海岛棉棉铃阶段性发育与产量品质的关系
2020-01-02曹新川胡守林韩秀锋何良荣郭伟锋
曹新川 胡守林 韩秀锋 何良荣 郭伟锋
研究简报
海岛棉棉铃阶段性发育与产量品质的关系
曹新川 胡守林 韩秀锋 何良荣 郭伟锋*
塔里木大学植物科学学院, 新疆阿拉尔 843300
运用包含环境互作效应的ADM发育遗传模型对3年海岛棉亲本及F1代不同发育时期单铃重及发育特点与产量、品质的关系研究表明, 海岛棉各时期单铃重对最终单铃重、纤维品质性状选择效果较好; 前期(10、20 d)单铃重大, 会减少无效铃数, 降低衣分; 后期(60、70 d)单铃重大, 会增加无效铃数, 提高衣分。而中期(30、40、50 d)单铃重对最终单株产量有一定选择效果。棉铃体积膨大期(0~10 d、10~20 d)净增量较大, 单铃重增加, 其他产量性状降低; 纤维长度、反射率、黄度等品质性状改善; 棉铃内容物充实期(30~40、40~50、60~70 d)干重增量大, 主要纤维品质性状变差。选择单铃重进入快增期较晚、生长速率较大而出现时间较晚的材料将提高单株产量。纤维形成过程中, 同化物质较早进入棉铃, 经历缓慢增长, 使纤维逐渐成熟而形成优良的品质。棉铃发育受到明显的母体效应影响, 因而选择合适的母本对改良纤维品质性状较为重要。
海岛棉; 单铃重; 发育遗传; 发育特点; 遗传相关
南疆塔里木盆地是我国海岛棉的最大适宜棉区。棉铃作为重要的产量、品质形成器官, 一直是研究的热点。曾有对棉铃各部分干物质发育、纤维建成及发育规律的许多研究[1-6], 关于棉花各性状遗传[7-10], 以及发育遗传模型在水稻、油菜、大麦、棉花等作物遗传分析中的应用的相关报道较多[11-16], 但运用该模型对海岛棉各时期单铃重和生长发育特点与最终产量品质的关系研究还未见报道。本文首先用Logistic模型拟合, 计算各材料的生长特征值, 运用包含环境互作效应的ADM发育遗传模型研究3年海岛棉不同发育时期单铃重及生长特征值与产量、品质的关系, 为杂种后代的选择、杂种优势的利用和遗传改良效率的提高和海岛棉的丰产栽培提供了理论依据。
1 材料与方法
1.1 试验材料
母本为4个零式海岛棉品种新海14、新海17、新海18和新海21, 父本为4个外引长果枝海岛棉品种吉扎45、吉扎75、吉扎81和A杂交铃, 于2002年、2009年、2012年按NCII交配设计配制16个杂交组合。
1.2 试验方法
在塔里木大学农试站将供试亲本及F1代分别于2003年、2010年、2013年4月20日左右人工点播种植。采用随机区组设计, 3次重复, 4个行区, 行长5 m, 株行配置(30+55+30) cm ×10.5 cm。其他管理同大田。7月初观察开花情况。于7月9日选取中部节位当日开的花挂牌, 作为铃发育的起点, 在棉铃发育第10、20、30、40、50、60、70天从每个小区随机选取10个棉铃, 在室内分解为铃壳、种子、纤维, 将鲜样在105℃杀青30 min, 60℃烘至恒重, 称量各组分干重, 各时期种子与纤维干重均值相加即为该时期单铃重。10月中旬取各小区中部品质样, 然后收获生长均匀的5株, 于室内考种, 调查单株铃数、单铃重、衣分、单株皮棉等产量性状。用HFT9000测定纤维长度、整齐度、马克隆值、纤维强度、伸长率等棉花纤维品质性状。
1.3 统计分析方法
对3年8个亲本及16个F1代的7个时期单铃重数据利用=m/[1+e(a+bt)]方程进行曲线拟合, 根据模型性质分别估算速度特征值(最大累计速率m及其出现时间0)、时间特征值(进入快增期的时间t1、结束快增期的时间2和快增期的历期D)、生长特征值(最大累计量m、快增期干物质累计量GT)[17-18], 以确定各材料的生长发育特点。采用朱军教授提出的基因与环境互作的加性-显性-母体(ADM)发育遗传模型[19], 运用MINQUE(1)法估算不同发育时期或时段的单铃重及生长发育特点与产量、品质性状的各项遗传相关(加性相关A、显性相关D、母体相关M、加性×环境互作相关AE、显性×环境互作相关DE、母体×环境互作相关ME、环境相关E、遗传相关G和表现型相关P); 采用Jackknife抽样计算相关系数的估计值及标准误并进行T测验[20]。数据采用朱军教授提供的QGA Station统计软件运算, 所得结果整理并作图(相关系数不显著的均当作零, 保留显著或极显著的相关系数分量)。
2 结果与分析
2.1 海岛棉不同发育时期单铃重与产量品质性状的相关分析
2.1.1 海岛棉不同发育时期单铃重与产量性状的相关分析 从图1可以看出, 第10、20、30、60、70天的单铃重与单株铃数呈显著的遗传和表型负相关, 其中第30天主要由加性相关造成, 第60、70天主要由母体相关造成, 能够稳定遗传, 海岛棉的铃数与单铃重同步改良时较为困难。第20天的单铃重与单株无效铃呈显著遗传和表型负相关, 而第70天则相反, 即选择第20天时单铃重大, 减少无效铃数, 后期单铃重大, 增加无效铃数。第10、20、30、50天单铃重与衣分呈显著的负向遗传和表型相关, 即这些时期单铃重大, 衣分降低; 而第60、70天时为正向的遗传相关; 原因可能在于后期虽然有向纤维中输送的遗传倾向, 但早期同化物质主要往种子中输送。各时期单铃重除第40天外, 均与最终单铃重呈显著或极显著的正向遗传和表型相关, 可通过各时期的铃重对最终单铃重进行选择。单株产量与第30、40天的遗传相关和表型相关均为显著负相关, 通过这2个时期单铃重对单株产量有一定的负向选择效果。

图1 海岛棉不同发育时期单铃重与产量性状的相关性
G: 遗传相关;P: 表现型相关;E: 环境相关;ME: 母体与环境互作相关;DE: 显性与环境互作相关;AE: 加性与环境互作相关;M: 母体相关;D: 显性相关;A: 加性相关。
G: genetic correlation;P: phenotypical correlation;E: environmental correlation;ME: maternal and environment interaction correlation;DE: dominant and environment interaction correlation;AE: additive and environment interaction correlation;M: maternal correlation;D: dominant correlation;A: additive correlation.
