应力刺激下TGFβ/Smad信号通路介导成骨细胞功能变化的研究
2019-07-24吕春晓李贵凤陶贵渝陈建伟
程 烨 吕春晓 李贵凤 陶贵渝 陈建伟 李 煌
成骨细胞是骨形成的效应细胞,在骨改建的机制中处于“中心调控地位”,被认为是力学信号转导过程中对应力应变信号进行感知的主要力学敏感性细胞。许多研究显示,体外培养的成骨细胞在机械应力刺激下会发生基因表达的改变。应力刺激在1小时内就能增加成骨细胞环氧合酶2(COX-2)和c-fos mRNA 的表达水平[1],24 小时内可提升其细胞外基质蛋白骨钙素(OCN)及骨桥蛋白的表达[2]。
转化生长因子β(TGFβ)是一类强力的多功能细胞因子,其主要的生理功能可能包括调控细胞的生长,刺激基质的分泌及免疫抑制。在牵张成骨过程中,TGFβ 是重要的参与者。在多项动物颌骨牵张实验中发现,TGFβ1 表达在延迟期、牵张期和早期固定期均有增加[3]。本课题组的前期研究通过生长期山羊骨缝牵张的实验发现,骨缝边缘出现大量成纤维细胞和成骨细胞,BMP 和TGFβ 在这些增殖活跃的细胞中呈高水平表达[4]。体外研究发现,由破骨细胞释放激活的TGFβ 可以通过活化成骨前体细胞并刺激其增殖而增加成骨细胞的数量来刺激骨的形成[5,6]。
成骨细胞中Ⅰ型胶原(COL-I)的表达始于增殖期,基质合成期达到高峰;碱性磷酸酶(ALP)是成骨细胞早期分化标志物,基质合成期开始表达,钙化期达到高峰;OCN 是成骨细胞成熟标志,从钙化早期开始表达,钙结节成熟后达高峰。功能性Runx2 水平决定了骨骼成熟程度及转换率。Runx2 存在于增殖活跃的成骨细胞细胞前体中,并在细胞开始分化时表达最高,Runx2 通过使细胞退出细胞周期和激活骨细胞表型基因促进成骨细胞分化成熟[7,8]。因此,OCN、COL-I、ALP 和Runx2 是判定成骨细胞功能活性的重要指标。
大鼠腭中缝牵张实验显示,骨改建活跃区域细胞TGFβ/Smad 信号分子表达水平发生变化[9]。然而,在此过程中,各类细胞的功能活动改变及机制仍未得到全面详细的描述。为了更深入的了解这一系列细胞分子事件的机械感应机制,我们对机械张应力作用下成骨细胞增殖活性,TGFβ/Smad、OCN、COL-I、ALP 和Runx2 等表达变化规律及可能机制进行了研究。
资料和方法
1.成骨细胞体外张应力加载模型:本实验使用的成骨细胞为MC3T3-E1 小鼠成骨细胞株,在含10%胎牛血清的α-MEM 培养基中常规培养。
成骨细胞按4x105/ml 密度接种于消毒后的加力板中心区域,48 小时后,将培养基换为无血清的α-MEM 培养液继续培养24 小时,以同步化成骨细胞细胞周期。
采用Focrel 四点弯曲体外细胞加力仪器,频率为0.5Hz,加力板变形量为2000μstrain,按0h(未加力),lh,3h,6h,12h,24h 不同时间,使细胞发生单轴牵张应变(图1),加力结束后按组收集细胞。对照组细胞不加力。
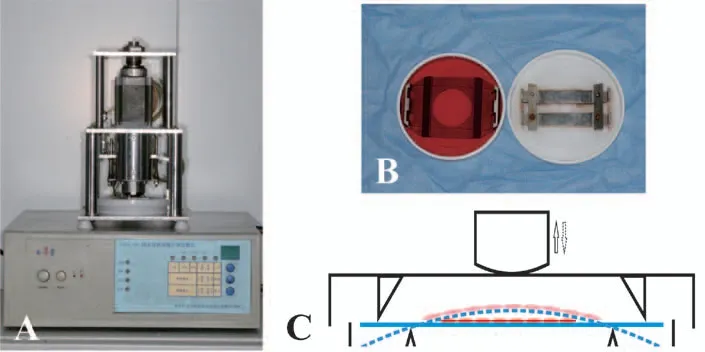
图1 Focrel 四点弯曲体外细胞加力装置
2.成骨细胞增殖活性分析:各组细胞加力结束后,取出加力板,消化收集加力板上的成骨细胞并固定。釆用流式细胞仪(Beckman Coulter)检测细胞DNA 含量,计算各样本中 G0/G1 期、S 期、G2/M 期细胞百分比。增殖指数(PI)反映了细胞群体的增殖活性和DNA 合成速度,分别计算各组的PI 值。PI=(S+G2/M)/(G0/G1+S+G2/M)×100%
