中国南瓜半短蔓种质‘CmSd1’的鉴定和CmSd-1t基因定位
2019-08-03罗文龙王青李俊星钟玉娟郭巨先李桂花陈木溪黄河勋
罗文龙 王青 李俊星 钟玉娟 郭巨先 李桂花 陈木溪 黄河勋

摘 要:植株适度矮生可使株型更紧凑、空间利用率更高,有利于密植栽培、机械化采收和提高单位面积产量。以从多亲本复交后代选育的中国南瓜半短蔓种质‘CmSd1为材料,从表型鉴定、外源赤霉素响应、遗传分析和基因定位等方面进行研究。表型观察发现,‘CmSd1的半短蔓表型主要是由于主蔓中下部节间变短所致;‘CmSd1对外源赤霉素表现为敏感且敏感度比长蔓对照更高,推测其半短蔓性状与赤霉素代谢途径密切相关;遗传分析表明,‘CmSd1的半短蔓性状受1对显性基因控制,暂命名为CmSd-1(t);利用F2极端表型混合池进行重测序和BSA分析,将CmSd-1(t)初步定位到中国南瓜第3染色体末端Cmo_Chr03:9.985-10.828 Mb之间。CmSd-1(t)控制的显性半短蔓材料,具有良好的农艺性状,在中国南瓜矮化育种中有较大的应用前景。
关键词:中国南瓜;半短蔓;赤霉素;基因定位
Abstract: The moderate dwarf of plants makes them more compact and higher space utilization, facilitating to grow with higher density, mechanized harvesting and hence producing a higher yield per unit area. Herein, we reported the phenotypic and genetic characterization of a semi-dwarf germplasm designated as ‘CmSd1 (Cucurbita moschata Semi-dwarf 1), which was isolated from crossing progeny of multiple pumpkin varieties (C. moschata). We found that the semi-dwarf phenotype of ‘CmSd1 was mainly resulted from shortened of internodes in the lower part of the vine. The ‘CmSd1 was more sensitive to exogenous gibberellin treatment than the normal long-vine variety, indicating that its semi-dwarf was associated with gibberellin metabolism. Genetic analysis revealed that semi-dwarf in the ‘CmSd1 was controlled by a dominant major-effect gene temporarily named as CmSd-1(t). We then mapped the CmSd-1(t) to the region Cmo_Chr03:9.985-10.828 Mb at the end of chromosome 3 of the C. moschata genome, via bulked-segregant sequencing of F2 pools of extreme individuals. We concluded that the CmSd-1(t), controlling semi-dwarf phenotype without significant negative effects on agronomic important traits, would have a great utilization foreground in developing moderated-dwarf pumpkin varieties.
Key words: Pumpkin (Cucurbita moschata); Semi-dwarf; Gibberellins; Genetic mapping