2.1.2 海岛棉不同发育时期单铃重与品质性状的关系研究 从图2可以看出, 第10、20、30、50、60、70天的单铃重与纤维长度存在正向的显著或极显著遗传相关和表型相关, 通过这些时期的单铃重可对纤维长度进行选择, 由于造成遗传相关的主要因素为母体相关, 选择是可靠有效的。整齐度与第60、70天的单铃重呈正向极显著的遗传和表型相关, 主要由母体相关引起, 这2个时期选择单铃重大的材料可以提高整齐度。除第40天外, 其余时期的单铃重均与反射率呈正向的显著遗传相关和表型相关, 因此在前期和后期均可通过单铃重对反射率进行选择。第20、30、50、60、70天的单铃重与纤维强力间存在显著的正向遗传和表型相关, 第30天时主要由于加性相关造成, 在第50、60、70天主要由母体相关引起, 通过这4个时期单铃重对强度均可进行可靠的选择。伸长率与第50、60、70天的单铃重存在显著的遗传正相关, 是由于母体相关造成并最终在表型表现出来, 能进行有效选择。第10、20、30、50、60、70天的单铃重与黄度存在显著的遗传和表型负相关, 选择这些时期单铃重大的材料可以有效降低黄度。综合来看, 单铃重与纤维品质性状关系密切, 通过第50天、60天的单铃重选择除使马克隆值有所增加外可改良大多数纤维品质性状。
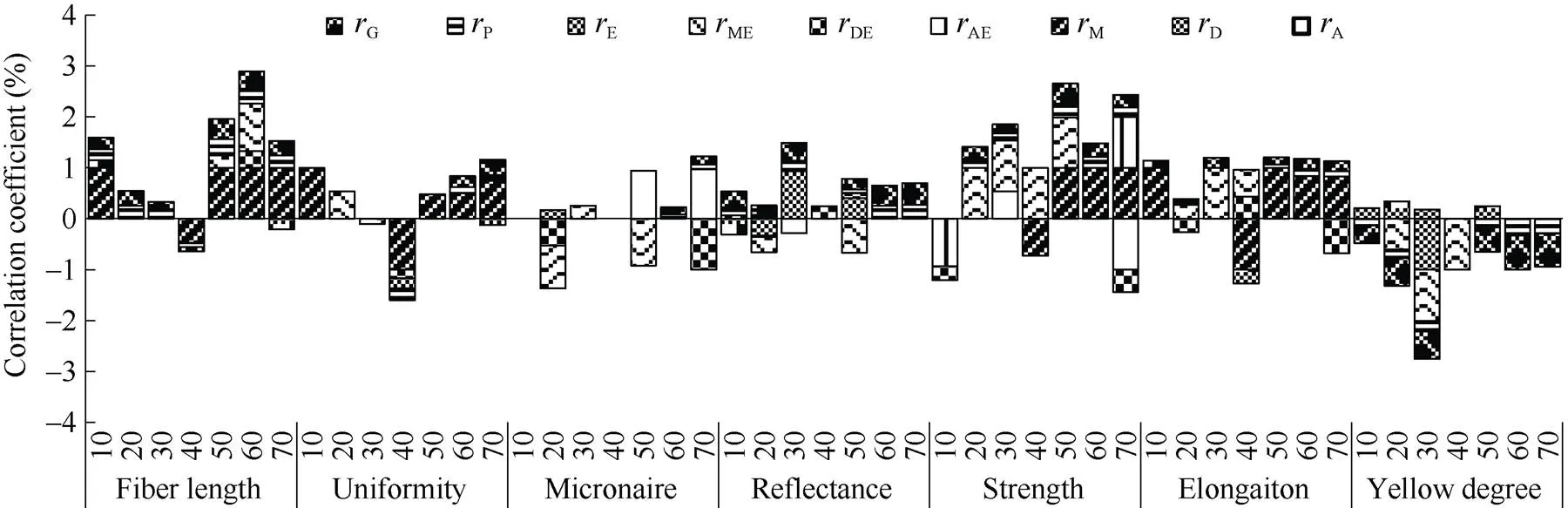
图2 海岛棉不同发育时期单铃重与品质性状的相关性
缩写同图1。Abbreviations are the same as those given in Fig. 1.
