运动生理学的学科起源
——基于骨骼肌相关研究的追溯*
2019-07-24苏利强张克峰温岱宗李加鹏陈海春
苏利强,张克峰,2,温岱宗,李加鹏,陈海春
体育学科追根溯源是体育学研究关注的领域之一。目前,较普遍认为运动训练学是体育的本源学科,其他众多学科是随着科学进步和学科分类的交叉重叠而融入体育领域,并成为体育学的重要支撑学科,如运动生理学、运动解剖学等基础课程均是从生理学、解剖学分支出来的,母学科知识融入体育领域的过程也是体育学科丰富发展的历程。随着现代社会的发展,体育对综合性知识需求越来越大,体医融合的落实需要体育学与临床医学的知识做支撑,体育运动项目社会化需要体育学与社会学知识的交叉。这些问题逼着我们“走出体育看体育”,从更广阔的视角来解决体育科学发展面临的问题。“述往事,思来者”,历史是一本教科书,是现实的指南。追溯母学科知识融入体育学的过程,讲述运动生理学诞生前期的科研故事,探寻知识产生、积累与融合之初衷,对正确理解体育学的学科本位有指导意义,对母学科知识与体育学的现代交融有启示作用,对解决新时期体育学遇到的新问题具有借鉴意义。
梳理运动生理学的起源有必要对生理学进行追溯。生理学成为一门科学是在1628年,威廉·哈维出版的一本An anatomical study of the motion of the heart and of the blood in animals,翻译为《心血运动论》[1-2](见图1),书中的血液循环学说被恩格斯评价为这是生理学成为科学的标志,18世纪以后生理学在神经、消化、呼吸以及内分泌诸方面获得了很大的进步,这些成果标志着生理学生根发芽[3-5]。生理学知识的丰富和发展是运动生理学起源的基础。

图1心血运动论及不同版本的译著[1-2] Figure1 The Motion of Heart and Blood and Different Versions of Translated Works(Cited From William Harvey
国内对运动生理学发展史的资料较少,虽记载了第1本运动生理学教材是1889年的《身体运动的生理学》,但对该教材之前的研究提及甚少[6-9],杨锡让[10]、王瑞元[11]曾综述了运动生理学现状与展望,未提及19世纪以前生理学中有关运动的研究。作为一门完整的学科,只有其内部知识体系完整统一、新旧知识演化清晰,才能明确学科发展性特征[12-13]。追溯早期骨骼肌及运动的研究成果,探索这些知识与运动生理学知识体系的逻辑关系,不仅对追溯运动生理学学科起源有历史意义,也对确立运动生理学的学科独立性有重要意义。
本研究运用文献资料法和文本分析法进行研究,检索数据库包括PubMed、MEDLINE、Nature及子刊,检索词为Muscle,检索语言为英语,检索时间为1889年12月31日之前,纳入标准:骨骼肌形态结构、骨骼肌收缩、骨骼肌神经支配、随意运动、疲劳;排除标准:疾病治疗、与本研究主题无关;共检索764篇文献,经过筛查102篇符合要求,其中43篇文献纳入本课题重点研究,通过总结、归纳上述研究成果,结合骨骼肌研究成果与学科知识体系框架,从以下几个方面阐述运动生理学的起源。
1 骨骼肌形态结构和功能的研究
