瓦氏黄颡鱼饥饿后再投喂的生长及转氨酶活性的个体差异
2019-11-19安徽农业大学动物科技学院安徽合肥230036农业农村部华南水产与畜禽饲料重点实验室广东恒兴饲料实业股份有限公司广东湛江524094安徽农业大学动物科技学院安徽合肥230036
(安徽农业大学动物科技学院,安徽 合肥 230036) (农业农村部华南水产与畜禽饲料重点实验室,广东恒兴饲料实业股份有限公司,广东 湛江 524094) (安徽农业大学动物科技学院,安徽 合肥 230036)
鱼类在自然水体中有时会面临饥饿胁迫[1],一定时间的饥饿后鱼类会发生失重现象,这种失重具有个体差异[2, 3],即某些个体失重较多,失重率较高,而某些个体则相反。鱼类在恢复正常摄食后会继续生长,其生长速度会受到饥饿时间、鱼体初始体重等因素影响[2, 3],而生长是否会受到失重率的影响,即是否饥饿时失重越多,恢复摄食后的生长就越快,对这一问题还未见相关研究报道。因此,探讨这一问题,可以更好地了解鱼类遭受胁迫后的生长特点,也可为鱼类生长性状的选育提供参考。
鱼类饥饿后的再生长速度与蛋白质在鱼体中的沉积密切相关,即与鱼体氨基酸的代谢密切相关[4, 5]。谷丙转氨酶和谷草转氨酶是重要氨基酸转移酶,在鱼体肝脏中含量丰富,推测鱼类饥饿后的再生长可能与转氨酶的活性密切相关,因此探讨鱼类饥饿后再生长的转氨酶活性变化个体差异,可更好地了解鱼类的生理特点。为此,笔者以我国重要的名特养殖鱼类瓦氏黄颡鱼 (PelteobagrusvachelliRichardson) 为对象,探讨了饥饿后的再生长及生理指标变化的个体差异。
1 材料与方法
1.1 试验材料与饲养设备
饲养设备为室内养殖系统,其主要设备为18只单个规格为40cm×20cm×30cm(水深20cm)的玻璃水族箱。瓦氏黄颡鱼由安徽省淮南市黄颡鱼良种场提供,运回后2%食盐水消毒。随机取试验鱼36尾,分别放入上述18个水族箱中,每箱2尾,暂养1周,暂养期间每天饱食投喂。
1.2 试验方法
试验开始,将36尾鱼取出,随机取18尾,分别称重,再分别放入18个水族箱中。初始体重为7.72~25.04g。自然室温(试验期间自然水温23.4~28.2℃),自然光照。试验期间,每周换水1次,1次,以虹吸法取出老水,再注入镇放后的新水。试验分2个阶段,首先将鱼饥饿2周后称重(精确到0.01g),从第3周开始投喂,每日中午12:40将过量摇蚊幼虫放入水族箱,24h后收集未吃完的残饵。投喂2周后,将鱼饥饿1d,称量体重,测定内脏、肝脏重,再测定肝脏谷草转氨酶(GOT)和谷丙转氨酶(GPT)活性。采用南京建成生物工程研究所试剂盒测定肝脏中谷丙转氨酶和谷草转氨酶活性,严格按照使用说明书中的方法进行。