2.2 海岛棉单铃重增量与产量品质性状的相关分析
2.2.1 海岛棉单铃重增量与产量性状的相关分析 从图3可以看出, 第0~10、10~20、20~30、30~40、50~60、60~70天这6个时段的单铃重净增量与单株铃数的遗传与表型呈显著或极显著负相关, 其中10~20天还存在负向的加性相关和显性、母体与环境互作相关以及正向的环境相关; 第20~30、30~40、40~50天存在显著或极显著的母体与环境互作相关。第10~20天、40~50天单铃重与单株无效铃存在显著或极显著的加性相关、环境相关、表型相关和遗传相关。第10~20天、40~50天的单铃重与衣分存在显著或极显著的加性、显性与环境互作、母体与环境互作相关, 其中第10~20还存在显著或极显著的负向遗传相关和表型相关。第0~10、10~20、50~60、60~70天单铃重与最终单铃重间存在显著或极显著正向的遗传和表型相关, 其中0~10、10~20、40~50、50~60天存在正向加性或母体相关, 具较好选择可靠性。第10~40天3个时段的单铃重与单株籽棉、单株皮棉重均存在显著或极显著的负向遗传和表型相关, 其中第30~40天主要由母体相关造成。遗传与环境互作相关大多数时段起到主要作用。前期(棉铃体积膨大期)各时段净增量较大的, 其铃数较少, 衣分较低, 单铃重会增加, 单株产量较低。而后期这种关系逐渐被打破, 第50~70天铃重增重快有利于产量的增加。
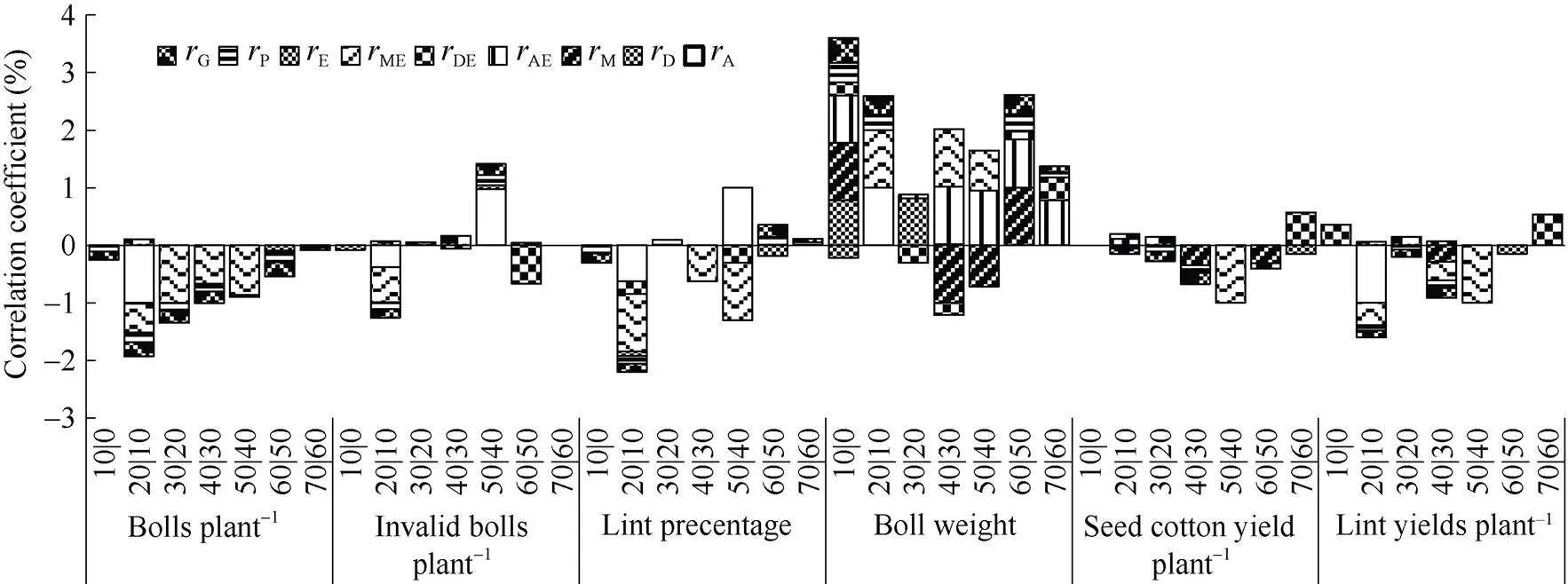
图3 海岛棉不同发育时段条件单铃重与产量性状的相关性
缩写同图1。Abbreviations are the same as those given in Fig. 1.
2.2.2 海岛棉单铃重增量与品质性状的相关分析 图4反映单铃重各时段增量对纤维品质的影响, 第0~20天这2个时段的单铃重与纤维长度存在显著的遗传和表型相关, 主要由母体或母体与环境相关造成, 这2个时段增重快, 纤维长度增加, 而第30~50、60~70天这3个时段存在显著或极显著的负母体相关、遗传相关和表型相关, 中后期(内容物充实期)铃重增速快, 纤维长度反而会降低, 这种效应主要通过母体遗传给后代。第40~50天单铃重与纤维强力存在显著或极显著的加性、母体、加性与环境、母体与环境以及负向的遗传和表型相关, 这一时段铃重增速慢, 将有利于纤维强度的增加, 其余时段关系不明显。体积膨大期(0~20 d)各时段棉铃干重增量大, 会使纤维长度增加, 反射率提高, 黄度降低。内容物充实期棉铃(30~50 d)各时段干重增量大, 会使纤维长度降低, 整齐度下降, 强力下降, 伸长率下降, 黄度上升。
