长时间不同频率电刺激对C2C12肌管细胞代谢指标和基因表达的影响
2019-10-18孙景权上官若男李松波严翊谢敏豪
孙景权 上官若男 李松波 严翊 谢敏豪



摘 要:探究长时间不同频率EPS对C2C12肌管细胞代谢指标和基因表达的影响,试图建立EPS诱发C2C12肌管细胞TG堆积和线粒体功能增强的细胞模型。方法:采用4种EPS方案诱发C2C12肌管细胞收缩,实验分为5组:对照组(C组)、30 V+2 ms+0.5 Hz组(0.5 Hz组)、30 V+2 ms+1 Hz组(1 Hz组)、30 V+2 ms+5 Hz组(5 Hz组)、30 V+2 ms+10 Hz组(10 Hz组)。比色法检测细胞内TG、乳酸和ATP含量,RT-PCR方法检测细胞中IL-6、MyHCⅠ、MyHCⅡa、MyHCⅡb、MyHCⅡx、Cyt C和PGC-1α等mRNA含量。结果:1)与C组相比,EPS后即刻,频率0.5~10 Hz组乳酸含量非常显著增加(P<0.01);频率1 Hz和10 Hz组细胞中ATP含量均显著降低(P<0.05),其中频率0.5 Hz和5 Hz组细胞中ATP含量非常显著降低(P<0.01)。2)与C组相比,频率0.5 Hz和1 Hz组MyHCⅠ和MyHCⅡb的mRNA含量显著增加(P<0.05),频率5 Hz组MHC4种亚型mRNA含量变化不明显(P>0.05),频率10 Hz组MyHCⅠ、MyHCⅡa和MyHCⅡb亚型mRNA含量非常显著降低(P<0.01),而MyHCⅡx亚型mRNA含量非常显著增加(P<0.01)。3)与C组相比,频率0.5~10 Hz组PGC-1α和Cyt C mRNA含量均显著增加(P<0.05),同时EPS增加了C2C12细胞中IL-6 mRNA的含量(P<0.05)。4)與C组相比,4 d EPS方案停止后12 h,各组肌管细胞中TG含量均没有出现变化;4 d EPS方案停止后24 h,0.5 Hz组和1 Hz组肌管细胞中TG含量增加但无显著性(P>0.05);4 d EPS方案停止后36 h,0.5 Hz组和5 Hz组细胞中TG含量显著增加(P<0.05)。结论:EPS方案(电压14 V、频率1 Hz、单次时长2 ms和每天2 h共2 d,紧接着,电压30 V、频率0.5 Hz或1 Hz或5 Hz、单次时长2 ms和每天3 h共2 d,EPS停止后36 h)能够明显诱发C2C12肌管细胞TG含量堆积和线粒体功能增强。
关键词:EPS;C2C12 肌管细胞;线粒体;TG;电脉冲;模型
中图分类号:G 804.8 学科代码:040302 文献标识码:A
Abstract:Objective: To explore the effect of long time and different frequency electrical stimulation on the metabolic index and gene expression in C2C12 myotubes, so as to establish a cell model for EPS-induced TG accumulation and the increase of mitochondrial function in C2C12 myotubes. Methods: Four protocols of EPS were used to induce contraction of C2C12 myotubes. The experiment was divided into five groups: control group (C group), 30V 2ms 0.5Hz group (0.5Hz group), 30V 2ms 1Hz group (1Hz group), 30V 2ms 5Hz. Group (5Hz group), 30V 2ms 10Hz group (10Hz group). Colorimetric assay was used to detect intracellular TG, lactate, and ATP content. RT-PCR was used to detect mRNA levels of IL-6, MyHCⅠ, MyHCⅡa, MyHCⅡb, MyHCⅡx, Cyt C, and PGC-1α. Results: 1) Compared with C group, the lactate content in the 0.5Hz-10Hz group was significantly increased (P<0.01) immediately after stimulation, and the ATP content in cells was significantly decreased in the 1Hz and 10Hz groups (P<0.05). However, the ATP content in cells at 0.5 Hz and 5 Hz groups significantly decreased (P<0.01). 2) Compared with C group, MyHCⅠ and MyHCⅡb were significantly increased in the 0.5 Hz/1 Hz group (P<0.05), and the four myosin heavy chain isoforms exhibited no change in 5 Hz group (P>0.05). The mRNA content of MyHCⅠ, MyHCⅡa, and MyHCⅡb subtypes was significantly decreased (P<0.01) in the 10 Hz group, while the mRNA levels of MyHCⅡx isoform was significantly increased (P<0.01). 3) Compared with C group, the mRNA content of PGC-1α and Cytochrome C were significantly increased (P<0.05) in 0.5-10 Hz group, and the mRNA content of IL-6 was significantly increased by EPS in C2C12 cells (P<0.05). 4) Compared with C group, TG content in EPS groups did not change in 12h after 4d electrical stimulation, and the TG content in 0.5Hz and 1Hz groups were increased in 24h after 4d electrical stimulation but there was no significant difference (P>0.05); however, the TG content in 0.5 Hz and 5 Hz groups were significantly increased in 36h after 4d electrical stimulation (P<0.05). Conclusion: the EPS protocols (Voltage 14V, a single stimulation duration 2ms, frequency 1Hz, a stimulation time of 2h, a total 2 days, followed by, voltage 30 V, a single stimulation duration 2ms, frequency 0.5 Hz or 1 Hz or 5Hz, a stimulation time of 3 h, and a total 2 days) have induced TG accumulation and mitochondrial function enhancement in C2C12 myotubes.
