遗传因素与男性不育相关性研究进展
2019-12-26马良宏
马 珂 田 稼 马良宏
1.宁夏医科大学临床医学院(宁夏银川 750004);2.宁夏医科大学总医院宁夏人类精子库;3.宁夏医科大学总医院泌尿外科;4.宁夏医科大学生育力保持教育部重点实验室
男性不育是由多种因素(男性精子形成、发育、成熟障碍或精子输出受阻)导致的,育龄夫妇有正常性生活,且未采用任何避孕措施,婚后1年不能受孕的男性因素的总称[1]。目前据WHO统计,世界约有15%育龄夫妇不能生育,男性因素至少占50%[2]。
一、目前男性不育致病因素研究进展
男性不育最常由精子发生障碍引起,临床上称为少精子症或无精子症。男性不育致病因素复杂(见图1),目前已知的主要因素包括遗传因素、抗精子抗体、感染、内分泌因素、疾病、锌缺乏、环境、饮食和吸烟等[3]。但以上各种因素引起少弱精症或无精子症的机制尚未完全阐明。目前导致男性不育的机制主要有以下三种:(1)诸多研究表明这些因素与生精细胞的增殖与凋亡失衡有关。凋亡是细胞主动退化的一种机制,生精过程中细胞凋亡对维持正常的精子生成至关重要,该过程的失衡可能是导致少精子症、非梗阻性无精子症的重要机制之一;(2)一部分因素通过影响生精细胞或精子体内的能量代谢通路,使其体内能量代谢异常,进而引起弱精子症而致男性不育;(3)一部分因素如睾丸炎症等可以通过影响精子从形成到射出的输出的通道阻塞而致梗阻性无精子症导致男性不育。根据这些因素作用机制大致可分为精前性、精子性及精后性。除此之外男性不育患者中约有1/3的患者无明确病因,这部分患者称为特发性男性不育症者。
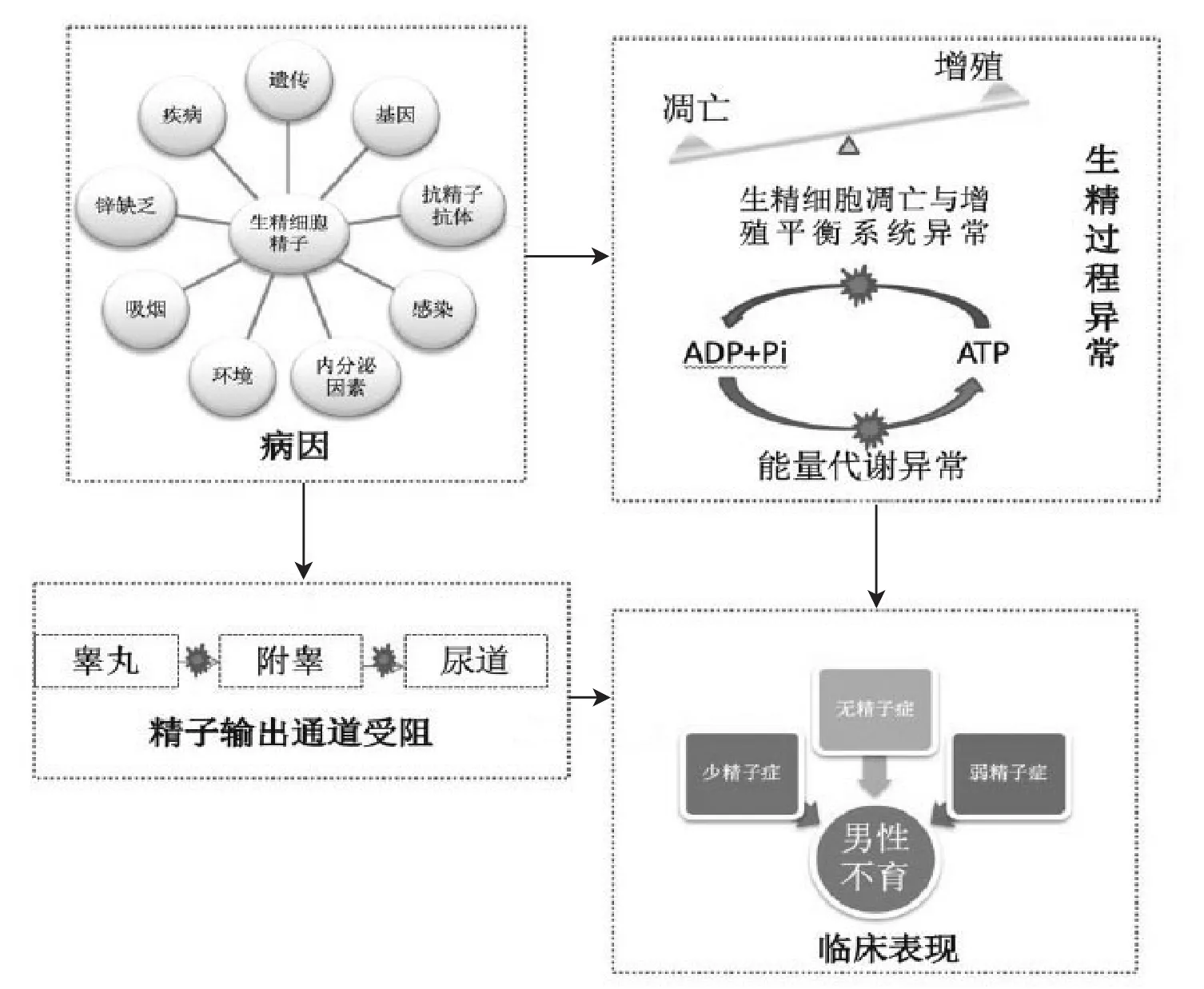
图1 男性不育相关因素致病过程模式图
二、遗传因素与男性不育相关性研究进展
精子的发生是一个受到精细调节的众多基因共同参与的复杂而有序的过程。研究表明,在少精子症、弱精子症及非梗阻性无精子症发生的因素中,遗传因素占到了大约30%,这其中包括基因突变和染色体异常[4]。无精子症(或严重的少精子症)最常见的遗传因素是克氏综合征(47,XXY)、染色体结构异常(例如易位)或Y染色体长臂上AZF区域的微缺失。染色体核型异常,与精子生成相关的基因缺失、突变或表达异常,都可能导致精子生成障碍,形成少精子症、弱精子症或无精子症,最终导致男性不育。
(一)性染色体与男性不育
人类有23对染色体,正常男性性染色体核型为46,XY。目前研究发现人类23对染色体的结构、数量均与生殖发育密切相关。
X、Y染色体作为决定性别发育的重要染色体,染色体上有决定性腺发育成熟和生殖细胞形成的重要基因。X、Y染色体的数量、结构变化往往直接决定了个体性发育成熟以及生殖细胞、胚胎的发育与结局,因此目前X、Y染色体结构、数量变化及其基因仍是学术界研究男性不育的重点(图2)。