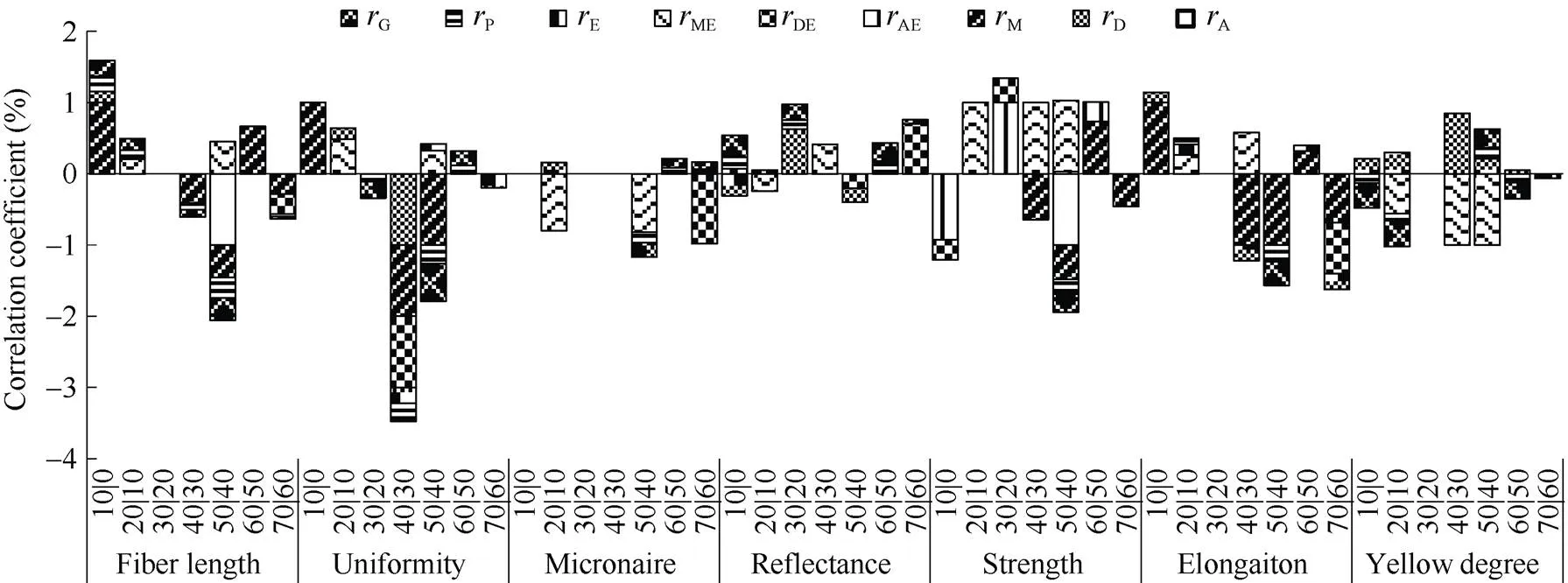
图4 海岛棉不同发育时段条件单铃重与品质性状的相关性
缩写同图1。Abbreviations are the same as those given in Fig. 1.
2.3 海岛棉单铃重发育特征值与产量品质性状的相关分析
2.3.1 海岛棉单铃重发育特征值与产量性状的相关分析
从图5可看出,m与单株铃数呈负向而与单铃重呈正向遗传相关和表型相关, 由能够稳定遗传的母体相关或加性相关引起, 单铃重累积量较大, 单株有效铃减少, 单铃重增加。0与衣分、单铃重、单株产量呈正向显著遗传和表型相关, 与衣分具正向母体相关, 与单铃重呈显著正向加性、显性、母体及环境相关, 互作相关不显著, 它们的关系较稳定;0大即进入快增期较晚, 衣分和单铃重增加, 单株产量提高。m与单株无效铃、单株产量呈显著正向遗传和表型相关, 最大生长速率较大, 无效铃增多, 单株产量增加。1与单株无效铃、衣分、单铃重、单株籽棉产量呈显著正向加性、遗传、表型相关, 与单株皮棉呈正向显著环境、遗传和表型相关, 快增期进入较晚无效铃数增多, 衣分和最终单株产量提高;2与单株铃数呈负向而与单铃重呈正向遗传和表型相关, 结束快增期的时期较晚, 使铃数减少, 单铃重提高。Δ与单铃铃数、单株无效铃呈现负向而与单铃重呈正向遗传、表型相关, 主要由母体与环境互作相关引起, 与单铃重还存在显性与环境互作相关, 快增期的历期长会使铃数减少, 无效铃减少, 单铃重增加, 与单株产量关系不大。因此要提高单株产量应选择进入快增期较晚、最大生长速率较大而出现时间相应较晚的材料。可能棉铃前期缓慢的增长使棉株体内物质分配均衡, 从而能够稳健生长, 铃数有增加的趋势; 进入快增期后同化物质快速往棉铃输送从而形成较高的单株产量。
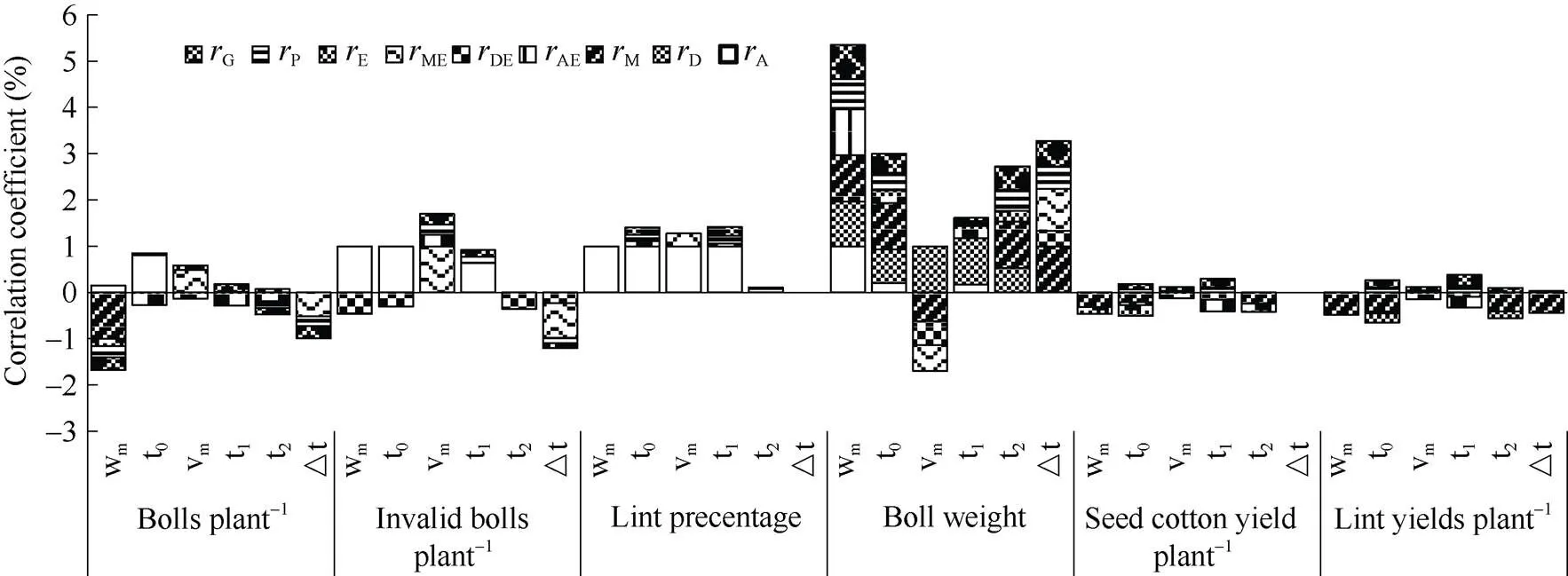
图5 海岛棉单铃重生长特征值与产量性状的相关性
缩写同图1。Abbreviations are the same as those given in Fig. 1.