Keywords:EPS; C2C12 myotubes; mitochondria; TG; electrical pulse stimulation; model
许多研究表明,长期耐力训练能够增加骨骼肌细胞中甘油三酯(triglyceride,TG)的含量和线粒体氧化脂肪酸能力,这与耐力性项目运动员骨骼肌耐力水平的增加有直接关系[1-3]。在长时间低强度运动中,骨骼肌收缩所需要的能量中有90%来源于游离脂肪酸的氧化代谢[4]。肌细胞内TG作为局部的高密度能量,其可以通过脂解作用释放脂肪酸来为骨骼肌收缩提供能量[5-6]。不论是耐力性项目运动员[7-8]、Ⅱ型糖尿病患者[2]还是老年人人群(60~75岁)[1,9],长时间中等强度的运动锻炼均可以提升骨骼肌细胞的TG含量、骨骼肌胰岛素敏感性、Ⅰ型骨骼肌纤维脂肪酸利用能力及骨骼肌整体氧化能力,但是运动诱发骨骼肌出现这些健康效应的分子机制还不清楚。通过研究骨骼肌细胞对收缩刺激的适应性变化,是阐明运动诱发骨骼肌产生健康效应机制的主要方法之一[10]。
一个科学的体外细胞培养模型是研究生物學现象机制的重要工具。在细胞培养中,电脉冲刺激(electrical pulse stimulation,EPS)能够加快骨骼肌细胞(肌管)分化,它能够替代神经电信号激活骨骼肌纤维[11-12]。目前,一些研究中已经出现使用EPS诱导细胞收缩的模型[13]。1997年,有研究发现,采用EPS激活培养的骨骼肌细胞后,其能够引起骨骼肌细胞产生收缩[11]。随后在2007年,有研究发现EPS能够加速骨骼肌细胞肌节的形成[12]。近几年,许多研究发现EPS能够诱发骨骼肌细胞相关基因表达和代谢特征的变化。例如一次性长时间EPS(14 V,1 ms,50 Hz,24 h)能够显著增加氧化型MyHCⅠ和MyHCⅡx,同时增加PGC-1α、Cyt C和中链乙酰辅酶A脱氢酶(medium chain acyl-coA dehydrogenase,MCAD)蛋白表达量,即显著增加骨骼肌氧化功能[14]。一次性短时间高频EPS(电压30 V,单次时长200 ms,频率100 Hz,持续5~60 min)能够显著增加骨骼肌细胞脱氧葡萄糖摄入量和乳酸产生量,同时降低磷酸肌酸含量;而一次性长时间低频EPS显著增加骨骼肌细胞葡萄糖和脂肪酸氧化能力,丙酮酸脱氢酶复合物4、IL-6、Cyt C、肉碱棕榈酰转移酶B和MyHCⅠ蛋白和mRNA表达量显著增高[13]。但是,目前还鲜少见到长时间EPS诱发骨骼肌细胞收缩进而导致TG堆积和线粒体功能增强的成功建立的细胞模型。
综上所述,本研究采用长时间不同频率的EPS方案诱发C2C12肌管细胞收缩,检测C2C12细胞中TG含量、乳酸含量、肌细胞收缩蛋白和线粒体相关功能蛋白含量变化情况,试图建立EPS诱发C2C12肌管细胞收缩并伴随TG堆积和线粒体功能增加的细胞模型,为进一步研究长期运动锻炼增加肌细胞中TG含量并伴随线粒体功能增强的机制提供实验基础。
1 材料和方法
1.1 细胞培养
小鼠成肌细胞细胞株C2C12(CRL-1772),接种于含培养基的100 mm培养皿中,在37 ℃、5%CO2恒温培养箱中培养。培养基包含高糖DMEM(4.00 mM L-谷氨酰胺,4 500 mg/L葡萄糖,无丙酮酸钠,HyClone,货号SH30022.01,美国犹他州洛根市)、10%胎牛血清(Ausvin,货号VS500T,澳大利亚)、1%青链霉素混合液(货号KGY0023,购自江苏凯基生物技术有限公司)。使用0.25%胰蛋白酶-EDTA消化液(货号KGY0012,购自江苏凯基生物技术有限公司)进行细胞消化。对于本次实验,细胞被铺入6孔板(Corning,New York,USA)。当6孔板中细胞融合到90~95%时,将C2C12细胞培养基更换为分化培养基(高糖DMEM,2%马血清,1%青链霉素混合液)。每24 h更换一次分化培养基。细胞分化4~6 d后,分化完成的肌管细胞被用于EPS处理。
1.2 EPS方案
本研究使用仪器C-Pace EP culture pacer(IonOptix,Dublin,Ireland)产生EPS使肌管细胞收缩。此仪器能够发出双极性脉冲到悬挂在培养基中的C-dish碳电极上。EPS方案如下:EPS共持续4 d,第1天和第2天:电压14 V,单次刺激时长2 ms,频率1 Hz,电刺激总时间2 h;第3天和第4天:电压30 V,单次刺激时长2 ms,频率分别为0.5 Hz、1 Hz、5 Hz和10 Hz,电刺激总时间3 h(见表1)。基于此,本研究实验细胞被分为5组:对照组(C组)、30 V+2 ms+0.5 Hz组(0.5 Hz组)、30 V+2 ms+1 Hz组(1 Hz组)、30 V+2 ms+5 Hz组(5 Hz组)、30 V+2ms+10 Hz组(10 Hz组),其中C组为未进行电刺激组。每1组均包含6个孔的细胞作为检测对象(n=6)。EPS前1 h每孔更换2 mL新配分化培养基。EPS每天进行1次,共持续4 d。根据以往研究[14],在最后一次EPS后3 h收集细胞提取总RNA进行MHC亚型、IL-6、PGC-1α、Cyt C等mRNA含量测试。另外,分别在最后一次EPS后即刻、12 h、24 h和36 h后收集细胞,使用裂解液后提取蛋白质和进行TG、ATP等含量的测试。
