亡诱导因子等释放入细胞质,激活半胱氨酸蛋白酶9(Caspase9),导致细胞DNA修复功能损伤、核酸内切酶活化,最终导致细胞凋亡[17]。此外,在糖尿病人群以及动物模型均发现氧化应激增强[18-19]。但生物钟基因在胰岛β细胞凋亡过程中的机制尚不明确。本研究对不同干预因素处理后的β细胞凋亡情况进行了检测,结果发现,高糖环境下,β细胞凋亡增加,这种作用在Bmal1基因沉默后更明显。此外,结果显示,Bmal1基因沉默后,ATP含量下降,ROS水平升高;在高糖环境下,Bmal1基因沉默组ATP水平明显下降,ROS显著增加。ROS氧化活性极强,其增加导致细胞结构损伤和代谢障碍。ATP是生物体各项生命活动最直接的能量来源。故ROS增加,ATP减少可导致相应器官组织功能障碍。有文献报道,在Bmal1-/-小鼠的脏器如肾脏、心脏和脾脏等,均发现ROS明显增加,导致相应组织细胞氧化应激反应增强,引起细胞功能障碍[20]。由于胰岛β细胞内抗氧化酶基因表达水平较低,如超氧化物歧化酶含量仅为肝脏的30%,过氧化氢酶及谷胱甘肽过氧化物酶仅为肝脏的5%[21],因此,β细胞清除ROS和抗氧化能力较弱,对氧化应激损伤更敏感。Lee等[22-23]的研究显示,Bmal1基因缺失使氧化应激增强,导致β细胞功能障碍,可能的机制是Bmal1基因缺失,导致线粒体解偶联蛋白(uncoupling protein2,UCP2)上调和抗氧化反应基因表达改变(如调控抗氧化反应的关键因子核因子E2相关因子2及其调控的靶基因如Sestrin2,过氧化还原酶3,谷氨酸半胱氨酸连接酶催化亚基和谷氨酸半胱氨酸连接酶修饰亚基下降),最终导致β细胞功能衰竭。这些资料提示Bmal1基因参与维持胰岛β细胞的正常功能。
本研究发现,Bmal1基因沉默后,Sirt1基因表达下降;在高糖环境下,Sirt1基因表达下降;Bmal1基因沉默组更明显,下降幅度为20.7%。去乙酰化酶Sirt1在细胞凋亡中起重要的调节作用,其主要通过组蛋白/非组蛋白去乙酰化作用影响基因转录,发挥抗氧化应激作用,减少细胞凋亡,增加细胞存活率。UCP2可使线粒体氧化磷酸化解偶联,ATP合成减少,影响ATP敏感的K+通道开放,导致细胞内Ca2+下降,抑制胞吐作用,减少胰岛素的释放;Sirt1可通过抑制UCP2间接促进胰岛素分泌[24]。
本研究结果初步提示生物钟基因Bmal1可通过调节氧化应激信号通路影响胰岛β细胞功能和凋亡。国内外对胰岛β细胞功能障碍进行了长期深入研究,但目前尚无有效保护和恢复胰岛β细胞功能的治疗措施。因此,进一步研究胰岛β细胞功能障碍的发病机制及其信号传导通路,对寻找有效的治疗靶点具有重要意义和潜在的临床价值。