对骨骼肌形态结构的研究经历了从宏观到微观不断深入的过程。19世纪初期对肌肉的认识是通过解剖尸体观察肌肉形态结构[14-16],只到显微镜的发明和使用后,人才逐渐认识到骨骼肌的微细结构,1866年有学者对骨骼肌光学显微镜下的微细结构进行了描述[17],但骨骼肌微细结构界限不太清晰,从显微镜下的视觉效果出发,W.ABBEY[18]在1867年在普通骨骼肌切片的基础上,采用了洋红(Carmine)对骨骼肌细胞进行了染色,从而将细胞核和细胞质与相邻结构通过不同颜色区分开来,使观察到的骨骼肌微细结构更加清晰。随着对骨骼肌微细结构的深入研究,1879年在检查冷冻青蛙的完全伸展肌肉的一些纤维时,显微镜下发现肌肉温度只要低于30℃,横向条纹是消失,但温度略高于30℃,横向暗条开始出现。同时发现纵向减少与肌肉横向测量的增加有关,没有任何体积损失,在静息状态下没有纵向条纹,并且在肌肉收缩期间出现[19],这一成果是基于严密的实验设计,探索了不同温度对肌肉微细结构的影响,从而发现了肌纤维的明带暗带,为骨骼肌收缩的肌丝滑行学说奠定了基础。
19世纪,骨骼肌运动功能的研究是基于医学的需求,随着对骨骼肌相关疾患的探索与治疗,使人们对骨骼肌运动功能的认识有了长足的进步。1871年在肌肉关节疼痛的相关研究中对腓肠肌的起点进行分析,描述了腓肠肌的屈膝功能,并指出了能轻微地帮助胫骨旋转[20];1872年对胸大肌的起点与止点的位置进行了观察,分析胸大肌和周围其他肌肉的位置关系,说明了胸大肌对肩关节的功能[21];1873年对腹肌的分布与结构研究中发现腹肌的后腹部是从深层而来的,前腹是从浅层来的,中间肌腱是原始未分层肌肉横隔的残余[22];1874年,在研究肌肉功能对骨骼发育的影响时,以没有股方肌的病例为对象,发现其第六腰椎的脊柱上第一肋骨不发育[23],肌肉的辅助结构病变影响肌肉的功能,因此对肌肉和肌腱的细微解剖结构进行了研究[24]。从早期对骨骼肌运动功能的描述看,为了解决肌肉相关疼痛疾病,学者在骨骼肌结构的基础上对其机能进行了研究,这些研究不仅有助于治疗骨骼肌疼痛的疾病,同时对骨骼肌运动功能的认识也有较大的促进。可见,人们对知识探求的动力源自于对生活实践的需求,现阶段运动生理学的研究也应该基于运动实践,解决现实生活中人们切实的运动需求问题。
2 生物电与骨骼肌收缩
上述研究从宏观到微细观察层面分析了骨骼肌结构与功能,但形态学解决不了骨骼肌收缩的动态机制,生物电检测技术的发展使人们对骨骼肌收缩的认识向前推进了一大步。1786年L.GALVANI(1737—1798)及其研究团队用带有静电的金属棒触碰了青蛙腿的神经并观察到肌肉收缩,发现肌肉收缩与生物电有关。肌肉收缩程度与生物电的具体关系未知,1867年,学者观察死亡后肌肉的生物电,结果发现发现死亡后的肌肉电流减少与肌肉兴奋性的减弱成正比,肌肉的电兴奋收缩产生的力量(electro-motor force)和兴奋性同时终止,认为肌肉收缩性功能离不开肌肉的电兴奋性[25]。