半矮生資源的挖掘和利用,推动了第一次“绿色革命”,在全球范围内促成了粮食的大增产[1-2]。植株适度矮生可使株型更紧凑、空间利用率更高,有利于密植栽培、机械化采收和提高单位面积产量,也成为园艺作物育种的重要目标。因此,挖掘创制矮生、半矮生材料,日渐受到育种家的重视。
南瓜为葫芦科(Cucurbitaceae)南瓜属(Cucurbita)的一年生蔓生藤本植物,是重要的世界性园艺作物。据统计,2013年我国南瓜种植面积为110万hm2,总产量超过3 000万t,是世界南瓜生产和消费第1大国[3]。南瓜遗传多样性丰富,其栽培种包括中国南瓜(C. moschata)、印度南瓜(C. maxima)、美洲南瓜(C. pepo,即西葫芦)、灰籽南瓜(C. mixta)和黑籽南瓜(C. ficifolia)。按主蔓长度,南瓜可以分为蔓生(长蔓)、半蔓生(半短蔓)和矮生(短蔓丛生)等类型;在生产上,美洲南瓜主要为矮生、半蔓生类型品种,印度南瓜主要为蔓生、半蔓生类型品种,而中国南瓜大多为长蔓品种,短蔓品种还很少[3-4]。
由‘蜜本南瓜及其衍生出的中国南瓜品种,是典型的大果型南瓜,曾占我国南瓜种植面积的80%以上,在很大程度上推动了我国南瓜品种的良种化。蜜本类型南瓜的选育和推广,在我国农业种植结构调整中起到重要作用。然而,近十多年来,以此类型为代表的中国南瓜品种同质化严重,产量、品质提高缓慢,也难以满足机械化生产的需求。针对这些问题,我们将中国南瓜矮化育种作为一个重点育种方向,加大短蔓、半短蔓材料的挖掘和创新创制。笔者以一个从多亲本复交后代选育的中国南瓜半短蔓材料为对象,从表型鉴定、喷施外源赤霉素处理、遗传分析和基因定位等方面进行分析,以期为后续开展分子标记辅助育种和基因功能研究奠定基础。
1 材料与方法
1.1 材料
供试材料为中国南瓜半短蔓自交系,暂命名为‘CmSd1(C. moschata Semi-dwarf 1),由广东省农业科学院蔬菜研究所自主选育和保存。‘CmSd1是由德国奶油南瓜‘Sperli Early Butternut F1(C. moschata)自交后代,与‘一串铃1号(C. moschata)自交后代、蜜本南瓜(C. moschata)自交后代复交,经16代连续自交定向筛选,于2016年定型后自交纯化而获得。经过2017—2018年共4代鉴定,‘CmSd1均表现为半短蔓,植株健壮,椭圆形瓜,单瓜质量在2.0 kg以上,主要农艺性状良好。
利用‘CmSd1与3个有代表性的大果型中国南瓜长蔓自交系(‘D205‘D214‘D220,均由广东省农业科学院蔬菜研究所选育)杂交,F1都表现为半短蔓。以长蔓自交系‘D205作为对照进行表型鉴定,并利用‘CmSd1与‘D205杂交产生的F1、F2群体开展遗传分析。
1.2 方法
1.2.1 田间试验 田间试验于2018年晚季(8—11月)在广东省农业科学院白云试验基地(广州)完成。自交系及杂交后代催芽育苗,在2片真叶时移栽。待主蔓伸长后,及时进行植株吊蔓、整枝。
1.2.2 外源赤霉素处理 试验所用药品赤霉素(GA3)购自生工生物工程(上海)股份有限公司。GA3处理设4个质量浓度处理,分别为0、100、300和600 mg·L-1;每个处理设3个重复,每重复处理9株。待幼苗处于3叶期,于傍晚将不同浓度的GA3喷施在供试植株叶片上,至有液滴形成。此后,每隔3 d喷施1次,连续喷施5次。在第5次喷施GA3后第3天,每个处理分别从每个重复取3株共9株,用直尺测量株高,然后取其平均值。
1.2.3 表型性状调查与统计分析 在整个生长期间,观察南瓜材料主蔓伸长情况。在移栽20 d时,对主蔓是否伸长进行调查和标记;到生长中后期(移栽60 d),调查的第1节茎粗、前15节蔓长和前30节蔓长。
1.2.4 极端个体混合池测序和BSA分析 在F2群体选取20个长蔓个体,取等量叶片混合提取DNA,作为长蔓极端混合池;选取20个半短蔓个体,取等量叶片混合提取DNA,作为半短蔓极端混合池。将极端混合池的高质量基因组DNA,依次进行DNA片段化、片段纯化、末端修复、3′端加A、连接测序接头、片段大小选择和PCR扩增构建测序文库(双末端150 bp),进而通过Illumina Hiseq平台进行测序。2个混合池测序产生的数据经过过滤,然后利用BWA[5]比对到中国南瓜基因组(C. moschata cv. Rifu, 下载地址ftp://cucurbitgenomics.org/pub/cucurbit/genome/Cucurbita_moschata/v1/),再通过SAMtools[6]和Varscan 2[7]检测SNP。利用获得的SNP计算平方欧氏距离(squared Euclidean distance, ED2)进行BSA分析[8],以确定候选基因在染色体上的区域。