3.TGFβ/Smad 及成骨分化相关因子表达检测:①细胞总RNA 提取:按照TRIzol 试剂盒(Invitrogen公司,美国)说明书步骤进行细胞总RNA 的提取,提取所得的总RNA 经琼脂糖凝胶电泳质检合格后备用。②引物设计与合成:根据GeneBank 中小鼠TGFβ1、TGFβ2、TGFβ3、TGFβR Ⅰ 、TGFβR Ⅱ 、Smad2、Smad3、Smad4、Smad7、OCN、ALP、COL-I 和Runx2 基因和GAPDH 管家基因序列设计引物,引物由生工生物工程(上海)有限公司设计合成(表1)。③逆转录合成cDNA:采用Takara PrimeScript®RT reagent Kit With gDNA Eraser 试剂盒(Takara公司,日本)在FTC2000PCR 合成仪上进行逆转录。④实时聚合酶链反应(RT-PCR):采用 Takara SYBR® Premix Ex TaqTMII(Tli RNaseH Plus)试剂盒(Takara 公司,日本)配置反应体系,在ABI PRISM ® 7300 Fast Real-Time PCR System(Applied Biosystems 公司,美国)进行检测,扩增条件均为:95℃,30s(预变性);95℃,5s,60℃,31s,40个循环(PCR 反应)。⑥统计分析:实验结果以平均值±标准偏差的形式表示。采用SPSS17.0 软件包,对PCR 结果在各个时间点进行单因素方差分析,并在此基础上应用LSD 法进行两两比较,规定P<0.05 有统计学差异。

表1 RT-PCR 引物序列
结 果
1. 成骨细胞增殖活性:随着加力时间的增加,G0/G1 期细胞所占比例呈下降趋势;S 期细胞比例在1h,3h,6h 组呈上升趋势,12h 组出现明显下降,而24h 组比上一阶段明显增高;G2/M 期细胞比例在1h,3h,6h 组不断减小,12h 组明显升高,但24h 组再次降低(图2)。
PI 指数随应力加载时间增加而呈上升趋势。除1h 与3h 组外,其他各时间点间PI 指数差异均具统计学意义。0h,1h,3h 加力组与对照组差异无统计学意义,6h,12h,24h 组差异具有统计学意义(图2)。
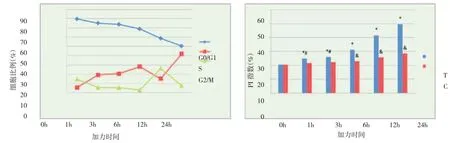
图2 张应力加载不同时间点细胞周期及增殖指数变化
2. 总RNA 样本质检结果:各组总RNA 样品28S、18S、5S 三条带清晰,OD260/OD280 比值均介于1.9 至2.0 之间,表明RNA 有良好的完整性和纯度,无降解及蛋白质等污染,符合逆转录要求(图3)。
3. 机械张应力刺激下成骨细胞ALP、OCN、Runx2、COL-I 及TGFβ/Smad mRNA 的表达:成骨细胞在受力3h、6h 后,其TGFβ1 mRNA 表达水平持续上升,受力12h 后,略有下降但仍明显高于0h对照组,加力24h 后,其表达达到顶峰。TGFβ2 mRNA 表达水平在加力3h 后明显上调,但6h 组,其表达与0h 对照组持平,12h、24h 组其表达再次上升。TGFβ3 mRNA 的表达一直持续在较低的水平,加力6h,其表达明显降低,12h 时略高于对照组,但24h 组再次下降。TGFβRⅠ的基因表达在加力3h后显著上升,6h 时受到明显抑制而下调,但随着加力时间的延长,其表达再次升高,24h 时,达到高峰。TGFβRⅡ的表达变化趋势与TGFβRⅠ类似,即6h时受到抑制,但其24h 组的表达低于12h 组(图4)。