肌肉收缩是可变电流还是恒定电流引发的?1868年,当W.RUTHERFORD用一个恒定电流通过刺激运动神经时,发现肌肉只在电流开始或结束时收缩,在电流恒定时肌肉保持静止状态(见图2),肌肉收缩是可变电流引发的,既然肌肉收缩时和安静时都存在生物电,那么可变电流引发的肌肉收缩时的生物电状态与安静时的生物电状态什么区别?1871年,C.B.RADCLIFFE发现,肌肉收缩时电状态不是静止的,而是流动的[26]。研究者将穿过神经和肌肉的电流引起一种奇特的状态称为负电状态(electrotonic state)或电紧张状态,认为这种神经和肌肉状态的特征在于电势和兴奋程度的改变[27]。当时有学者提出神经和肌肉的生物电状态是由极性分子(peripolar molecules)组成,神经和肌肉的电流源自于极性分子,但是这一观点很多生理现象不能解释,如神经肌肉活动过程中电流的减弱、两个相反的电流如何区分。1871年,T.L.BRUNTON等[28]在这一学说的基础上对肌肉收缩时电流变化的机制进行了进一步的推理,认为肌肉中的电流由不同的原生质之间的差异所产生。1876年,为验证这一学说,G.J.ROMANES采用神经断裂的模型研究神经渗透肌肉的电子条件,结果发现肌肉内的神经终板在电刺激时具有和神经干不同的生物学特性[29]。1878年学者对所有可兴奋的组织进行研究,发现可兴奋组织在刺激作用下作出电-运动反应时,原生质在兴奋部位的电位变为了负值,而没有兴奋的部位保持不变,这对认识骨骼肌收缩时的负电状态有了较大的进步[30]。这一时期的研究从生物电与肌肉收缩现象出发,在控制电流变化情况下了解了肌肉收缩是需要可变电流的,为了解释这一现象,学者提出了学说,认为电流是由不同原生质之间的差异所产生,这一学说为今后的膜电位和动作电位的离子学说奠定了基础。
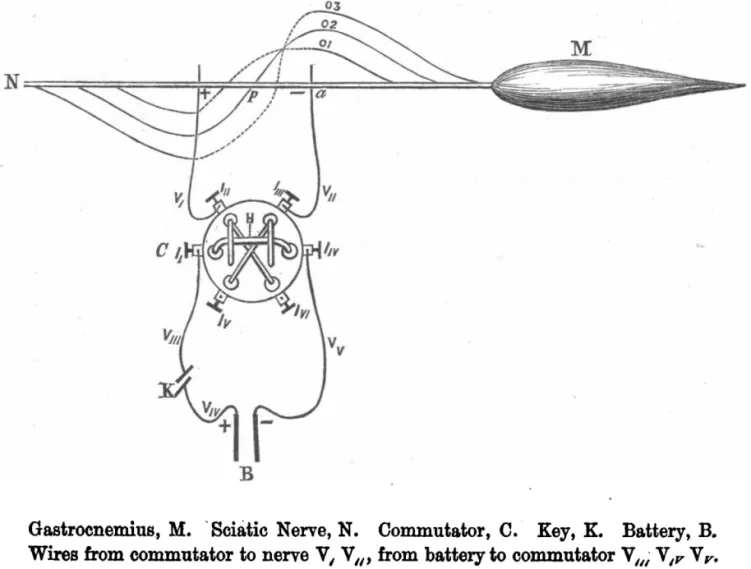
图2电刺激神经监测肌肉活动示意图[26] Figure2 Schematic Diagram of Muscle Activity Monitored by Electrical Stimulation of Nerves[26]
在认识可变电流引起骨骼肌收缩的基础上,学者开始对不同电流刺激骨骼肌进行了研究。1881年J.T.CASH采用磁电机旋转装置产生不同电流,通过反复测试并找到引起肌肉不同负重的最大收缩时的电流,通过肌肉最大收缩的电流刺激肌肉,记录肌肉不同负重的肌肉收缩曲线,结果发现同一肌肉不同负重的肌肉收缩曲线不同,后来采用不同动物进行实验,发现鸽子的胸肌、兔子的肌肉以及青蛙的肌肉存在不同的电刺激-收缩特征,这种电刺激-肌肉收缩的特征在不同动物、不同部位均表现不同特征(见图3)[31]。改变研究中变量条件,获得不同的实验结果,是科学探索的重要手段。正如J.T.CASH[31]的研究,通过不同电流刺激不同骨骼肌,分析电刺激-肌肉收缩的特征,使人们逐渐认识了红白肌的特征,而红白肌的形态机能特征则是当时到以后一段时间内运动生理学研究的重要内容之一。
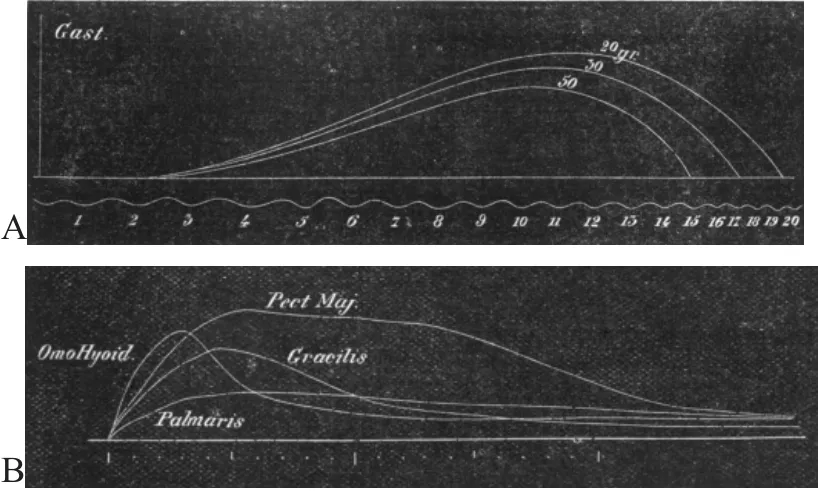

图3不同负重不同肌肉大刺激收缩曲线Figure3 Contraction Curves of Different Muscle Stimuli with Different Weights
3 骨骼肌收缩的潜伏期