人类Y染色体大约长60 Mb,有拟常染色体区(pseudoautosomal regions,PARs)和男性特异性区域(male-specific region of the Y chromosome,MSY)。拟常染色体区位于Y染色体两端,在减数分裂过程中可与X染色体发生重组,男性特异性区域约占Y染色体总长的95%,由异染色质序列以及3类常染色质序列拼接而成[5]。
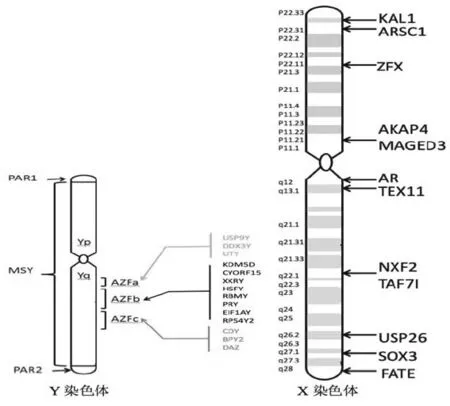
图2 性染色体与男性不育基因
Li等[6]通过对我国东北4659位特发性不育患者的研究,发现其中412位患者存在染色体异常,其中性染色体异常314(6.74%)例;常染色体异常98(2.10%)例,染色体数目和结构异常分别占到259(62.86%)例和153(37.14%)例,其中常见的性染色体核型异常为:46,XX、47,XYY和45,X/46,XY。
1.Y染色体异常与男性不育 男性不育患者中,有10%~15%的非梗阻性无精子症和5%~10%的严重少精子症由Y染色体微缺失引起[7],Y染色体长臂(Yq11)常染色质区存在精子发生相关基因,称为无精子因子(azoospermia factor,AZF)。该因子通常与少精子症有关,尤其是无精子症,是男性不育的第二大常见遗传病因。AZF分为AZFa、AZFb和AZFc 3个区域,现已发现这3个区域内至少有15个与精子发生相关的基因。这些区域含有基因和转录单位,其中大多数呈现特定的睾丸表达,也是目前研究发现与严重少精子症、非梗阻性无精子症发生最常见的染色体微缺失区域[8-10]。
除了已报道的3个区域外,在AZFb和AZFc区之间重复区域也存在STS片段的丢失,有学者将这个区域命名为AZFd区,主要包括sY145位点,AZFd是否存在,目前仍有争议,最新修订的2014年版欧洲男科指南将sY145单独列出[11]。AZF缺失的类型和临床表型丰富多样。中国人无精子症和严重少精子症患者Y染色体微缺失率约为6.59%,其中AZFc区和AZFb+c+sY145区缺失最为常见[12]。
Y染色体微缺失的相对高发生率与其结构相关。它富含高度重复的序列(包括直接重复序列,反向重复序列和回文序列)和高度多态性,这使得长臂特别容易受到通过具有升高序列同源性的区域之间的染色体内同源重组而出现的缺失[13]。
无精子缺失基因(DAZ),位于AZFc区域,是第一个被确定为导致人类无精子症的基因,在睾丸特异性表达[14]。它编码366个氨基酸的蛋白质,含有具有RNA结合功能的RNP/RRM结构域和7个串联重复的14个残基[15]。
RNA结合基序基因(RBMY),位于AZFb区域,编码RNA结合蛋白(同DAZ),在胎儿、青少年和成人睾丸生殖细胞中特异性表达,但在体细胞如Sertoli细胞中不表达。RBMY缺失与生殖细胞分化有关,主要影响早期减数分裂[16]。
USP9Y基因,位于AZFa区域,该基因缺失会导致移码和缺乏外显子7,导致蛋白质水平降低约90%[17]。