2 结果与分析
2.1 中国南瓜半短蔓材料‘CmSd1的表型特征
对半短蔓材料‘CmSd1和长蔓自交系‘D205的表型进行持续观察。移栽20 d(生长前期),‘D205有8~10片真叶,节间在6~8节之后明显拉长,使其主蔓迅速伸长;而‘CmSd1生长相对较慢,只有6~8片真叶,且全部节间都很短,主蔓没有伸长(图1-A)。‘CmSd1在13~15节后,节间才逐渐伸长。到生长中期(移栽40 d),可以观察到‘CmSd1主蔓基部节间明显比‘D205要短(图1-B~C)。生长中后期(移栽60 d),‘CmSd1与‘D205主蔓基部节间的长度差异更为明显,但2者中上部节间(25~30节)长度差异不明显(图1-D)。结合前15节和前30节蔓长(表1),推测‘CmSd1基部节间相对更短是导致其主蔓更短的主要原因。此外,‘D205瓜为长条形(图1-E),‘CmSd1瓜为椭圆形(图1-F),2者瓜都较大,其F1瓜型呈长棒形。半短蔓材料‘CmSd1综合农艺性状优异,在大果型南瓜育种中的潜在应用价值较大。
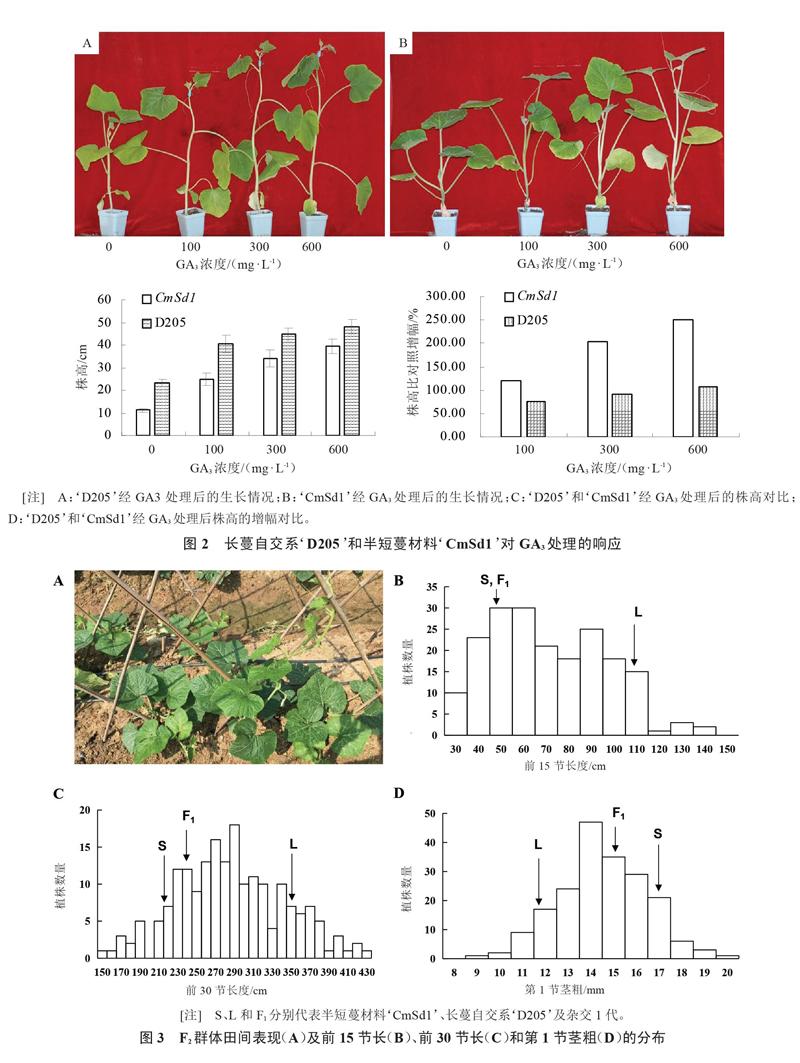
2.2 喷施外源赤霉素能促进中国南瓜半短蔓材料的生长和伸长
植物激素能够对植物的生长和发育进行调控,其中赤霉素对株高(蔓长)的影响最大。对长蔓自交系‘D205和半短蔓材料‘CmSd1喷施不同浓度的GA3进行处理,以分析其对外源赤霉素的敏感性。结果表明,在相同处理下2个材料的株高差异都达到极显著水平;相对于对照(喷施清水),喷施GA3的2个材料节间都明显伸长,使得株高均极显著高于对照,且触须提早出现(图2-A~C)。但是,半短蔓材料‘CmSd1株高增幅要更大,说明该材料对喷施GA3更加敏感(图2-D)。可见,‘CmSd1对外源赤霉素敏感,且敏感度比长蔓材料更高,推测其半短蔓性状很可能与赤霉素合成或信号传导途径密切相关。
2.3 中国南瓜半短蔓材料‘CmSd1的遗传分析
利用‘CmSd1与‘D205杂交产生的F1、F2群体开展遗传分析,其中F2群体在生长前期(移栽20 d后)的表现如图3-A所示。移栽20 d后,‘D205主蔓已明显伸长,而F1表现为全部节间很短、主蔓没有伸长,与‘CmSd1完全一致。以‘D205和‘CmSd1为对照,对F2群体进行调查,发现主蔓未伸长和已伸长的单株数量分别为144株和51,符合3∶1的分离比(χ2检验p=0.71>0.05)。由此可知,在生长前期,‘CmSd1主蔓未伸长相对‘D205主蔓已伸长为显性,是受1对基因调控的质量性状。
‘CmSd1和F1主蔓在13~15节后逐渐伸长,在生长中后期(移栽60 d)对前15节蔓长、前30节蔓长和第1节茎粗进行调查。F1前15节蔓长与‘CmSd1基本一致,明显短于‘D205;其前30节蔓长要比‘CmSd1更长,但仍然明显比‘D205短(表1和图3-B)。F2群体前15节蔓长为较明显的双峰分布(图3-A),而前30节蔓长则表现为偏向于‘CmSd1的偏态分布(图3-B)。可见,由于‘CmSd1与‘D205的节间长度差异主要在下部节位,而上部节间长差异不明显,因此F2群体蔓长在中后期表现为主效QTL控制的数量性状遗傳。此外,F2群体第1节茎粗分布符合正态分布(图3-D),表明此性状由微效基因控制,与蔓长没有相关性。综合分析,我们认为‘CmSd1的半短蔓表型是由中下部节间变短所致,受1对显性基因控制,暂命名为CmSd-1(t)。
2.4 中國南瓜半短蔓基因CmSd-1(t)的快速定位
为了对半短蔓基因CmSd-1(t)进行定位,在F2群体取移栽20 d主蔓已伸长(隐性)和未伸长(显性)的个体各20个,分别构建极端表型混合池进行基因组重测序和BSA分析。其中,隐性混合池取样个体前15节平均长105.06 cm,前30节平均长338.94 cm;显性混合池取样个体前15节平均长54.12 cm,前30节平均长220.77 cm。利用2个池的SNP频率计算平方欧式距离(ED2),绘制散点图和拟合曲线进行连锁分析,发现在第3染色体末端的ED2显著高于其他区域(图4)。进一步比较分析该区域SNP频率在2个池的差异,将半短蔓基因CmSd-1(t)初步定位在中国南瓜第3染色体末端Cmo_Chr03:9.985-10.828 Mb。