Smad2、3 具有一致的表达变化规律,即加力后3h,表达均上调,6h 时其表达水平与0h 对照组无差异,随着加力时间的增加,其表达水平持续上调,且Smad3 的表达水平在24h 组明显升高。Smad4 在短时加力组的表达水平与对照组无统计学差异,但仍有3h 组表达上升,6h 组受抑制的趋势,在12h,24h组,其表达明显上调。Smad7 在加力各时间点均有较高表达,其高峰位于3h 组。随后逐渐降低,24h 时,略有回升(图4)。
ALP、OCN、Runx2 和COL-I 在加力后3h,基因表达明显上调均高于对照组,但6h 时,其表达受到不同程度的抑制,随着加力时间的增加,其mRNA表达水平呈上升趋势且远远高于对照组(图4)。

图3 各组总RNA 电泳质检
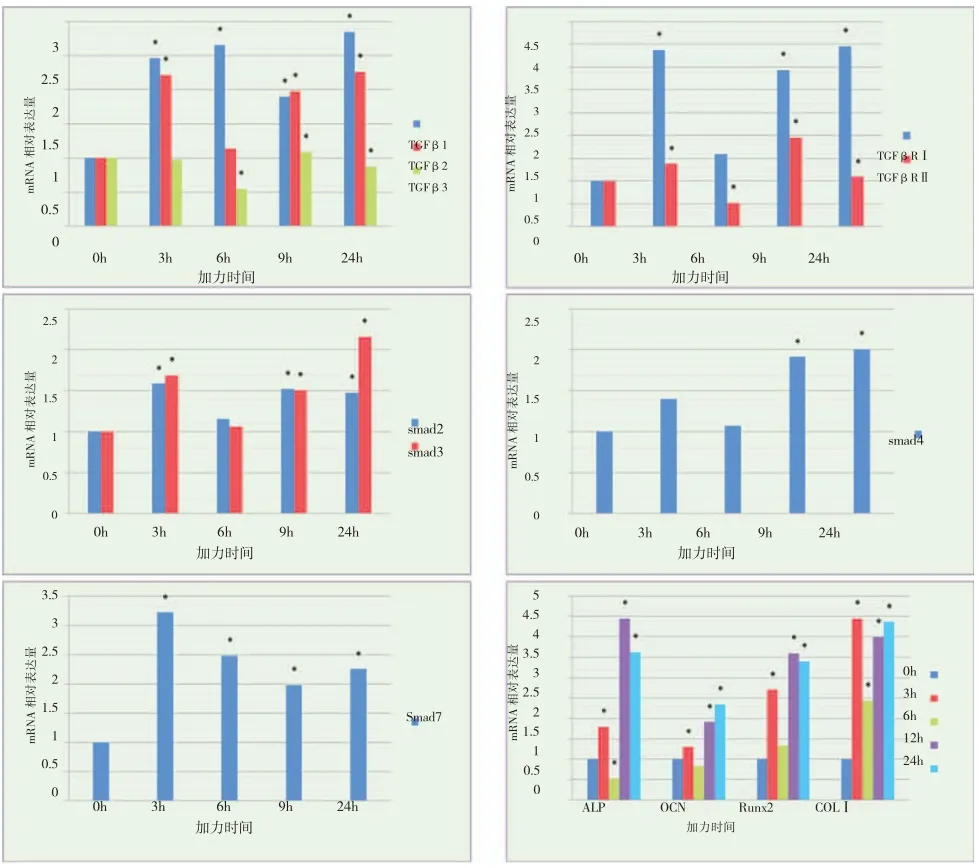
图4 张应力加载不同时间点成骨细胞TGFβ/Smad、ALP、OCN、Runx2 及COL-I mRNA 表达变化(*表示与对照组有统计学差异)
讨 论
在外界机械力刺激下,成骨细胞的反应是骨组织发生骨改建及再生重建的源泉。成骨细胞是对应力应变信号进行感知转导的主要力学敏感细胞,可以通过多种途径感受体内外力学刺激,并将其转化为生物化学信号,介导力相关敏感基因的表达,合成各种酶类等活性物质,从而激活信号网络级联反应,促成一系列复杂的生理病理活动[10]。因此,了解成骨细胞的生物学行为,尤其是在机械应力作用下,其行为的变化规律及机制,对于我们研究机械力作用下,骨骼的改建及再生,具有极其重要的意义。
关于应力刺激对细胞增殖周期的影响,经过无血清培养基培养24h 后,大部分细胞位于G0/G1期。随着加力时间的增加,处于G0/G1 期的成骨细胞比例逐渐下降,提示在这一过程中,进入细胞周期的细胞逐渐增加。加力24h 后,仍有约50%的细胞处于G0/G1 期,这部分细胞可能与维持生物学功能有关。加力1h,3h,6h 后,S 期细胞比例稳定增加,说明在短时应力刺激下,细胞的DNA 合成复制增加。12h 组中,S 期细胞明显减少而G2/M 期细胞比例上升,说明在应力加载12h 后,细胞顺利进入了分裂期。24h 组,S 期细胞比例再次增加,提示细胞进入了下一次的有丝分裂期。PI 指数显示,随着张应力加载时间的增加,细胞的增殖活性增强。但1h 和3h组PI 指数差异无统计学意义,证明在短时应力刺激下,细胞的增殖活性变化不明显,故在后续研究中,我们采用了0h,3h,6h,12h 和24h 作为观察点。
TGFβ 信号通路在软骨内成骨的形成中具有十分重要的作用。Smad 分子作为TGFβ 在细胞内的重要信号转导分子,其在成骨细胞中的表达及机制的报道远远少于TGFβ 及其受体。本研究结果显示,张应力作用下3h 组成骨细胞TGFβ1、2 表达上调,而TGFβ3 的表达无明显变化。加力6h 后,TGFβ1 仍维持较高的表达水平,而TGFβ2、3 的表达则受到了明显的抑制。随着加力时间的延长,TGFβ1、2 的表达回升,并高于对照组,而TGFβ3的表达仍处于较低水平。对成骨样细胞的体外培养实验发现,TGFβ 对其具有直接的、强有力的趋化作用,同时,TGFβ 是强有力的有丝分裂原, 能刺激成骨细胞及软骨细胞的增殖,使细胞外基质蛋白和胶原合成增加并抑制其降解[11~14]。