1.3 细胞中TG测试
肌管细胞中总TG含量采用组织甘油三酯酶法测定试剂盒(组织细胞)(E1013,购自北京普利莱(APPLYGEN)基因技术有限公司)进行测试,操作步骤按照试剂盒说明书进行。最后使用酶标仪(benchmark plus microplate spectrophotometer,BIO-RAD,California,USA) 在550 nm波长下测量吸光度。根据标准样品读数绘制标准曲线,并计算样品中TG浓度。样品中相对TG含量使用细胞总蛋白浓度进行校准。
1.4 细胞中乳酸含量测试
细胞中乳酸含量测定使用乳酸比色法测定试剂盒(货号:#K607-100,BioVision,Milpitas CA,USA),测试方法依照说明书进行。最后,使用酶标仪(BIO-RAD,California,USA)在波长570 nm下读取吸光度。根据标准曲线计算出样品中乳酸含量(nmoL),根据公式计算:
样品乳酸浓度=每孔中乳酸含量(nmoL)/每孔中样品体积(μL)×样品稀释比例。(1)
样品中相对乳酸含量使用细胞总蛋白濃度进行校准。
1.5 细胞中ATP含量测试
细胞中ATP含量测试使用ATP检测试剂盒(货号:S0026,购自上海碧云天生物技术有限公司),测试方法依照说明书进行。最后,在检测孔内加上20 μL样品或标准品,迅速用微量移液器混匀,至少间隔2 s后,用GloMaxR 20/20 Luminometer化学发光检测仪(货号:E5311,美国Promega-GloMax Promega GloMax)测定RLU值;根据标准曲线计算出样品中ATP的浓度,相对ATP含量用样品中蛋白浓度校准。
1.6 蛋白质提取及浓度测定
6孔板中每孔加细胞裂解液(货号:110000-L2,购自北京赛驰生物科技有限公司) 200 μL,用细胞刷将细胞刮下,收集在1.5 mL离心管中,置于冰上。用枪头反复吹吸细胞沉淀,8~10次,以充分悬浮细胞。旋涡剧烈震荡1 min,静置15 min。期间取出震荡2~3次,每次30 s。4 ℃离心,12 000 g,15 min,将上清分装至预冷离心管中,-80 ℃保存备用。总蛋白浓度测定采用Thermo Scientific公司的BCA蛋白定量分析试剂盒(Prod# 23225,Life technologies,Eugene,Oregon),采用BSA作为标准液制作标准曲线,测试方法依照说明书进行。
1.7 实时荧光定量RT-PCR
细胞中总RNA提取采用TaKaRa MiniBEST Universal RNA Extraction Kit(货号9767,Takara),操作步骤按照试剂盒说明书进行。然后,使用微量分光光度计(NanoDrop 2000)测定总RNA浓度,纯度要求:A260/A280比值在1.9~2.0,RNA浓度=A260×稀释倍数×0.04 μg/μL。提出的mRNA使用PrimeScriptTM RT Master Mix (Perfect Real Time)(货号:RR036A,Takara)来进行逆转录。然后Bio-RAD上机,反转录程序为37 ℃15 min,85 ℃5 s,4 ℃无穷。
实时荧光定量RT-PCR。使用SYBRR Premix Ex TaqTM Ⅱ(Tli RNaseH Plus)(货号:RR820A,Takara)试剂盒。按照试剂盒说明书要求配制20 μL的反应体系,在Applied Biosystems7500 Fast Real-Time PCR System仪器上进行定量分析。两步法 PCR 扩增标准程序:95 ℃预变性30 s;95 ℃ 5 s,60 ℃ 30~34 s的PCR反应设置40个循环;之后熔解曲线程序95 ℃ 15 s,60 ℃15 s,95 ℃ 60 s,并且用核糖体18S1 RNA作为内参基因校准。PGC-1α、MyHCⅠ、MyHCⅡa、MyHCⅡx、MyHCⅡb、Cyt C、IL-6和18S rRNA mRNA采用SYBR green方法进行检测。引物购自生工生物工程(上海)股份有限公司,序列见表2,并且用18S rRNA进行内参基因校准。定量方法用2—△△Ct表示基因的表达量。计算公式为:
△△Ct=[Ct(实验组目的基因)-Ct(18S1)]-[Ct(对照组目的基因)-Ct(18S1)](2)
1.8 数据处理方法