随着对电刺激肌肉单收缩研究的不断推进,在描记肌肉单收缩图形时发现从电刺激开始到肌肉收缩总有一定的时间间隔,这一时间间隔是仪器产生的误差还是客观存在的?另外神经的电信号是如何传到肌肉的?基于这些问题学者展开了一系列的研究。1859年,潜伏的肌肉激发期的持续时间(当时没有正式命名为潜伏期)和重要性受到质疑,当时的观点认为运动的有形表达的延迟可能是由于从骨骼肌收缩到描记杠杆传递所需的时间,以及描记图形时使用的书写笔与纸张的摩擦,在当时的测试方法中所有的条件都有可能延迟书写杠杆和书写笔的可见运动,不存在肌肉潜在的激发期的理由是没有任何一个物体的运动速度与电的速度相当。可见当时对潜伏期的认识处在推断阶段,对假设的科学性和合理性有效的质疑可以推动科学的不断进步,接下来的实验,为了避免仪器带来的误差,对多种检测肌电的仪器(见图4~图6)进行了的不断改进,测试方法优化,确定了电刺激与反应的最短时间间隔,图7是当时的仪器和测试出来的“潜伏期”图。这一阶段是对“潜伏期”的质疑与解决阶段。

图4 电信号检测仪示意图[32] Figure4 Cambridge Electric Signal[32]

图5 检测完整肌肉或半块肌肉的潜伏期装置示意图[32] Figure5 Showing the Arrangement of Lever for Comparing Latency of the Whole and Half Muscle[32]

图6 记录肌电反映的仪器示意图[33] Figure6 Records the Instrumental Schematic of EMG[33]

图7 2种仪器记录五通道潜伏期电信号特征图[32] Figure7 Facsimile of Five Tracings of the Two Kinds of Electric Signal Showing its Latency[32]
在确定了潜伏期存在的基础上,学者开始对潜伏期的特征进行了研究。1883年G.F.YEO等对蛙腓肠肌通过改变刺激强度、增加肌肉的负重重量、改变温度观察单收缩的特征,并得出了增加肌肉收缩的速度和力量的合适温度,并对极度疲劳的肌肉进行实验,发现疲劳后的肌肉表现为潜伏期变长、收缩时间延长、曲线高度降低等特征[34]。随着对肌肉单收缩研究的深入,学者也开始对第1次刺激后的第2次刺激产生兴趣,为了检测肌肉收缩特征,对有学者对肌动描记器进行了改进(见图9)。1885年的一项研究采用两次连续的诱导电刺激神经,用肌动描记器记录肌肉变化,结果发现肌肉出现第2次收缩刺激效果的最短间隔是1/600 s,随后采用肌动描记器(专门针对此目的进行调整的摆锤形测量仪)进行了进一步的研究,结果发现最大刺激的间隔只有0.001 s,并且认为获得最大刺激的最小间隔,第2刺激应略强于第1刺激。在此基础上对肌肉产生疲劳的刺激特征和兴奋性特征进行了阐述[35]。

图8 摆式肌动描记器示意图[37] Figure8 Description of Figure[37]