最近发现了AZF区一些较小的缺失,例如AZFc区域的拷贝数变异(CNVs)(主要是gr/gr缺失[18]),可能与生精障碍有关,尽管这种情况也可能在正常精子男性中发现[19];b2/b3缺失与精子发生障碍也有关联[20]。Y染色体微缺失还与试管婴儿胚胎质量差、神经精神类疾病、肿瘤存在相关性[21,22]。
2.X染色体异常与男性不育的关系 到目前为止已经发现4种主要的X染色体畸变:(1)在克氏综合征中发现的X染色体的非整倍性 (47,XXY或其他X染色体的镶嵌性)。(2)涉及X染色体的易位,例如非综合性46,XX性睾丸性发育障碍(XX-男性综合征)或X-常染色体易位[23]。 (3)CNVs影响 X 染色体。 (4)点突变破坏X染色体基因[24]。
克氏综合征:是男性不育最常见的核型异常。不育男性中的KS患病率非常高:严重少精子症患者高达5%,无精子症患者高达10%。在80%的病例中,KS的核型是47,XXY。剩余患者呈现更多的非整倍体,例如48,XXXY或49,XXXXY,其特征是更严重的临床形式。过多的性染色体源于精子发生和卵子发生过程中,父母配子的错误分离,主要是不正确的减数分裂造成的[25]。
减数分裂在产生生物多样性和支持物种生存方面发挥着作用,参与染色体交换和DNA重组的许多减数分裂蛋白在进化中高度保守。减数分裂的改变会破坏配子体发生,导致部分或完全的不育。位于X染色体上的TEX11部分缺失,可以导致减数分裂停滞,这意味着前体细胞不能进行适当的减数分裂。这是与X染色体上的基因有关的男性不育的第一个主要遗传原因[26]。
雄激素受体(androgen receptor,AR)基因位于X染色体长臂上(Xq11-12)[27],由8个外显子组成,编码4个功能性区域:N-末端转录区 (外显子1)、DNA结合区(外显子2和3)、铰链区、配体结合区(外显子4-8),编码919个氨基酸残基的蛋白质[28]。突变会引起雄激素不敏感综合征(AIS)并导致少精子症或无精子症[29]。AR基因外显子1是目前研究的热点,外显子1突变会导致其转录激活效能降低,减弱AR对靶基因的表达调节,使精子发生过程改变[30]。约2%~3%的无精子或严重少精子患者是由AR基因突变引起[31]。
ANOS1(以前称为KAL1)位于X染色体短臂上(Xq22,31),在促性腺激素释放激素(GnRH)产生神经元的迁移中起重要作用,突变会引起Kallmann综合征导致低促性腺性功能减退症和精子发生的改变[32]。
RHOX包含3个基因:RHOXF1,RHOXF2和RHOXF2B,它们在人类卵母细胞和雄性生殖细胞中选择性表达[33,34]。RHOXF2/2B突变显著损害了调节HSP70家族转录因子和伴侣等下游基因的能力[35,36]。此外,USP26的变异也与男性不育相关,但有些结果相互矛盾,仍需进一步研究[37]。
(二)常染色体与男性不育
迄今为止,研究发现与少弱精子症、无精子症发生相关的基因不仅存在于性染色体上,在常染色体中也大量存在。