数据以平均数±标准误(X±SEM)表示,采用SPSS18.0统计软件进行处理。组间比较采用单因素方差分析,显著水平取P<0.05,非常显著水平取P<0.01。作图软件采用Excel 2010和GraphPad Prism 5。
2 结果
2.1 一次性EPS对C2C12肌管细胞中乳酸和ATP含量的影响
为了检测EPS诱导肌管细胞收缩后细胞中乳酸和ATP含量的变化,分化4~6 d的肌管细胞先进行2 d EPS(电压14 V,单次刺激时长2 ms,频率1 Hz,EPS总时间2 h/d),第3 d进行一次性高电压不同频率刺激(电压30 V,单次刺激时长2 ms,频率分别为0.5 Hz、1 Hz、5 Hz和10 Hz,EPS总时间3 h)。EPS后即刻收集细胞检测乳酸和ATP含量,采用单因素方差分析事后检验方法计算结果。
如图1所示,与C组相比,一次性的电壓30 V、频率0.5 Hz和10 Hz、单次时长2ms、共3 h的EPS均能显著升高刺激即刻时C2C12肌管细胞中乳酸含量(P<0.05),同时发现电压30 V、频率1 Hz和5 Hz、单次时长2 ms、共3 h的EPS均能非常显著升高刺激即刻时C2C12肌管细胞中的乳酸含量(P<0.01)。如图2所示,与C组相比,一次性的电压30 V、频率0.5~10 Hz、单次时长2 ms共3 h的EPS均能够非常显著降低C2C12细胞中的ATP含量(P<0.01)。
2.2 不同EPS方案对C2C12肌管细胞中MHC亚型mRNA含量的影响
本研究观察这几种EPS方案是否能够改变C2C12肌管细胞中4种MHC亚型的mRNA含量。如图3所示,与C组相比,电压30 V、频率0.5 Hz、单次时长2 ms共3 h的EPS有增加氧化型蛋白重链Ⅰ型(P<0.05)和快肌纤维标志性肌球蛋白重链Ⅱb型mRNA含量的趋势,但未见显著性(P<0.05)。电压30 V、频率1 Hz、单次时长2 ms共3 h的EPS能够显著增加氧化型蛋白重链I型(P<0.05)和快肌纤维标志性肌球蛋白重链Ⅱb型mRNA含量(P<0.01)。而电压30 V、频率5 Hz、单次时长2 ms共3 h的电刺激对骨骼肌MHC4种亚型mRNA含量的影响不明显。然而,电压30 V、频率10 Hz、单次时长2 ms共3 h的EPS非常显著降低MyHCⅠ、MyHCⅡa和MyHCⅡb亚型mRNA含量(P<0.01),非常显著增加MyHCⅡx亚型mRNA含量(P<0.01)。

2.3 不同EPS方案对C2C12肌管细胞中线粒体相关功能蛋白mRNA含量的影响
本研究检测了EPS对线粒体相关蛋白Cyt C、PGC-1α和IL-6mRNA含量的影响。如图4A所示,与C组相比,电压30 V、频率0.5~10 Hz、单次时长2 ms的EPS方案均能够显著增加C2C12肌管细胞中Cyt C mRNA表达量(P<0.05)。如图4B所示,与C组相比,电压30 V、频率1~10 Hz、单次时长2 ms的EPS方案均能够显著增加C2C12肌管细胞中PGC-1αmRNA含量(P<0.05),而0.5 Hz组有增加趋势但无显著性(P>0.05)。如图4 C所示,与C组相比,随着频率的增加,C2C12细胞中IL-6 mRNA含量出现增加趋势,至10 Hz呈现非常显著性增加(P<0.01)。

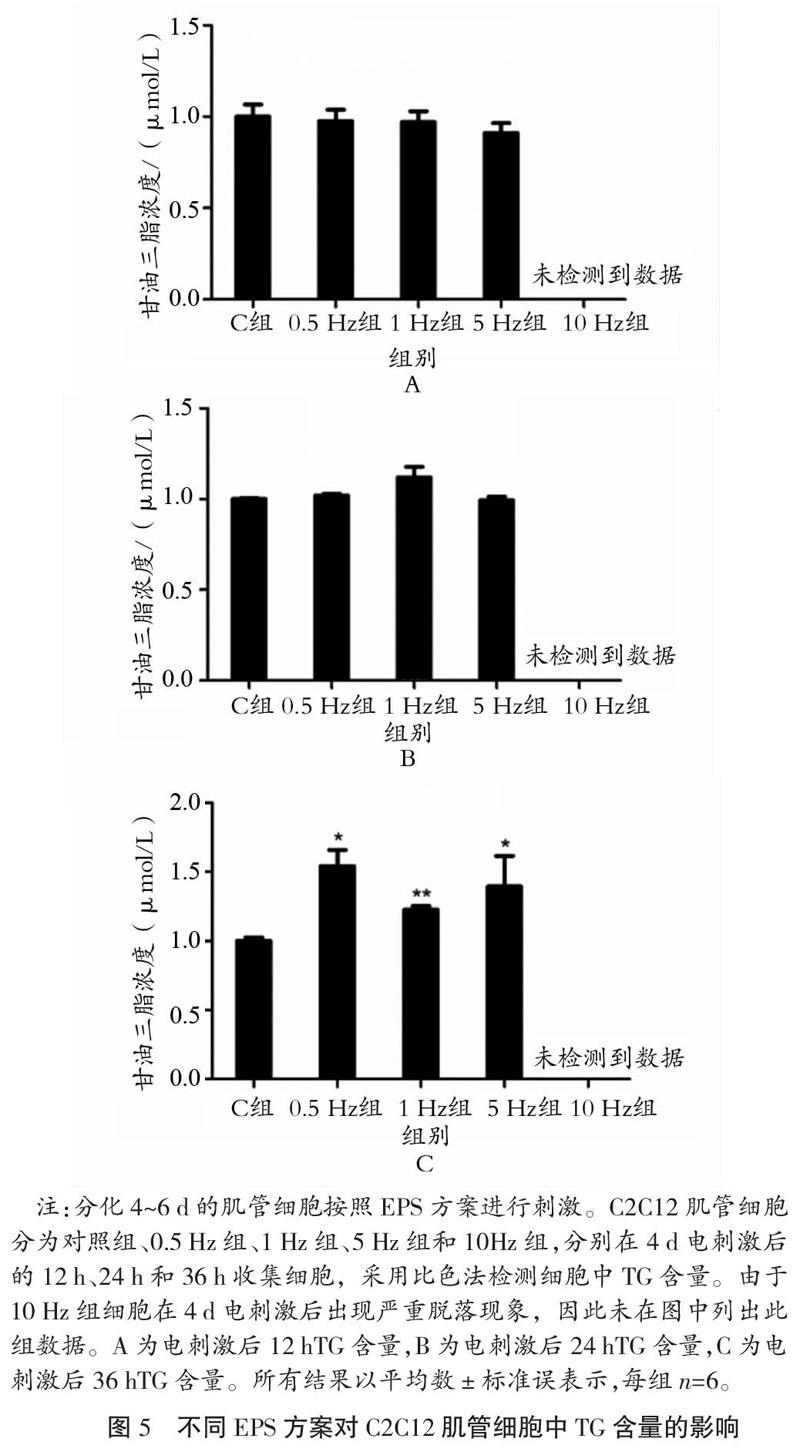
2.4 不同EPS方案对C2C12肌管细胞中TG含量的影响
如图5所示,与C组相比,4 d电刺激方案(电压30 V,频率0.5~5 Hz,单次时长2 ms)停止后12 h肌管细胞中TG含量均没有出现变化。4 d电刺激方案停止后24 h,与C组相比, 0.5 Hz组和1 Hz组肌管细胞中TG含量增加但未见显著性(P>0.05)。4 d电刺激方案停止后36 h,与C组相比,0.5 Hz组和5 Hz组细胞中TG含量显著增加(P<0.05),1 Hz组细胞中TG含量非常显著增加(P<0.01)。
3 讨论
本文的研究目的是采用EPS诱发C2C12肌管细胞收缩来模拟运动性骨骼肌收缩,从而研究不同EPS方案对C2C12肌管细胞TG、乳酸、ATP、MHC亚型和线粒体相关功能蛋白mRNA含量的影响,试图建立EPS诱发C2C12肌管细胞TG堆积和线粒体功能增加模型,为研究长期运动锻炼增加骨骼肌细胞中TG堆积并伴随线粒体功能增加的机制提供实验方案。本研究结果如下:1)一次性的电压30 V、频率0.5~10 Hz、单次时长2 ms共3 h的电刺激均能够非常显著增加C2C12细胞中乳酸含量并降低ATP含量;2)电刺激方案(电压14 V、频率1 Hz、单次时长2 ms和每天2 h共2 d,紧接着,电压30 V、频率0.5 Hz或1 Hz或5 Hz、单次时长2 ms和每天3 h共2 d,EPS停止后36 h)均能够显著诱发骨骼肌细胞中TG堆积和线粒体功能蛋白mRNA含量增加,在细胞水平上为进一步建立长期运动锻炼诱发骨骼肌细胞TG堆积和线粒体功能增强模型打下良好基础。