2.3.2 海岛棉单铃重发育特征值与品质性状的相关分析
从图6可看出,m与长度、整齐度、马克隆值、反射率、强度、伸长率呈显著或极显著正向遗传相关和表型相关, 与黄度呈现显著或极显著负向遗传和表型相关, 其中纤维长度、整齐度、强度、伸长率呈显著或极显著正向母体相关, 与马克隆值呈显著的正向加性相关; 棉铃累积量较大, 其纤维长度较长, 整齐度较高, 马克隆值较大, 反射率较大, 强力较大, 伸长率较大, 黄度较低。0对纤维长度、整齐度、马克隆值、反射率、黄度等具有正向的表型和遗传相关, 其中与长度、整齐度、伸长率呈正向母体相关, 与马克隆值呈正向加性相关; 最大生长速度出现时间较晚会使纤维长度、整齐度、马克隆值、反射率提高, 黄度有所降低, 对强力和反射率影响不大。m与品质性状无稳定遗传的表型和遗传相关。1与马克隆值呈正向显著加性、遗传、表型相关, 进入快增期早可有效降低马克隆值。2、Δ与纤维长度、整齐度、伸长率呈显著正向母体、遗传和表型相关, 与马克隆值、反射率、黄度呈显著遗传和表型相关, 虽与纤维强度呈显著正向母体相关, 在当代能够表现出来, 但并没有在遗传和表型相关中表现出来, 因而不能稳定遗传给后代。因此纤维品质主要与单铃重的最终累积量和累积的历期有关, 而与速度无关。这应与纤维的形成机制有关。纤维形成过程中, 同化物质较早进入棉铃, 经历较长时期的缓慢增长, 使纤维逐渐成熟具有优良品质的纤维。
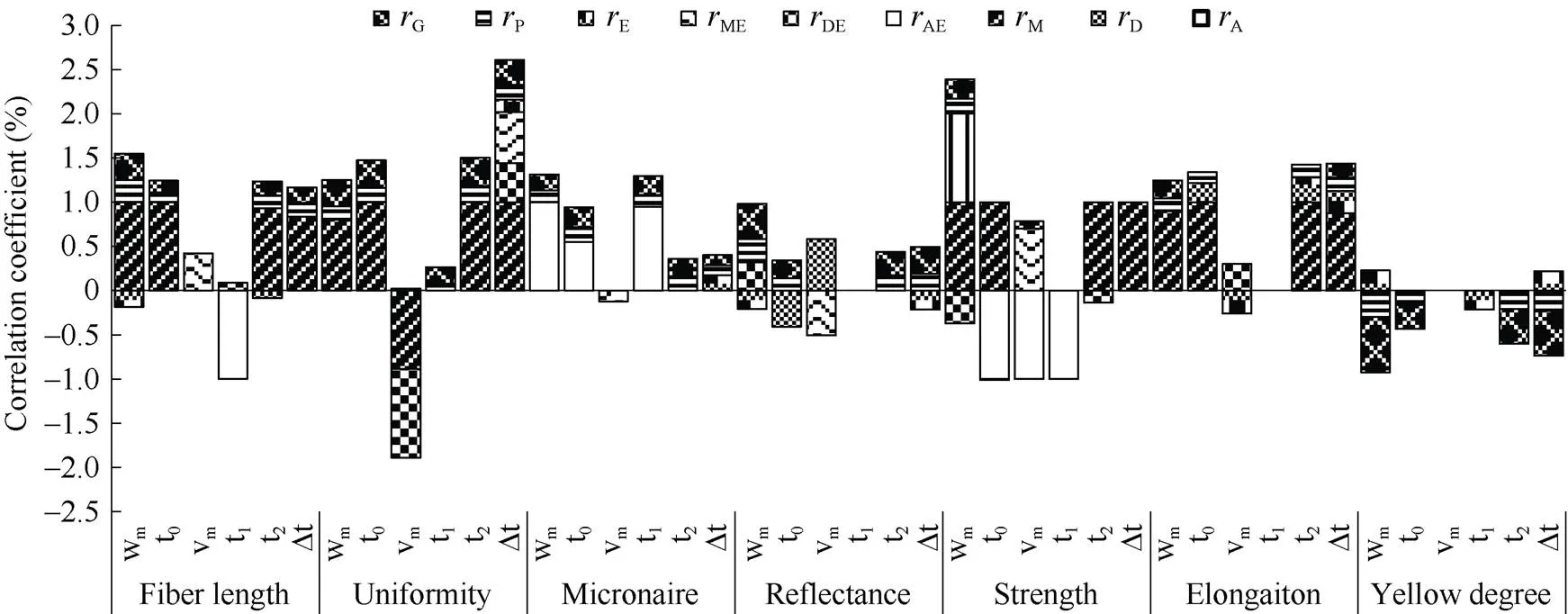
图6 海岛棉单铃重生长特征值与品质性状的相关性
缩写同图1。Abbreviations are the same as those given in Fig. 1.