结合本实验结果,我们推测:TGFβ1、2 对于成骨细胞在机械力刺激下增殖活性的增加具有十分重要的作用,尤其是TGFβ1,其表达规律与细胞S 期比例的增加趋势基本一致。而TGFβ2 除在6h 组表达下降外,在其余各加力时间点均维持较高的表达水平。颅骨缝组织块体外实验证实,外源性TGFβ1、2 能促进骨缝细胞增殖、成骨,并最终导致其闭合;而外源性TGFβ3 则会降低细胞DNA 的合成,维持骨缝的开放,在去除TGFβ3 后,细胞DNA 合成增强,骨缝融合[15]。由于颅骨缝的发育主要为膜内成骨,因此我们可以推断,在应力刺激下,成骨细胞TGFβ1、2 的高表达是其增殖指数上升的原因之一。而TGFβ3 的低表达也印证了其虽然与TGFβ1、2 使用同样的受体,却具有不同的生物学效应。
TGFβRⅠ、Ⅱ的表达趋势与其配体基本一致。体内及体外研究发现,TGFβRⅠ的失活会导致成骨细胞早期分化标志物Runx2、Osterix 和ALP 表达的降低。Smad2、3 具有类似的表达规律,在12h 和24h组,其表达维持较高水平,尤其是Smad3。其表达规律基本与其配体TGFβ1、2 一致。同时,Smad2、3 表达规律的细微差异提示我们两者在这一过程中发挥作用的机制及时序可能存在差异。研究显示,Smad2能够完成Smad3 在软骨形成早期的大部分功能,但是Smad3 通过调节TGFβ 信号通路抑制软骨细胞的终末分化从而维持软骨的内稳定。Smad4 作为唯一的通用型Smad 分子,是TGFβ 信号通路的中心介导者。本实验中,Smad4 在短时张应力作用下,具有较稳定的表达,但随着加力时间增加,其表达明显上调。这一表达规律与TGFβ1、2 及Smad2、3 的变化趋势基本一致。因此我们认为,在机械张应力刺激下,TGFβ/Smad 经典信号通路参与了机械应力对细胞的增殖和早期分化的调控。
Smad7 对TGFβ 介导的细胞作用具有重要的调控作用,如细胞增殖与细胞周期阻滞、细胞凋亡和上皮细胞向间充质细胞转变等。在张应力作用下,Smad7 加力后3h 即出现了较高的表达,并随着加力时间的延长而逐渐下降。I-Smads 能够受TGFβ 超家族成员比如TGFβ、Nodal 和BMP 等的诱导而表达,因此形成一个负反馈的机制[16,17]。本实验中,在张应力作用下表达上调的TGFβ 及Smad2、3 和4可能激活了Smad7 的负反馈作用,其在应力刺激前期的高表达,可能是成骨细胞在1h、3h 增殖活性变化不明显的主要原因之一。随着加力时间的增加,其表达下调,成骨细胞增殖指数明显增大。但在24h组,其表达较12h 组有所上升,这可能与细胞进入下一个有丝分裂期,其表达变化受前一阶段细胞分裂的影响有关。由于Smad7 对于TGFβ 和BMP 均有抑制作用,并且参与了多条信号通路的串话,因此,需要更深入的实验来阐明整个作用机制。
本实验对成骨细胞在张应力作用下,ALP、OCN、Runx2 及COL-I mRNA 的表达进行了检测,发现在牵张力作用3h 后,成骨细胞上述基因表达均被上调,这一结果与以往研究一致。在加力后6h,上述基因的表达均下降。这可能是由于持续力作用下,细胞形变达到一定程度,对力学刺激的敏感性降低,而出现了不应期。随着加力时间的延长,上述成骨细胞分化标志物mRNA 表达水平再次上升,并远远高于对照组,提示牵张力能诱发成骨细胞的分化成熟趋势,从而影响了细胞基质合成及矿化。