罗伯逊易位:发生于近端着丝粒染色体的一种易位形式。两个近端着丝粒染色体于着丝粒或附近断裂后,导致两条中心染色体长臂融合 (D组染色体:13,14,15 和 G 组染色体:21,22eY)[38],形成一条由长臂构成的衍生染色体。通常为双着丝粒,丢失了相应的短臂。在载体表型中无明显变化,但可能导致睾丸体积和睾酮水平降低,影响精子发生或导致胎儿死亡[39]。
CFTR基因:位于 7号染色体上(7q31.1-31.2),纯合子形式引起囊性纤维化,是高加索人群中最常见和最严重的常染色体隐性疾病之一。CFTR表达的突变会引起男性先天性双侧输精管缺失(CBAVD),导致阻塞性无精子症[40]。
INSL3-RXFP2基因:INSL3的编码基因位于19号染色体上,由两个带内含子的外显子组成。在患有隐睾症的男性中,与这两个基因突变相关的患病率约为4%~5%[41]。
SYCP3基因:位于12号染色体上(12q23),在睾丸中特异性表达。它编码236个氨基酸的蛋白质,具有两个卷曲螺旋结构域,是一种与生殖细胞减数分裂过程中涉及的突触相关的DNA结合蛋白。突变引起减数分裂异常,造成卷曲螺旋结构域腺苷碱基的缺失,导致移码突变并出现早期终止密码子,导致结构域不完整。对突变进行功能分析,结果显示变短的SYCP3几乎没有功能性蛋白质-蛋白质相互作用[42]。SYCP3突变通过男性减数分裂停滞和女性复发性妊娠引起无精子症[43]。
此外,KLHL10突变会损害少精子症患者的同源二聚化[44];AURKC的纯合子突变产生大头多倍体精子[45],均与男性不育相关。
(三)线粒体DNA异常与男性不育
精子活力是男性生育力的主要决定因素之一,精子中的线粒体是提供精子活力必需的细胞器。人类mtDNA仅由没有内含子的外显子组成。由于高转换率,没有校对或有效的DNA修复系统,缺乏组蛋白保护或DNA结合蛋白,mtDNA突变率比核DNA高10~100倍。线粒体DNA突变可能会减少ATP产生从而降低男性生育能力。线粒体DNA拷贝数也是线粒体遗传变异和病理学的重要因素[46]。
研究表明,精液参数改变的患者mtDNA拷贝数显著增加,mtDNA完整性显著降低,活性氧(ROS)产生增加[47]。mtDNA的大规模缺失可能导致不完整的呼吸链的产生,这可能导致男性生育能力和精子活力的下降[48]。
最近有研究显示mtDNA 4,977-bp缺失与精索静脉曲张引起的不育相关,但有必要进行进一步研究[49]。
三、结语
男性不育的遗传学研究是一个快速发展的领域。寻找与男性不育致病相关的染色体、基因变化越来越受到研究界的重视。过去几年的遗传学关联研究,基因突变筛查,动物模型和基础研究清楚地表明生精障碍和男性不育的高发病率。相信随着研究的不断深入,将会有更多的染色体、基因异常在男性不育致病方面的作用与机制被发现。这也将为进一步阐明男性不育症发病的遗传因素提供重要的理论依据,同时也为男性不育症的基因靶向治疗提供思路。