3 讨论
3.1 利用条件和非条件分析方法, 能够反映不同时期或时段海岛棉铃重发育与产量品质的遗传关系
棉铃是构成产量和品质的基础。棉花产量取决于同化物质运输分配到棉铃的量; 关于海岛棉铃部性状与产量品质性状的关系, 前人已有相应的报道, 通过铃部性状选择产量和品质是可行的[21]; 陆地棉中也有相似的结果, 但这些研究主要反映最终单铃重与产量品质的关系[22-24]。在广义遗传模型的建模原理基础上, Zhu[20]提出了基于混合线性模型的发育遗传模型及相应的统计方法(非条件与条件遗传分析方法), 不但可以获得某一性状从发育起始时间(0)到某一特定发育时刻(t)的累加遗传效应(非条件分析法), 而且还可以获得数量性状在两个任意发育时刻之间的遗传效应净增量(条件分析法), 以了解各遗传分量对性状的相对贡献量, 为数量性状的发育遗传研究提供了新的分析方法和研究思路。本文利用非条件与条件遗传分析方法对不同时期单铃重与产量品质的关系进行研究, 不仅可以了解各时期或时段遗传效应及其与累积遗传效应的相互关系,而且能够了解不同发育时期或时段单铃重与产量品质之间的关系。
3.2 海岛棉棉铃生长发育特点与产量品质的密切关系
棉铃干物质的累积以及在棉铃各部分的分配是棉铃产量和品质形成的基础[4]。棉铃的形态建成早于干物质的累积过程; 而棉铃各部分干物质的积累及分配规律基本相同, 其出现、结束的早晚却因地域、材料的花期、品种熟性、品种遗传特性、栽培方式及植株长势而不同, 说明决定棉铃发育的根本因素在于其遗传基础, 外界条件对其具有一定的影响, 但不能改变其总体趋势[25-28]。本研究海岛棉亲本及其F1代的单铃重均能用Logistics方程进行拟合, 这与大多数学者的观点一致[6,15-17,26]。通过研究海岛棉单铃重的生长特征值与产量品质之间的关系, 能够明确间接选择策略, 前期棉铃缓慢的生长, 将使棉株体内物质分配均衡, 生长稳健; 进入旺增期后同化物质快速往棉铃中输送易形成较高的产量。纤维品质的改良主要取决于单铃重的最终累积量和累积的历期; 纤维形成过程中, 有机物质较早进入铃中, 经历缓慢增长, 使纤维逐渐成熟产生具有优良品质的纤维。不少学者研究发现, 在某一生态区影响棉铃发育的并不是单一因子, 而是2个或多个因子的相互作用[27-28]。不同成铃期的棉铃发育规律可能不同, 本研究对象为海岛棉伏桃, 其他时期棉铃遗传规律还有待进一步研究。
3.3 棉铃发育过程中受到明显的母体植株基因的影响。
对于陆地棉前人做了大量相关试验, 获得较为丰富的研究成果, 而对于海岛棉由于种植区域的限制, 研究基础相对较为薄弱。认为海岛棉单铃重符合加性-上位性模型、加性-显性遗传模型等均有报道[29-30]。郭伟锋等[14,31]对海岛棉开花成铃性状研究发现, 其受到明显的母体效应的影响; 而关于海岛棉铃重的发育遗传规律还未见报道。生物体在发育过程中的表现既受内在遗传因素的调节, 又受外界环境的深刻影响, 同时还存在基因与环境的互作[26]。棉铃及各组分在发育过程中, 母体植株通过提供营养物质, 其性状表现不仅受核基因的影响, 还不同程度受母体基因型的影响[14,31-33], 根据各遗传组分的特点为间接选择提供依据[8]。运用包含环境互作的ADM模型对相关系数进行剖析, 估算不同发育时期或时段海岛棉单铃重及生长发育特点与最终产量品质性状的遗传相关分量, 以揭示生长发育特点与产量品质性状的关系, 这与前人的研究方法有所不同。本研究发现在第10、40、50、60、70天检验到极显著的母体非条件方差, 0~10 d、30~40 d、40~50 d、50~60 d和60~70 d这5个时段检测到极显著母体条件方差; 因此在棉铃发育过程中受到明显的母体效应的影响。
[1] 马富裕. 棉铃发育及纤维品质形成的生态效应与模拟研究. 南京农业大学博士论文, 江苏南京, 2004. Ma F Y. Eco-study and Simulation of Boll Development and Fiber Quality Formation in Cotton. PhD Dissertation of Nanjing Agricultural University, Nanjing, Jiangsu, China, 2004 (in Chinese with English abstract).
[2] 陈兵林, 曹卫星, 周治国. 棉花单铃干物质积累分配的分期动态模拟及检验. 中国农业科学, 2006, 39: 487–493. Chen B L, Cao W X, Zhou Z G. Simulation and validation of dry matter accumulation and distribution of cotton bolls at different flowering stages., 2006, 39: 487–493 (in Chinese with English abstract).
[3] Wanjura D F. Yield accumulation rates and properties development of cotton fiber., 1985, 10: 205–218.
[4] 赵新华, 束红梅, 王友华, 陈兵林, 周治国. 播期对棉铃生物量和氮累积与分配的影响及其与棉铃品质的关系. 作物学报, 2010, 36: 1707–1714. Zhao X H, Shu H M, Wang Y H, Chen B L, Zhou Z G. Effects of sowing date on accumulation and distribution of biomass and nitrogen in cotton bolls., 2010, 36: 1707–1714 (in Chinese with English abstract).
[5] Nibouche S, Goze E, Babin R, Beyo J, Brevault T. Modeling(Hubner) (Lepidoptera: Noctuidae) damages on cotton., 2007, 36: 151–156.
[6] Lemer H R. Development of cotton fruit accumulation and distribution of dry matter., 1976, 68: 205–218.
[7] 吴吉祥, 朱军, 季道藩, 许馥华. 陆地棉产量性状的遗传效应及其与环境互作的分析. 遗传, 1995, 17(5): 1–4. Wu J X, Zhu J, Ji D F, Xu F H. Analysis of genetic effect × environment interactions for yield traits in upland cotton., 1995, 17(5): 1–4 (in Chinese with English abstract).