上述对神经刺激肌肉收缩的研究中基本确定了潜伏期的存在,并记录了潜伏期的一下规律,接下来的问题是潜伏期是如何形成的?神经的电刺激信号是如何传递到肌肉的?一些学者提出神经冲动从神经到肌肉收缩的过程可能经历了特定的“中间器官”,这一中间器官可能是潜伏期的形成原因,其中具有代表性的研究是1885年A.W.HOISHOLT等通过直接刺激肌肉来获得最大肌肉收缩与刺激神经干引起肌肉收缩的潜伏期存在很大的差异,推测可能是运动神经末梢刺激过程的延迟,在此过程中,释放的潜在功率将累积,直到其相当于肌肉所需的刺激为止才引起肌肉收缩[36]。
这一时期的一系列研究是以问题为导向,以检测技术革新为基础,从检测技术层面逐步解决了对潜伏期的质疑,当然了解了潜伏期的存在并不是研究的终点,潜伏期产生的机制又成为了下一个新的研究问题,在提出“中间器官”的假设基础上,学者进行了各种实验验证了,这些结果已具有神经肌肉接点的突触传递理论的雏形,也很好的解释了潜伏期产生的机制(见表1)。可见,无论是生理学还是运动生理学的研究,以问题为导向的研究思路是获取知识的重要途径。

表1 骨骼肌兴奋收缩耦联知识发展与追溯Table1 Development and Retrospect of Excitation-Contraction Coupling
4 骨骼肌的神经支配与随意运动
19世纪初,电刺激神经引起肌肉收缩被广泛认可,但神经冲动传达给骨骼肌的方式仍然有争议。一些实验结果显示,在大脑和脊髓被破坏之后心脏仍然会持续一段时间收缩,另外,刺激心脏和肠道神经,其肌肉收缩效应和骨骼肌收缩不同,认为肌肉本身可以产生兴奋收缩,当时这一观点占决定性优势。与之相反的另一种观点是,神经对肌肉收缩有调控作用,电信号通过神经将兴奋传播到肌肉,引起肌肉纤维收缩。在这种争论的背景下,为了验证神经对肌肉的调控作用,一些神经生理学家设计了一系列的实验。1832年,学者采用6周龄的兔子暴露切断坐骨神经,当用电偶导线链接切断神经两端,刺激切断神经的中枢时,可引起肌肉收缩,但这种刺激神经引起肌肉收缩一段时间内会失效,相继的一些实验证明骨骼肌兴奋性和神经力量(nervous power)之间存在着一种基本的关系[38]。肌肉的可刺激性通常被认为是肌肉组织的特性,它在刺激的影响下可以收缩,但离开了神经的支配肌肉的兴奋性将会耗竭,也证明了肌肉收缩是受神经支配的[25],无疑这些研究通过实验证实了骨骼肌收缩需要神经的调控。同时这一争论也推动了人们对心肌和平滑肌的深入研究。可见,在研究神经支配骨骼肌收缩的过程中,科学的争论无疑是促进学科知识发展的动力,严谨的科学实验是验证和解决争论的途径。
对中枢控制与骨骼肌的随意运动的研究经历了从局部到整体的研究历程。电刺激引起骨骼肌的收缩是局部实验的结果,验证了神经对骨骼肌的支配作用,但是人体是完整的有机体,科学研究最终还应回归整体。1886年,V.HORSLEY观察到肌肉反应的节律与神经中枢的电兴奋所产生的节律基本相同,骨骼肌的每一次收缩都是由神经中枢任何部位的兴奋引起的[39]。为了确定神经束分支是否对应相应的骨骼肌,学者通过对骨盆和肩带的肌肉与其支配神经关系的相似性进行了观察和分析,发现当肌肉的位置发生变化时,可通过其供应神经来识别肌肉,认为肌肉应被视为神经的终末器官[40]。这些实验研究成果显示了骨骼肌收缩需要神经支配,中枢兴奋可以引起骨骼肌收缩,从整体上使骨骼肌收缩与中枢控制建立了联系,为开展骨骼肌的随意运动研究奠定了基础。