3.1 不同EPS方案诱发的C2C12肌管细胞收缩
规律性体力活动对健康具有促进作用,比如预防和治疗各种慢性疾病(Ⅱ型糖尿病、心血管疾病、抑郁症等)[15]。研究还显示运动过程中收缩的骨骼肌通过分泌肌因子参与调节运动使健康受益的过程[16]。近年来,在关于骨骼肌细胞运动适应性分子机制的研究中,EPS刺激骨骼肌细胞已经作为鉴定新颖肌肉因子的主要工具。尤其是近来发现的人类骨骼肌细胞的EPS方案,其对于理解规律性运动和健康促进之间分子机制具有重要作用[17]。体外细胞模型可能适用于研究收缩细胞(尤其是不同运动方案诱发细胞收缩)的信号通路和代谢适应性变化过程[18]。使用模拟骨骼肌收缩的体外模型,在理解骨骼肌代谢调控作用与其他器官之间复杂和多方向的“crosstalk”方面已经起到实质性进展[19]。研究表明,在细胞培养水平上,给予细胞EPS处理有利于骨骼肌细胞分化成肌管细胞,这些研究显示EPS能够激活骨骼肌细胞并且可以作为一种神经电信号的替代方法[11-12]。早期研究发现,不同电刺激方案诱导MHC亚型中快肌和慢肌的特异性改变,因此,确认EPS是研究体外骨骼肌收缩活性的一种有效模型[20]。C2C12细胞和主要大鼠细胞株对EPS是敏感的,它们主要被用于研究骨骼肌收缩-诱导代谢改变的过程。第1个报道EPS作为一种体外运动模型应用到骨骼肌细胞研究中的是C2C12细胞[21]。在此研究中,Nedachi等 [21]发现EPS诱导肌细胞出现明显收缩,同时伴随AMPK和MAPK激活、葡萄糖摄入量和胰岛素敏感性增加。本研究首先要确定不同EPS方案是否能够诱发C2C12肌管细胞收缩,采用EPS后肌管细胞中乳酸含量、ATP含量及MHC亚型mRNA含量变化作为判定肌管细胞收缩的指标。Nikolic等[13]采用一次性高频率EPS方案(30 V,100 Hz,间歇5 s,脉冲持续时间200 ms共5~60 min)能够显著增加细胞内乳酸含量、增加葡萄糖摄入量、降低ATP和PCr含量。本研究采用EPS方案为:分化4~6 d的肌管细胞,先进行2 d EPS(电压14 V,单次刺激时长2 ms,频率1 Hz,电刺激总时间2 h/d),其目的为利用此EPS方案来加速C2C12肌管与肌小节的生成[12];第3 天和第4 天分别进行一次高电压不同频率刺激(电压30 V,单次刺激时长2 ms,频率分别为0.5 Hz、1 Hz、5 Hz和10 Hz,EPS总时间3 h)。本研究结果发现:一次性EPS方案(电压30 V、频率0.5和10 Hz、脉冲2 ms 共3 h)也能够显著增加刺激后即刻细胞内乳酸含量,同时降低细胞内ATP含量。另一个EPS方案(电压11.5 V,频率1 Hz,脉冲2 ms共2~24 h)也发现低频刺激能够导致细胞骨架的重组、肌节结构重新合成和肌管明显收缩[22]。这些结果说明本研究采用的EPS方案能够促进肌管细胞收缩并消耗ATP作为能源供应。同时本研究也成功建立C2C12肌管细胞收缩后乳酸堆积模型,而且发现电压30 V是诱导C2C12肌管细胞乳酸堆积的一个关键参数。另外,MHC亚型基因表达情况常被用于区分不同骨骼肌纤维类型[23]。大量的研究表明,EPS能够导致MHC亚型基因表达的改变。长时间低频EPS促进大鼠肌管细胞分化和增加慢肌纤维类型标志物MyHCⅠ表达[24]。同时,在持续2周EPS过程中,肌纤维由快肌类型逐渐过渡到慢肌类型[25]。也有研究发现,1周EPS方案(电压7.5 V,频率3 Hz,脉冲10 ms,每次10 min)和1周EPS方法(电压7.5 V、频率3 Hz,脉冲10 ms,每次30 min)均使快肌纤维表达量降低而慢肌纤维则无改变[26]。另外,研究报道:电压30 V、频率1 Hz、脉冲时长2 ms共24 h或者48 h 的EPS能够显著增加MyHCⅠ(慢缩氧化型肌纤维标志物)含量[13]。这些结果表明,长时间低频EPS促进肌管细胞慢肌纤维增加。本研究数据也显示:电压30 V、频率0.5 Hz或1 Hz、单次时长2 ms共3 h的EPS能够显著增加MyHCⅠ和MyHCⅡb型mRNA含量,这说明这一EPS方案能够显著增加骨骼肌氧化型和快速收缩型肌纤维含量,这可能与本研究所采用的电刺激方案中电压较大且持续时间相对较短有关。而电压30 V、频率10 Hz、单次时长2 ms共3 h的EPS相对C2C12肌管细胞来说强度较大,主要以增加快缩型MyHCⅡb亚型mRNA含量为主。另外,IL-6被认为是机体骨骼肌收缩的另一个标志细胞因子[27-28]。Nedachi等[21]研究发现24 hEPS(频率1 Hz)诱导C2C12细胞中许多来源于收缩骨骼肌细胞肌因子的表达,例如IL-6和CXCL等。本研究也发现:EPS方案能够使C2C12肌管细胞中IL-6 mRNA含量增加,而且呈现频率依赖性。综合这些数据,认为本研究采用的EPS方案(电压14 V、频率1 Hz、单次时长2 ms和每天2 h共2 d,紧接着,电压30V、频率0.5 Hz~10 Hz、单次时长2 ms和每天3 h共2 d)能够诱发C2C12肌管细胞收缩。