[8] 王国建, 朱军, 臧荣春, 许馥华, 季道藩. 陆地棉种子品质性状与棉花产量性状的遗传相关性分析. 棉花学报, 1996, 8: 295–300. Wang G J, Zhu J, Zang R C, Xu F H, Ji D F. Analysis of covariance components between seed and agronomy traits in upland cotton., 1996, 8: 295–300 (in Chinese with English abstract).
[9] 张文英, 梅拥军. 陆地棉铃形和纤维品质的遗传和相关研究. 作物学报, 2004, 30: 739–744. Zhang W Y, Mei Y J. Analysis of genetic correlation between boll shape traits and fiber quality traits in upland cotton., 2004, 30: 739–744 (in Chinese with English abstract).
[10] 刘建平, 梅拥军, 张利莉, 胡守林, 郭伟锋, 熊仁次. 零式果枝海岛棉铃部性状和纤维品质的遗传及相关分析. 作物学报, 2005, 31: 1069–1073. Liu J P, Mei Y J, Zhang L L, Hu S L, Guo W F, Xiong R C. Analyses of heredity and correlation between boll traits and fiber quality traits in “0” plant type island cotton., 2005, 31: 1069–1073 (in Chinese with English abstract).
[11] 石春海, 吴建国, 樊龙江, 朱军, 吴平. 不同环境下籼稻糙米重的发育遗传研究. 植物学报, 2001, 43: 603–609. Shi C H, Wu J G, Fan L J, Zhu J, Wu P. Developmental genetic analysis of brown rice weight under different environmental conditions in indica rice., 2001, 43: 603–609 (in Chinese with English abstract).
[12] 梁康迳, 林文雄, 陈志雄, 李亚娟, 梁义元, 郭玉春, 何华勤, 陈芳育. 不同环境下水稻谷粒重的发育遗传分析. 中国农业科学, 2003, 36: 1113–1119. Liang K J, Lin W X, Chen Z X, Li Y J, Liang Y Y, Guo Y C, He H Q, Chen F Y. Development genetic analysis of grain weight under different environmental conditions in rice (L.)., 2003, 36: 1113–1119 (in Chinese with English abstract).
[13] 张莉, 吴建国, Variath M T, 陈国林, 石春海. 不同环境条件下油菜籽饼粕蛋白质含量的发育遗传研究. 中国粮油学报, 2010, 25(9): 45–51. Zhang L, Wu J G, Variath M T, Chen G L, Shi C H. Developmental genetic analysis for protein content of rapeseed meal in different developmental environments., 2010, 25(9): 45–51 (in Chinese with English abstract).
[14] 郭伟锋, 曹新川, 胡守林, 梅拥军. 海岛棉单株成铃性状的发育遗传研究. 棉花学报, 2010, 22: 83–88.Guo W F, Cao X C, Hu S L, Mei Y J. Genetic analysis for developmental behavior of boll number per plant in island cotton., 2010, 22: 83–88 (in Chinese with English abstract).
[15] 陈国林. 油菜籽饼粕氨基酸含量的近红外模型创建及发育遗传研究. 浙江大学博士学位论文, 浙江杭州, 2010. Chen G L. Construction of NIRS Model and Analysis of Developmental Genetics for Amino Acid Contents in Rapeseed Meal. PhD Dissertation of Zhejiang University, Hangzhou, Zhejiang, China, 2010 (in Chinese with English abstract).
[16] 莫旺成, 汤飞宇, 肖文俊. 高品质陆地棉铃-叶系统干物质质量的发育遗传研究. 棉花学报, 2012, 24: 140–146. Mo W C, Tang F Y, Xiao W J. Developmental genetic analysis of dry matter weight of boll-leaf system in upland cotton cultivars with high fiber quality., 2012, 24: 140–146 (in Chinese with English abstract).
[17] 周可金, 裴训武, 江厚旺, 吴宁, 张韪, 夏静, 李爱青. 不同开花期棉铃干物质积累规律研究. 棉花学报, 1996, 8: 145–150.Zhou K J, Pei X W, Jiang H W, Wu N, Zhang W, Xia J, Li A Q. Studies on the dynamics of dry matter accumulation of cotton., 1996, 8: 145–150 (in Chinese with English abstract).
[18] Leffler H R, Hunter J H. Reproductive development and seed quality of cotton cultivars as affected by nitrogen fertilization., 1985, 10: 219–228.
[19] 朱军. 遗传模型分析方法. 北京: 中国农业出版社, 1997. pp 98–112. Zhu J. Analysis of Methods of Genetic Models. Beijing: China Agriculture Press, 1997. pp 98–112 (in Chinese).
[20] Zhu J. Analysis of conditional genetic effects and variance components in developmental genetics., 1995, 141: 1633–1639.
[21] 由宝昌, 刘键萍, 梅拥军, 董安春, 孔向东. 海岛棉铃形与铃重纤维品质关系的研究. 西北农业学报, 1995, 2(1): 27–33. You B C, Liu J P, Mei Y J, Dong A C, Kong X D. A Study of the relations between boll form and boll weight and fibre quality of sea-island cotton., 1993, 2(1): 27–33 (in Chinese with English abstract).
[22] 王伟, 崔秀珍, 李哲. 棉花海陆杂交种主要性状的遗传相关分析. 江苏农业科学, 2009, (3): 51–53. Wang W, Cui X Z, Li Z. Genetic correlation analysis on main traits of hybrids between island and upland cotton., 2009, (3): 51–53 (in Chinese).
[23] 常俊香. 棉花海陆杂交主要经济性状的遗传与杂种优势研究. 河南农业大学硕士论文, 河南郑州, 2007. Chang J X. Studies on Genetic Effects and Heterosis of Main Economic Characters ofÍ. MS Thesis of Henan Agricultural University, Zhengzhou, Hennan China, 2007 (in Chinese with English abstract).