19世纪中后期,在体动物实验对生理学知识的扩充起到了重要作用,毕竟动物的研究始终不能代替人体的研究,对人体功能和规律的认识仍需以人体作为研究对象。为全方位了解肌肉的收缩,发现用听诊器可以听到骨骼肌收缩时的声音,听到的声音通常归因于周期性活动组织的某些振动,当时认为正常的肌肉音可能被认为是证明正常神经冲动是间歇性的重要环节,连续的自主收缩是由单个收缩的总和所组成的,就像人工强直是由间断的刺激造成的,人们认为收缩骨骼肌产生的声音取决于强直性收缩的张力。1885年的研究通过声学手段以准确的音调进行了研究,发现在自主收缩中听到的音调并不是自然神经冲动不连续的证据,而是仅仅取决于力的变化或刺激分布的颤动[41],1886年,E.A.SCHÄFER研究意识对随意肌节奏反应的影响,发现人的中枢意识产生的神经冲动引起肌肉反映与其他哺乳动物相当,对人体意志刺激的肌肉反应速率与脊髓活动引起的肌肉反应速度相同,人骨骼肌长时间的随意收缩是由每秒8~13次连续神经冲动产生的肌肉不完全强直收缩(见图9)[42]。在探索这些科学问题时,采用不同的研究手段可从不同的角度获取知识,有助于多方位阐释和解决科学问题。

图9 随意肌收缩记录仪[42] Figure9 Apparatus Employed to Record the Contractions of the Voluntary Muscles in Man[42]
随意运动是生理学研究领域,也是神经生物学研究的内容,更是运动生理学研究的基础。神经科学的知识是没有界限的,在运动层面这些知识可以用来解释中枢控制与肌肉收缩的生理机制,从学科角度来看,运动生理学研究也需要掌握中枢控制与肌肉收缩相关的生理学、神经科学的知识,从而更好的解决运动科学中出现的问题。
5 骨骼肌疲劳与工作效率
在农业、手工业时代体力劳动是创造价值的主要手段,生产劳动离不开劳累,劳累是疲劳的一种,虽然当时没有对疲劳明确定义,但19世纪中期已经有相关疲劳与抗疲劳的研究,然而对疲劳的研究也是逐步深入的过程。1863年S.HAUGHTON对长时间静力性收缩引起肌肉不适注意到了这一现象,并对运动时肩关节角度变化与参与肌肉不适的关系进行研究,认为静力性收缩的肌肉维持肢体状态的能量消耗可引起肌肉疲劳,并计算了手臂保持水平时静力性收缩肌肉的能量消耗[43];1871年,S.HAUGHTON始关注肌肉动力性收缩做功情况,他对投掷、抗阻和维持姿势的肌肉效率(Useful effect)与肌肉负荷(weight)进行了一系列的测试,并绘制了函数关系图(见图10)[44],得出了不同运动形式的肌肉效率与肌肉负荷的关系。1875年,F.E.NIPHJER对肌肉疲劳与做功(Mechanical Work)的关系进行多次实验,提出肌肉收缩到疲劳的时间与肌肉工作时的负荷、收缩速度以及肌肉恢复情况有关[45]。肌肉疲劳程度是影响肌肉工作的重要因素,1875年G.HINRICHS研究了肌肉疲劳的评价方法,指出静力性工作时肌肉持固定负荷、保持固定姿势,在肌肉不出现疼痛的情况下维持这一姿势和负荷的时间长短可以评价静力性疲劳,动力性疲劳可以固定负荷,做周期性运动,到肌肉疲劳重复的次数评价疲劳[46]。这些研究通过观察记录相关数据,分析运动与疲劳的关系,获取了一些评价疲劳的简单方法,目前这些方法依然是评价运动疲劳的经典方法。