1.3 指标计算及数据分析
失重率(WLR):
特定湿重生长率(SGR):
摄食率(FR):
饵料转化效率(FCE):
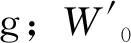
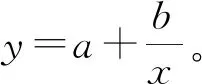
利用数据处理软件Excel 2007进行数据处理,使用SPSS V22.0软件检验回归关系式的显著性,P< 0.05为显著性标准。
2 结果与分析
2.1 恢复投喂后湿重特定生长率、摄食率、饲料转化效率与失重率的关系
表1所示为瓦氏黄颡鱼饥饿处理和恢复摄食阶段鱼体生长与饲料利用基本情况。
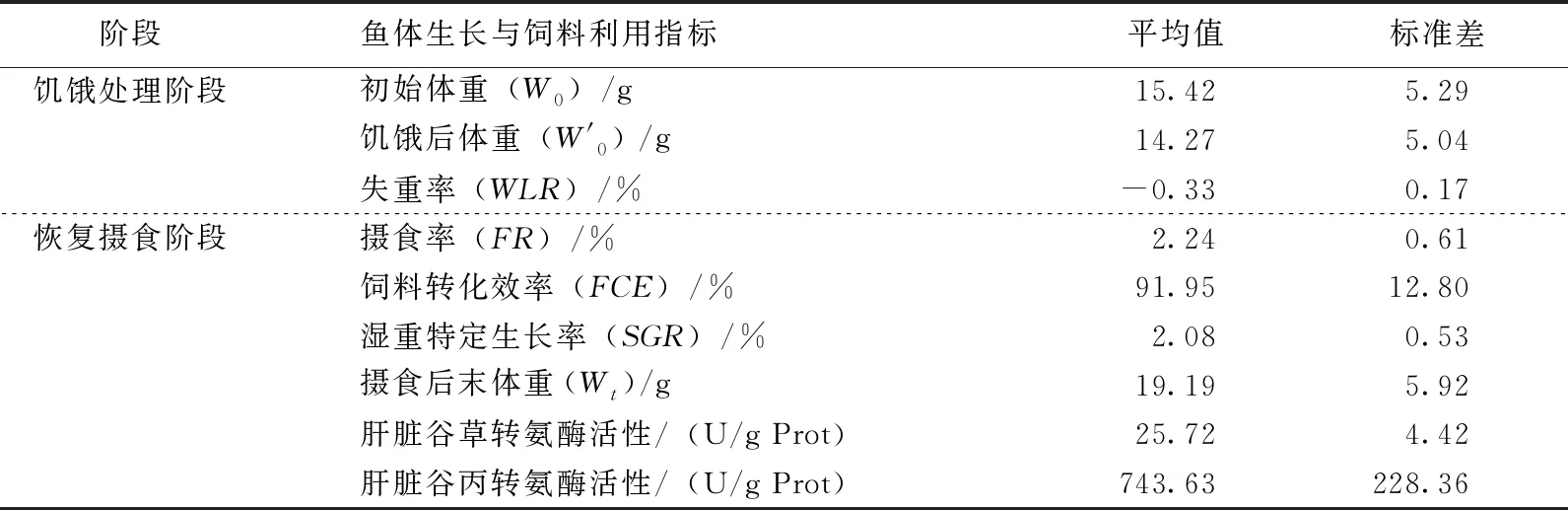
表1 瓦氏黄颡鱼饥饿处理和恢复摄食阶段鱼体生长与饲料利用基本情况
由图1及表2可知,瓦氏黄颡鱼个体湿重特定生长率、摄食率及饲料转化效率与失重率之间无显著相关关系(P>0.05)。
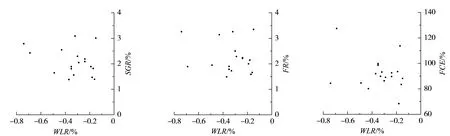
图1 瓦氏黄颡鱼恢复投喂后湿重特定生长率、摄食率、饲料转化效率与失重率的关系

yNabR2P湿重特定生长率(SGR)181.772-0.9140.0840.243摄食率(FR)182.023-0.6670.0340.465饲料转化率(FCE)1885.125-20.4990.0730.278
注:回归关系式均为y=a+bx,x为失重率(WLR),N为样本数。
2.2 恢复投喂后湿重特定生长率、摄食率、饲料转化效率与初始体重的关系
由图2及表3可知,瓦氏黄颡鱼个体特定湿重生长率、摄食率与初始体重显著负相关(P<0.05),而饲料转化效率与初始体重显著正相关(P<0.05)。
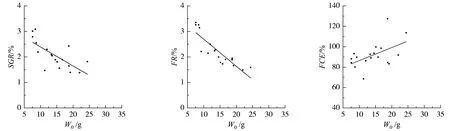
图2 瓦氏黄颡鱼恢复投喂后湿重特定生长率、摄食率、饲料转化效率与初始体重的关系

yNabR2P湿重特定生长率(SGR)183.165-0.0710.495<0.05摄食率(FR)183.857-0.1450.818<0.05饲料转化率(FCE)1871.2651.3410.308<0.05
注:回归关系式均为y=a+bx,x为初体重(W0),N为样本数。
2.3 恢复投喂后谷丙转氨酶活性、谷草转氨酶活性与湿重特定生长率、摄食率及饲料转化效率的关系
由图3及表4可知,瓦氏黄颡鱼谷丙转氨酶活性与湿重特定生长率、摄食率及饲料转化效率之间无显著关系(P>0.05);谷草转氨酶活性与饲料转化效率之间无显著关系(P>0.05),但与湿重特定生长率及摄食率显著正相关(P<0.05)。