[24] 邢朝柱. 杂交棉遗传效应及基因差异表达与杂种优势关系研究. 中国农业科学院博士学位论文, 北京, 2005. Xing C Z. Genetic Effect of Hybrid Cotton and the Relationship between Gene Differential Expression and Heterosis. PhD Dissertation of Chinese Academy of Agricultural Sciences, Beijing, China, 2005 (in Chinese with English abstract).
[25] 徐敏, 金路路, 王子胜. 辽河流域棉区棉铃发育进程研究. 作物杂志, 2015, (5): 69–73. Xu M, Jin L L, Wang Z S. Research of cotton boll development process in Liaohe basin., 2015, (5): 69–73 (in Chinese with English abstract).
[26] 冯艳波. 棉铃发育过程干物质积累的温度调节效应研究. 南京农业大学硕士论文, 江苏南京, 2007. Feng Y B. Research of Temperature Regulating on Day Matter Accumulation During Developing of Cotton Boll. MS Thesis of Nanjing Agricultural University, Nanjing, China, 2007(in Chinese with English abstract).
[27] 张旺锋, 勾玲, 王振林, 李少昆, 余松烈, 曹连莆, 李伟明. 不同生态棉区棉花单铃重的变化及与气象因子关系的研究. 中国农业科学, 2002, 35, 872–877. Zhang W F, Gou L, Wang Z L, Li S K, Yu S L, Cao L P, Li W M. Changes of cotton single boll weight at different eco-region and correlation with meterological factor., 2002, 35: 872–877 (in Chinese with English abstract).
[28] 陈冠文, 余渝, 朱彪, 李桂玲. 新疆陆地棉棉铃发育特点研究. 新疆农业大学学报, 2003, 26(4): 1–5. Chen G W, Yu Y, Zhu B, Li G L. Study on bolls development characteristics of Xinjiang cotton., 2003, 26(4): 1–5 (in Chinese with English abstract).
[29] 曹新川, 康志钰, 何良荣, 梅拥军, 胡守林. 海岛棉铃部性状的遗传分析. 西北农业学报, 2006, 15(5): 65–68. Cao X C, Kang Z Y, He L R, Mei Y J, Hu S L. Genetic analysis of the boll traits in island cotton., 2006, 15(5): 65–68 (in Chinese with English abstract).
[30] 丁胜, 鲁乃曾, 杨东杰, 张利莉, 梅拥军. 新疆自育海岛棉品种间杂交产量性状的杂种优势分析. 新疆农业科学, 2010, 47(1): 42–46. Ding S, Lu N Z, Yang D J, Zhang L L, Mei Y J. Analysis on heterosis of yield and traits of self-cultivated intervarietal crossing of sea island cotton., 2010, 47(1): 42–46 (in Chinese with English abstract).
[31] 郭伟锋, 曹新川, 胡守林, 梅拥军. 海岛棉开花性状的发育遗传研究. 华北农学报, 2008, 23(增刊2): 173–176. Guo W F, Cao X C, Hu S L, Mei Y J. Genetic analysis for developmental behavior of flowering traits in island cotton., 2008, 23(S2): 173–176 (in Chinese with English abstract).
[32] 吴吉祥, 王国建, 朱军, 许馥华, 季道藩. 陆地棉种子性状直接效应和母体效应的遗传分析. 作物学报, 1995, 21: 659–664. Wu J X, Wang G J, Zhu J, Xu F H, Ji D F. Genetic analysis on direct and maternal effects of seed traits in upland cotton (L.)., 1995, 21: 659–664 (in Chinese with English abstract).
[33] 韩祥铭, 刘英欣, 宋宪亮. 陆地棉新种质纤维品质性状的遗传分析. 作物学报, 2002, 28: 245–248. Han X M, Liu Y X, Song X L. Genetic analysis for fiber traits of new germplasms in upland cotton., 2002, 28: 245–248 (in Chinese with English abstract).
Relationship of stage development of cotton bolls with yield and quality in island cotton
CAO Xin-Chuan, HU Shou-Lin, HAN Xiu-Feng, HE Liang-Rong, and GUO Wei-Feng*
College of Plant Science, Tarim University, Alar 843300, Xinjiang, China
The ADM developmental genetic model containing environmental interaction was used to study the relationship of single boll weight at different developmental stages and developmental characteristics with yield and quality for island cotton parents and F1generations in three years. Selecting single boll weight at different developmental stages had a better effect on the selection of final single boll weight and fiber quality traits in island cotton. When the single boll weed heavy, the number of invalid bolls were reduced and lint in the early stage (10, 20 d), while increased in the later stage (60, 70 d). In the medium term (30, 40, 50 d), the boll weight had a certain selection effect on the final yield per plant. At cotton boll volume expansion period (0–10 d, 10–20 d), the net increment was larger, single boll weight increased, other yield traits decreased, and quality traits such as fiber length, reflectivity and yellowness were improved. At cotton boll filling period (30–40, 40–50, 60–70 d), the boll dry weight increased, main fiber quality traits became worse. The yield per plant was increased by selecting materials with boll weight fast increasing in the growth period later. In the process of boll formation, assimilates entered cotton boll earlier and boll volume increased slowly, so that the fiber matured gradually, forming good quality. The fiber development was obviously affected by maternal effect; therefore, it is important to select suitable maternal parents for improving fiber quality traits.
island cotton; boll weight; developmental heredity; developmental characteristics; genetic correlation
本研究由塔里木大学校长基金项目( TDYZZX201201)和国家自然科学基金项目(31660410)资助。
This study was supported by the Presidential Fund of Tarim University (TDYZZX201201) and the National Natural Science Foundation of China (31660410).
10.3724/SP.J.1006.2020.94051
郭伟锋, E-mail: gwfzky@163.com
E-mail: cxczky@163.com
2019-03-28;
2019-09-26;
2019-10-12.
URL: http://kns.cnki.net/kcms/detail/11.1809.S.20191011.1658.009.html