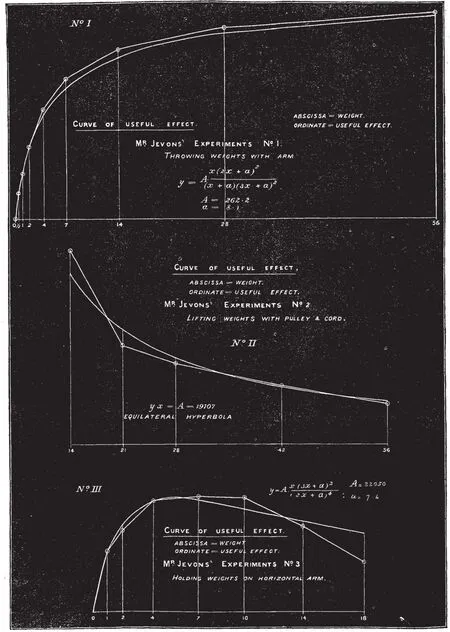
图10 不同运动形式肌肉效率与肌肉负荷曲线[44] Figure10 Curve of relation of Useful Effect and Muscle Load in Different Action[44]
在观察疲劳表象的同时,生理学家没有停止探讨疲劳发生的内在规律,生物电与骨骼肌疲劳的研究也在不断深入,各种组织的电信号检测仪也在不断发展(见图11)。1871年学者采用了当时精密的仪器做了一系列的实验,推导出肌肉兴奋性的变化在一定程度上影响疲劳的变化[27]。1883年发现了极度疲劳的肌肉用电刺激引起收缩的潜伏期变长、收缩时间延长、曲线高度降低[34],在对骨骼肌兴奋性维持的研究时,发现离开了血液的营养,肌肉的兴奋性将会耗竭,认为血液为肌肉提供营养,并可维持肌肉兴奋性[25]。1885年验证了良好的休息可使连续电刺激致疲劳的肌肉收缩能力恢复[35]。1887年,盐水中肌肉频繁收缩引起肌肉收缩功能减弱(既肌肉出现疲劳),向盐水中添加钾盐或石灰盐时肌肉收缩功能加强[47],添加钙盐溶液还可以改善肌肉的状态,并使收缩和放松加速,增加收缩次数[48]。这一时期对运动疲劳的研究历程来看,学者采用观察法认识了疲劳现象,采用实验法从生物电和肌肉兴奋性层面探索了运动疲劳产生的机制,使运动疲劳机制的研究取得了较大进展。
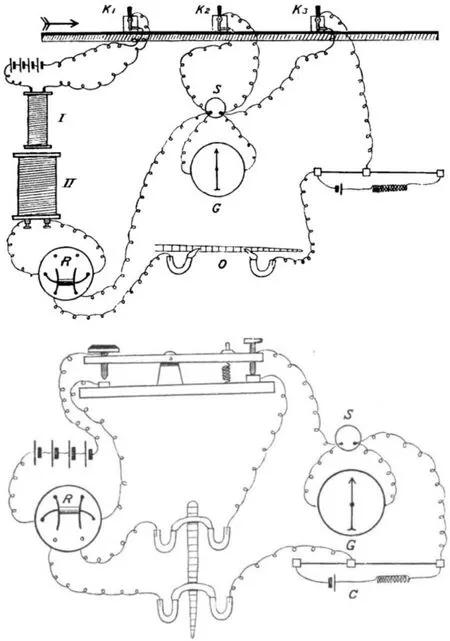
图11 组织电信号检测仪示意图[49] Figure11 Organization of Schematic Diagram of Electrical Signal Detector[49]
疲劳产生的机制与消除是运动生理的主要研究课题,在运动生理学萌芽前期已广泛开展了疲劳的相关研究,当时对肌肉做功效率的研究、对疲劳的评价以及各种体外消除疲劳的方法为后续研究疲劳奠定了基础。运动疲劳是运动生理学学科特征性表现的重要概念,本研究中提供的早期肌肉疲劳素材使疲劳相关知识体系演变的逻辑起点更加清晰,是运动生理学演变为一门独立学科的开端。
6 骨骼肌中糖的发现
肌肉中糖的存在被认为是生理化学领域中最美丽的发现之一。1849年,德国化学家H.FEHLING为了鉴定溶液中还原性的糖存在发明了斐林试剂(fehling's solution),斐林试剂是含有硫酸铜和酒石酸钾钠的氢氧化钠溶液,加热斐林试剂,若无糖存在溶液中会生成黑色的氧化铜沉淀,若有还原糖存在,则产生黄色或砖红色的氧化亚铜沉淀,因此,斐林试剂可通过加热溶液观察沉淀物的颜色来鉴定糖的存在[50]。在1861年8月,梅斯纳宣布肌肉中发现了糖,这一发现被记录在戈廷根皇家科学院题为“关于我们对肌肉中物质的蜕变的认识”的一份声明中。肌肉中糖的存在开启了对骨骼肌功能研究的新领域。处于静息状态和运动状态的肌肉中糖如何变化?1867年的实验显示强直收缩后肌肉中糖含量高于静止的肌肉,还发现运动后肌肉中确实存在一种可发酵的糖,这种糖的量是通过肌肉的运动而增加,并推断肌肉中的糖确实来源于这个器官本身(而不是血液)[51]。