3 讨 论
植株适度矮生可使株型更紧凑、空间利用率更高,有利于密植栽培、机械化采收和提高单位面积产量。水稻中,除了sd-1之外,虽然已鉴定了超过70个基因可以导致矮杆、半矮秆性状,但这些基因除了引起矮化还会影响其他农艺性状,使其难以应用于培育矮杆抗倒伏品种[9]。因此,挖掘和创制具有良好农艺性状的矮生资源尤显重要。
目前,南瓜属作物矮生性状的研究还较少,而且主要集中在美洲南瓜(西葫芦)和印度南瓜。已报道的中国南瓜矮生(丛生)资源仅有Bu和cga,2者均表现为主蔓极短,植株丛生[10-11]。由于中国南瓜(尤其是大果型中国南瓜)在生产上以爬地栽培、采收老瓜为主,矮化丛生较为不利,因而这些矮生材料的直接育种利用价值有限。本研究对自主创制的一个中国南瓜半短蔓材料‘CmSd1进行了鉴定,发现其半短蔓表型主要是由中下部节间相对更短导致(图1),到生长中后期蔓长相对于长蔓自交系短约1/3(表1)。相比其他已知的中国南瓜矮生材料,‘CmSd1的半短蔓(半矮生)表型明显不同,更适于爬地栽培,单瓜大(2.0 kg以上),在育种中应用价值更高。
研究者们利用大量突变体逐渐揭示了植物激素调控株高的复杂网络,其中赤霉素代谢和信号传导途径起到关键作用[12]。大多数植物矮化是由于突变体内赤霉素和油菜素内酯合成或信号转导受到影响,少数与生长素、独脚金内酯有关[13]。水稻“绿色革命”基因sd-1就是编码赤霉素生物合成途径关键酶GA20ox的基因GA20ox-2发生突变,导致节间缩短而不影响穗部性状,使植株矮化、株型更紧凑和更耐肥,最终推动单位面积产量大幅提高[2]。本研究中鉴定的半短蔓材料‘CmSd1对赤霉素敏感,且敏感度比长蔓自交系更高。由此推测,‘CmSd1的半短蔓表型可能与由于赤霉素代谢或信号传导途径密切相关。此外,激素代谢和信号通路存在广泛的互作,某种激素含量的改变往往会引起其他激素的变化,从而导致植株矮化,同时影响到生长发育和其他性状。‘CmSd1 在前期的生长速度比长蔓材料更慢,暗示其生长发育可能受到多种激素相互作用的影响。
通过对半短蔓材料‘CmSd1与长蔓自交系的杂交后代群体进行遗传分析,发现半短蔓表型受1个显性基因调控,暂命名为CmSd-1(t)。利用极端池进行BSA测序分析,我们将CmSd-1(t)初步定位于中国南瓜基因组第3染色体末端Cmo_Chr03:9.985~10.828 Mb之间(图4)。在南瓜属作物上,包括qCmB2(定位于印度南瓜第3连锁群)[14]、qCpDy1和qCpDm1(定位于美洲南瓜第20连锁群)[15]、dm1[16](印度南瓜)、Bu(中国南瓜,比较定位至黄瓜5 号染色)[17]等多个矮生性状相关的基因/QTL已得到定位。然而,由于南瓜属作物的基因组发展较晚,对不同栽培种的比较基因组研究还较少,笔者定位的CmSd-1(t)和上述基因/QTL是否是等位基因或同源基因还有待进一步深入分析。上述这些基因/QTL控制的矮生表型均为短蔓丛生,尤其是携带中国南瓜矮生基因Bu的突变体表现为对外源赤霉素低敏感。相比这些已知的基因/QTL,CmSd-1(t)调控显性半短蔓表型,对外源赤霉素高度敏感,由此推测,CmSd-1(t)调控蔓长的分子机制与已知南瓜矮生基因/QTL可能是不同的,有可能是新的南瓜矮生基因。
综上所述,CmSd-1(t)控制半短蔓表型且对其他重要农艺性状无不利影响,在中国南瓜育种中的潜在价值较大,有必要对其开展精细定位、分子克隆和功能研究,从而为更加科学合理地利用其开展南瓜矮化育种提供支撑。
参考文献
[1] SASAKI A,ASHIKARI M,UEGUCHI-TANAKA M,et al.Green revolution: a mutant gibberellin-synthesis gene in rice[J].Nature,2002,416(6882):701.
[2] HEDDEN P.The genes of the Green Revolution[J].Trends in Genetics,2003,19(1):5-9.
[3] 方智远.中国蔬菜育种学[M].北京: 中国农业出版社,2017.