3.2 长时间不同频率EPS诱发C2C12肌管细胞代谢指标和基因表达变化
大量研究表明,长时间耐力训练能够增加骨骼肌细胞中TG含量和線粒体功能。例如耐力性项目运动员往往伴随着骨骼肌细胞中脂滴含量增加和线粒体功能增强[7-8]。同时,长时间锻炼中老年人其骨骼肌细胞中脂质含量显著增加伴随骨骼肌氧化能力增强[1,7]。另外,有研究发现2周的训练(有氧耐力和间隙功率自行车训练)就能够使无训练男性(年龄(23.3±3.2)岁)股外侧肌中的IMCL显著增加(42±14)%[3],但是其机制目前尚不清楚。建立一种能够模拟长时间运动诱发骨骼肌收缩的体外细胞模型从而研究其机制显得尤为重要。本研究探究了长时间不同频率EPS对C2C12肌管细胞代谢指标和基因表达的影响,其将为研究长时间耐力训练增加骨骼肌细胞TG含量的机制提供帮助。本研究发现:电压30 V、单次刺激时长2 ms、频率分别为0.5 Hz或1 Hz或5 Hz、停止电刺激后36 h的方案能够显著增加C2C12肌管细胞TG含量。本研究结果显示:EPS能够显著增加C2C12肌管细胞分泌乳酸含量、降低ATP含量和改变MHC亚型mRNA含量等,本研究认为此EPS方案(电压14 V、频率1 Hz、单次时长2 ms和每天2 h共2 d,紧接着,电压30 V、频率0.5 Hz或1 Hz或5 Hz、单次时长2 ms和每天3 h共2 d,EPS停止后36 h)是可用于研究长期运动训练诱导骨骼肌TG堆积的成功建立的细胞模型。
另外,耐力训练诱发线粒体功能和氧化能力增强[29-30]。PGC-1α作为一个转录调节因子,是线粒体功能和合成增加的关键调节蛋白[31]。作为肌肉训练的标志蛋白,PGC-1α常被认为是运动诱发骨骼肌出现代谢改变的细胞内因子[13,32]。PGC-1α这一线粒体蛋白是作为转录因子发挥作用,其mRNA含量代表着线粒体功能和合成能力[31]。研究发现长时间EPS能使PGC-1αmRNA含量增加2.4~2.9倍[14]。Cyt C是线粒体氧化呼吸电子传递链成分,其含量高低代表着细胞线粒体氧化还原功能[33]。本研究采用的EPS方案均能够显著增加C2C12肌管细胞中PGC-1α和Cyt C 的mRNA含量。另外,被认为是机体骨骼肌收缩的标志肌源性细胞因子之一的IL-6[27-28],EPS后细胞中含量也出现增多。Nikolic等[13]采用的48 hEPS能够增加线粒体含量和柠檬酸合成酶活性。Lambernd等[22]发现24 hEPS方案可增加蛋白复合物Ⅱ、Ⅳ和Ⅴ含量。这些说明EPS使C2C12肌管细胞线粒体出现适应性变化,线粒体功能增强。EPS诱导PGC-1α增加与骨骼肌氧化型肌纤维类型增多的结果是一致的,因此PGC-1α作为转录调节因子,它的升高伴随着骨骼肌类型由糖酵解、快缩型向氧化型肌纤维类型转化[34-35]。EPS诱发C2C12肌管细胞Cyt C和PGC-1α mRNA含量的增加,这可能是由于EPS促使肌管细胞收缩分泌乳酸引起的。因为研究发现:6 h乳酸处理L6细胞后其Cyt C和PGC-1α的mRNA含量和蛋白含量显著增加[36]。综上所述,电刺激方案(电压14 V、频率1 Hz、单次时长2 ms和每天2 h共2 d,紧接着,电压30 V、频率0.5 Hz或1 Hz或5 Hz、单次时长2 ms和每天3 h共2 d,EPS停止后36 h)均能够显著诱发骨骼肌细胞中线粒体功能蛋白mRNA含量增加,同时“乳酸”这一代谢物将为长期运动训练增加骨骼肌线粒体功能增加提供新思路。
4 结论
EPS方案(电压14 V、频率1 Hz、单次时长2 ms和每天2 h共2 d,紧接着,电压30 V、频率0.5 Hz或1 Hz或5 Hz、单次时长2 ms和每天3 h共2 d,EPS停止后36 h)能够诱发C2C12肌管细胞TG堆积和线粒体功能增强,该方案是诱发C2C12肌管细胞TG堆积和线粒体功能增强的成功建立的细胞模型。
参考文献:
[1] PRUCHNIC R, KATSIARAS A, HE J, et al. Exercise training increases intramyocellular lipid and oxidative capacity in older adults[J]. Am J Physiol Endocrinol Metab, 2004, 287(5): 857.