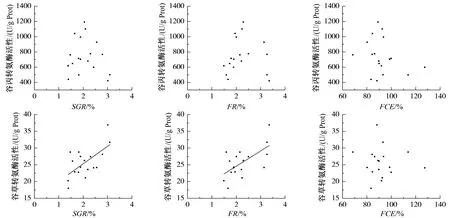
图3 瓦氏黄颡鱼恢复投喂后谷丙转氨酶、谷草转氨酶活性与湿重特定生长率、摄食率、饲料转化率的关系
yxNabR2P谷丙转氨酶活性湿重特定生长率(SGR)18857.090-54.6620.0160.615摄食率(FR)18735.1003.8000.00010.968饲料转化率(FCE)181170.300-4.6400.0680.297谷草转氨酶活性湿重特定生长率(SGR)1814.8355.2440.398<0.05摄食率(FR)1815.4924.5570.399<0.05饲料转化率(FCE)1827.372-0.0180.0030.837
注:回归关系式均为y=a+bx,N为样本数。
3 讨论
研究结果表明,个体饥饿后再摄食,其生长率与饥饿期间的失重率无关,而软形鲳鯵(Trachinotusovatus)[6]、厚颌鲂(Megalobramapellegrini)[7]稚鱼饥饿时的失重率越高,饥饿后再摄食的生长率则越高。可能的原因是后两者的失重率与饥饿时间有关,饥饿时间越长,失重率越高。饥饿时间相同,失重率差异来自于鱼类的个体生理特点差异,而这种差异与后续生长没有显著相关性。研究中,这种再摄食后的生长速度与饥饿前的鱼体初始体重显著负相关,初始体重越小,摄食率越高,虽然饲料转化效率下降,但总体仍表现为生长率越高。鱼体生长速度与体重间存在显著的负相关[3,8,9],研究结果显示,即使经过饥饿胁迫,这种关系仍然存在。
谷丙转氨酶与谷草转氨酶主要存在于动物体的肝脏中,是与蛋白质代谢相关的重要代谢酶,其活性反映蛋白质代谢强度,活性越高则蛋白质代谢强度越高[10]。研究表明,肝脏中转氨酶活性随饲料蛋白质水平的提高而增强[11, 12],研究中不存在饲料蛋白质水平差异,但由于个体初始体重不同,个体间摄食率与饲料转化效率存在差异,致使不同个体的蛋白质代谢强度也不同。结果显示,肝脏谷丙转氨酶活性与湿重特定生长率、摄食率及饲料转化效率之间无显著关系;而谷草转氨酶活性与湿重特定生长率、摄食率显著正相关,即摄食率、生长率越高的个体,谷草转氨酶活性越高,这与团头鲂(Megalobramaamblycephala)周期性饥饿研究[13]中的现象一致。2种转氨酶活性变化趋势不同的原因可能是谷丙转氨酶在肝脏中活性最高,能够满足氨基酸酶促降解中转氨基作用(主要发生在肝、肾)的需要,而谷草转氨酶活性在肝脏内次之,当蛋白质代谢增强后,因转氨基作用的需要,肝脏中谷草转氨酶活性随之提高。但谷草转氨酶活性变化与摄食之间的具体作用关系比较复杂,其内部的生理学机制亟待进一步研究。