这一研究存在诸多不足,如斐林试剂实验中肌酐会对检验糖的结果造成干扰、血糖和肌糖原之间转换关系被忽略等,但也确定了肌肉中重要能源物质糖的存在,并指出了运动与糖发酵有关系,这无疑对骨骼肌运动功能认识的巨大进步。在19世纪中后期对糖和肌肉的关系已有相关实验,能够验证肌肉运动和糖有关系,并能认识到了肌肉中发酵糖的存在,这为研究运动的能量供应和疲劳机制研究提供了突破口(见表2),为运动生理学形成一门学科奠定了基础。

表2 疲劳堵塞学说知识发展与追溯Table2 Knowledge Development and Retrospect of Theory of Sporting Fatigue Jam
这期间还有一些运动生理相关的研究成果相继出现,1871年有实验发现血红蛋白与肌肉活动之间存在一定的关系[27],1878年作者刺激桡神经会引起伸肌反射收缩,导致流量增加,而刺激迷走神经影响呼吸肌不会引起伸肌反射收缩,对血流量增加影响不大[52],1887年的实验显示肌肉发酵物可以加速肌肉凝固[53],1888年发现咖啡因对中枢神经系统的作用可引起肌肉强直[54]。随着对骨骼肌研究的不断深入,使骨骼肌运动功能的知识不断积累,逐步形成了系统化的知识体系,为运动生理学科的形成奠定了基础。
7 小结
17~19世纪生理学家对骨骼肌的研究历程来看,为了解决生活实践活动中肌肉相关疼痛疾病,探索了骨骼肌形态、结构和功能,这些内容属于普通生理学范畴,为了对骨骼肌机能进行深入研究,骨骼肌收缩的生物电、潜伏期和神经支配相关的专门生理学研究随之兴起,这种专门生理学的研究对骨骼肌收缩本质的认识起重要推动作用。同时,在研究骨骼肌潜伏期时学者采用了比较的方法进行探讨,发现了骨骼肌在相应电刺激条件下的不同适应,为全面了解骨骼肌的功能提供了实验支持。比较生理学的进步促进了应用生理学的发展,骨骼肌不同条件下疲劳及消除的比较研究与人类的生产、生活密切相关,所以,对骨骼肌疲劳和工作效率的研究将生理学发展推到了运动生理学层面,因此,运动生理学是在普通生理学和比较生理学等学科知识的基础上,研究生产生活中运动相关的生理现象和规律的一门应用生理学。
1889年,第1本运动生理学教材问世之前,骨骼肌运动功能的研究并非一帆风顺,但也已取得了一些可喜的成果。总结这一阶段研究的经验与教训,对未来运动生理的发展有如下启示:运动生理学的研究应以问题为导向,以解决现实生活中人们切实遇到的运动需求问题是研究出发点和落脚点;对运动生理学中的未解问题进行合理的推论,鼓励开展自由的学术争论,通过严谨的科学实验来验证和解决未解问题,可促进运动生理学知识的丰富和发展;运动生理的研究从运动现象开始,逐步深入到器官、细胞、分子水平,但也要从整体方面考虑机体的调控;1867年,M.ROUGET明确提到使用肌动描记器研究肌肉功能[55],在运动领域;1890年,意大利学者A.MOSSO[56]使用肌动描记器首次记录了肌肉疲劳曲线。因此,解决体育领域的科学问题时,可更早的采用生命科学领域的先进技术和手段,以便运动生理学与生命科学在实验手段上站在同一科学技术水平的高度。