[2] SHAW C S, SHEPHERD S O, WAGENMAKERS A Journal, et al. Prolonged exercise training increases intramuscular lipid content and perilipin 2 expression in type I muscle fibers of patients with type 2 diabetes[J]. Am J Physiol Endocrinol Metab, 2012, 303(9): 1158.
[3] SCHRAUWEN-HINDERLING V B, SCHRAUWEN P, HESSELINK M K, et al. The increase in intramyocellular lipid content is a very early response to training[J]. J Clin Endocrinol Metab,2003, 88(4): 1610.
[4] HAVEL R J. Influence of intensity and duration of exercise on supply and use of fuels[M]. New York: Plenum, 1971: 315.
[5] HURLEY B F, NEMETH P M, MARTIN W R, et al. Muscle triglyceride utilization during exercise: effect of training[J]. J Appl Physiol (1985), 1986, 60(2): 562.
[6] HOPPELER H, HOWALD H, CONLEY K, et al. Endurance training in humans: aerobic capacity and structure of skeletal muscle[J]. J Appl Physiol (1985), 1985, 59(2): 320.
[7] GOODPASTER B H, HE J, WATKINS S, et al. Skeletal muscle lipid content and insulin resistance: evidence for a paradox in endurance-trained athletes[J]. J Clin Endocrinol Metab, 2001, 86(12): 5755.
[8] AMATI F, DUBE J J, ALVAREZ-CARNERO E, et al. Skeletal muscle triglycerides, diacylglycerols, and ceramides in insulin resistance: another paradox in endurance-trained athletes [J]. Diabetes, 2011, 60(10): 2588.
[9] DUBE J J, BROSKEY N T, DESPINES A A, et al. Muscle characteristics and substrate energetics in lifelong endurance athletes[J]. Med Sci Sports Exerc, 2016, 48(3): 472.
[10] NEDACHI T, FUJITA H, KANZAKI M. Contractile C2C12 myotube model for studying exercise-inducible responses in skeletal muscle[J]. Am J Physiol Endocrinol Metab, 2008, 295(5): 1191.
[11] THELEN M H, SIMONIDES W S, VAN HARDEVELD C. Electrical stimulation of C2C12 myotubes induces contractions and represses thyroid-hormone-dependent transcription of the fast-type sarcoplasmic-reticulum Ca2+-ATPase gene[J]. Biochem J, 1997, 321 (3): 845.
[12] FUJITA H, NEDACHI T, KANZAKI M. Accelerated de novo sarcomere assembly by electric pulse stimulation in C2C12 myotubes[J]. Exp Cell Res, 2007, 313(9): 1853.