[4] 雷逢进,王晓民,马理军,等.不同矮蔓性西葫芦产量与其构成因素的关系[J].中国瓜菜,2013,26(3):23-26.
[5] LI H,DURBIN R.Fast and accurate short read alignment with burrows-wheeler transform[J].Bioinformatics,2009,25(14):1754-1760.
[6] LI H,HANDSAKER B,WYSOKER A,et al.The sequence alignment/map format and SAMtools[J].Bioinformatics,2009,25(16):2078-2079.
[7] KOBOLDT D C,ZHANG Q,LARSON D E,et al.VarScan 2:somatic mutation and copy number alteration discovery in cancer by exome sequencing[J].Genome Research,2012,22(3):568-576.
[8] HILL JT,DEMAREST BL,BISGROVE BW,et al.MMAPPR:mutation mapping analysis pipeline for pooled RNA-seq[J].Genome Research,2013,23(4):687-697.
[9] LIU C,ZHENG S,GUI J,et al.Shortened basal internodes encodes a gibberellin 2-oxidase and contributes to lodging resistance in rice[J].Molecular Plant,2018,11(2):288-299.
[10] 李云龙,李海真,崔崇士,等.与南瓜矮生基因连锁的分子标记[J].农业生物技术学报,2007,15(2): 279-282.
[11] WU T,CAO J,QIN Z,et al.Identification of a novel ga-related bush mutant in pumpkin (Cucurbita moschata Duchesne)[J].Pakistan journal of botany,2015,47(4):1359-1366.
[12] WANG Y,ZHAO J,LU W,et al.Gibberellin in plant height control:old player,new story[J].Plant Cell Reports,2017,36(3):391-398.
[13] 白丽君,尹淑霞.植物矮化突变体的来源及矮化机理研究进展[J].生物技术通报,2014(6):34-39.
[14] ZHANG G,REN Y,SUN H,et al.A high-density genetic map for anchoring genome sequences and identifying QTLs associated with dwarf vine in pumpkin (Cucurbita maxima Duch.)[J].BMC Genomics,2015,16(1):1101.
[15] XIANG C,DUAN Y,LI H,et al.A high-density EST-SSR-based genetic map and QTL Analysis of dwarf trait in Cucurbita pepo L.[J].International Journal of Molecular Sciences,2018,19(10):3140.
[16] WANG R,HUANG H,LIN Y,et al.Genetic and gene expression analysis of dm1,a dwarf mutant from Cucurbita maxima Duch.ex Lam,based on the AFLP method[J].Canadian Journal of Plant Science,2014,94(2):293-302.
[17] 王深浩.中國南瓜矮生基因Bu的比较定位及其矮生性状的生理研究[D].北京:中国农业科学院,2010.