[13] NIKOLIC N, BAKKE S S, KASE E T, et al. Electrical pulse stimulation of cultured human skeletal muscle cells as an in vitro model of exercise[J]. PLoS One, 2012, 7(3): 33203.
[14] BURCH N, ARNOLD A S, ITEM F, et al. Electric pulse stimulation of cultured murine muscle cells reproduces gene expression changes of trained mouse muscle[J]. PLoS One, 2010, 5(6): 10970.
[15] PEDERSEN B K, SALTIN B. Exercise as medicine - evidence for prescribing exercise as therapy in 26 different chronic diseases[J]. Scand J Med Sci Sports, 2015, 25(S3): 1.
[16] FEBBRAIO M A, PEDERSEN B K. Contraction-induced myokine production and release: is skeletal muscle an endocrine organ [J]. Exerc Sport Sci Rev, 2005, 33(3): 114.
[17] GORGENS S W, ECKARDT K, JENSEN J, et al. Exercise and regulation of adipokine and myokine production[J]. Prog Mol Biol Transl Sci, 2015(135): 313.
[18] NIKOLIC N, GORGENS S W, THORESEN G H, et al. Electrical pulse stimulation of cultured skeletal muscle cells as a model for in vitro exercise - possibilities and limitations[J]. Acta Physiol (Oxf), 2017, 220(3): 310.
[19] ECKARDT K, GORGENS S W, RASCHKE S, et al. Myokines in insulin resistance and type 2 diabetes[J]. Diabetologia, 2014, 57(6): 1087.
[20] NAUMANN K, PETTE D. Effects of chronic stimulation with different impulse patterns on the expression of myosin isoforms in rat myotube cultures[J]. Differentiation, 1994, 55(3): 203.
[21] NEDACHI T, FUJITA H, KANZAKI M. Contractile C2C12 myotube model for studying exercise-inducible responses in skeletal muscle[J]. Am J Physiol Endocrinol Metab, 2008, 295(5): 1191.
[22] LAMBERND S, TAUBE A, SCHOBER A, et al. Contractile activity of human skeletal muscle cells prevents insulin resistance by inhibiting pro-inflammatory signalling pathways[J]. Diabetologia, 2012, 55(4): 1128.
[23] SPANGENBURG E E, BOOTH F W. Molecular regulation of individual skeletal muscle fibre types[J]. Acta Physiol Scand, 2003, 178(4): 413.
[24] WEHRLE U, DUSTERHOFT S, PETTE D. Effects of chronic electrical stimulation on myosin heavy chain expression in satellite cell cultures derived from rat muscles of different fiber-type composition[J]. Differentiation, 1994, 58(1): 37.
[25] KUBIS H P, SCHEIBE R J, MEISSNER J D, et al. Fast-to-slow transformation and nuclear import/export kinetics of the transcription factor NFATc1 during electrostimulation of rabbit muscle cells in culture[J]. J Physiol, 2002, 541(3): 835.
[26] ZEBEDIN E, SANDTNER W, GALLER S, et al. Fiber type conversion alters inactivation of voltage-dependent sodium currents in murine C2C12 skeletal muscle cells[J]. Am J Physiol Cell Physiol, 2004, 287(2): 270.
[27] PEDERSEN B K, STEENSBERG A, FISCHER C, et al. The metabolic role of IL-6 produced during exercise: is IL-6 an exercise factor [J]. Proc Nutr Soc, 2004, 63(2): 263.
[28] PEDERSEN B K, STEENSBERG A, FISCHER C, et al. Searching for the exercise factor: is IL-6 a candidate [J]. J Muscle Res Cell Motil, 2003, 24(2-3): 113.
[29] COFFEY V G, HAWLEY J A. The molecular bases of training adaptation[J]. Sports Med, 2007, 37(9): 737.
[30] HOOD D A, IRRCHER I, LJUBICIC V, et al. Coordination of metabolic plasticity in skeletal muscle[J]. J Exp Biol, 2006, 209(12): 2265.
[31] GLEYZER N, VERCAUTEREN K, SCARPULLA R C. Control of mitochondrial transcription specificity factors (TFB1M and TFB2M) by nuclear respiratory factors (NRF-1 and NRF-2) and PGC-1 family coactivators[J]. Mol Cell Biol, 2005, 25(4): 1354.
[32] HANDSCHIN C. The biology of PGC-1alpha and its therapeutic potential[J]. Trends Pharmacol Sci, 2009, 30(6): 322.
[33] JOSEPHS T M, LIPTAK M D, HUGHES G, et al. Conformational change and human cytochrome c function: mutation of residue 41 modulates caspase activation and destabilizes Met-80 coordination[J]. J Biol Inorg Chem, 2013, 18(3): 289.
[34] LIN J, WU H, TARR P T, et al. Transcriptional co-activator PGC-1 alpha drives the formation of slow-twitch muscle fibres[J]. Nature, 2002, 418(6899): 797.
[35] HANDSCHIN C, CHIN S, LI P, et al. Skeletal muscle fiber-type switching, exercise intolerance, and myopathy in PGC-1alpha muscle-specific knock-out animals[J]. J Biol Chem, 2007, 282(41): 30014.
[36] HASHIMOTO T, HUSSIEN R, OOMMEN S, et al. Lactate sensitive transcription factor network in L6 cells: activation of MCT1 and mitochondrial biogenesis[J]. The FASEB Journal, 2007, 21(10): 2602.
