蓝莓Vco-miR_n10的克隆及对干旱的响应分析
2019-06-12单炳辉柴普今李鹤鹏张琳贺翟璐璐
单炳辉,谢 鑫,李 季,柴普今,李鹤鹏,张琳贺,翟璐璐
(吉林大学植物科学学院,吉林 长春 130062)
【研究意义】MicroRNA(miRNA)是一类参与动植物转录后表达与调控的非编码单链小分子RNA,一般由内源基因编码,长度为18~24个核苷酸,通过碱基互补配对原则降解或抑制靶标mRNA,从而对生物体内的各项生命过程起到重要的调控作用[1]。最早的miRNA(lin-14)是1993年由LEE等在秀丽线虫(Caenorhabditis elegans)中发现的,随后几年中并未有研究报道植物中也有miRNA的存在,直至2002年在拟南芥中发现了miR171,这也是首次发现的植物miRNA[2-3]。随着科研技术的不断完善,越来越多的植物miRNA被鉴定到。截至目前,miRBase(Release 22.1,2018年8月)数据库已释放植物成熟miRNAs 10 414条,共涵盖了大豆(756条)、苜蓿(756条)、水稻(325条)、拟南芥(428条)、马铃薯(343条)、苹果(322条)、葡萄(183条)、番茄(147条)等在内的82个植物物种。越来越多研究表明,在植物生命过程中,这些miRNA作为有效调控因子影响着包括营养器官[4-6]和生殖器官[7-9]发育、植物激素调控[10]、营养平衡[11-12]、胁迫响应[13-14]和miRNA生物合成途径自控[15]等大量生理和生长发育过程。植物生长过程中难免会受到非生物胁迫的影响,引发一系列生理生化反应,可能会降低植物抗逆性、影响品质、减少产量等[15-18]。因此,揭示植物对非生物胁迫的应答反应机制显得尤为重要。【前人研究进展】大量研究表明,植物中一些miRNA会在低温寒冷、盐碱、干旱等非生物胁迫条件下被诱导,其表达量发生变化后进一步调控靶标基因及其下游基因的表达,从而发挥相应的功能。例如,拟南芥中,miR393、miR397b和miR402的表达量会在低温、干旱、盐碱和ABA处理后上升[19]。此外,拟南芥中miR397在高盐条件下表达量增加,其靶标基因LACs和CKB3的表达量降低,该基因过表达后转基因植株的耐盐能力高于野生型拟南芥[20]。Zhang等[21]研究表明,低温胁迫诱导miR319表达量下降,反而提高了植株抗低温的能力。众所周知,干旱是全球面临的主要农业气象灾害之一,其发生频率高、持续时间长、影响范围广、后延破坏力大,严重影响着我国很多地区的农业生产,不仅不利于植株的正常生命活动,而且会降低产品性状,造成较大的经济损失[22],因此,提高植物对干旱胁迫的耐受能力具有重要意义。迄今为止,已发现了大量干旱响应基因,然而这些基因是如何在转录及翻译水平上协调从而发挥功能的仍不是十分清晰,而植物miRNA的功能研究对揭示上述问题提供了大量线索。前人研究表明,干旱胁迫条件下,拟南芥中miR394过表达植株相比于野生型植株对干旱的抵抗能力更强[23]。而miR169则会促进植物叶片水分的散失,从而加速植株枯萎[24]。有研究报道,马铃薯在干旱条件下,miR171会激活SCR基因从而表达SCL蛋白,激活赤霉素信号转导途径,增强抗旱能力[25]。此外,拟南芥中miR156、miR167、miR393、miR396 和 miR408[26], 丹 参中 miR156、miR157、miR166、miR168[27]以 及水稻中miR169、miR393、miR395的表达量在干旱条件下均有提高[28]。对这些表达量变化的miRNAs,它们是否参与植物对干旱胁迫的应答反应?还有哪些miRNA能够调控植物对干旱环境的耐受能力,又是如何进行调控的?这些问题还需要进一步深入研究。【本研究切入点】蓝莓(Vaccinium corymbosum L.)为杜鹃花科越橘属多年生常绿或落叶灌木,其果实呈深蓝色,表皮被白霜,富含花青苷。蓝莓因其为浅根系植物,主根不发达,且根系纤细,在栽培中不能吸收深层土壤的水分,故对土壤水分要求较高,容易受到干旱影响。本研究基于前期高通量测序的结果,从小RNA数据库中筛选到Vco-miR_n10,该新型miRNA在蓝莓果实成熟过程中表达量发生了变化,在蓝果期表达量最高,而在果实生长过程中蓝莓对水分的需求量逐渐增大,此过程中干旱胁迫会严重影响果实品质,我们推测该miRNA可能在蓝莓果实发育尤其是果实成熟期发挥重要的调控作用。【拟解决的关键问题】为了探究VcomiR_n10的基本功能,本研究对蓝莓Vco-miR_n10进行了序列验证,以确定其是否为miRNA,随后克隆并将其转化至拟南芥中,以探究其作用机理,同时,我们利用300 mmol/L甘露醇进行处理,分析Vco-miR_n10对干旱胁迫的响应情况,以期为进一步研究蓝莓Vco-miR_n10的调控功能及提高蓝莓耐旱能力提供重要信息与研究思路。
1 材料与方法
1.1 试验材料
将Col-1野生型及转基因拟南芥种子在4 ℃黑暗环境下春化3 d,播于蛭石与草炭以1∶3比例混合的湿润基质中,随后于22 ℃、16 h/8 h(光照/黑暗)条件下培养。
1.2 试验方法
1.2.1 蓝莓花器官RNA的提取与cDNA的获得 采用RNAprep Pure多糖多酚植物总RNA提取试剂盒(离心柱型)对所采集蓝莓花组织进行总RNA的提取,详细步骤参考试剂盒说明书。反转录详细步骤参照PrimeScript ™ RT reagent Kit with gDNA Eraser(Perfect Real Time)(TaKaRa Bio.,Code No. RR047A)试剂盒说明书。
1.2.2 蓝莓Vco-miR_n10成熟体及前体序列数据来源 序列信息来源于本课题组蓝莓不同发育时期果实小RNA高通量测序的结果(原始数据已上传至NCBI数据库SRX2658700和SRX2676839中)[29],并利用在线软件The mfold Web Server(http://unafold.rna.albany.edu/?q=mfold)对该新型蓝莓miRNA进行二级结构的预测,进一步验证其确为miRNA。
1.2.3 蓝莓Vco-miR_n10基因克隆 设计带有Gateway接头的引物Vco-miR_n10-F(5’-ggggac aagtttgtacaaaaaagcaggcttcGGATTGTTTGACGACGAG AGAGAG-3’)和 Vco-miR_n10-R(5’-ggggacca ctttgtacaagaaagctgggtcACATAGCCCATTTCCACTCA ACC-3’),以蓝莓花组织混合cDNA为模板,利用高保真酶PrimeStar(TaKaRa)进行PCR扩增,程序如下:94 ℃预变性5 min;94 ℃变性30 s;55℃复性30 s;72 ℃延伸30 s,35次循环;72 ℃延伸7 min。琼脂糖凝胶电泳检测PCR产物的特异性。将获得的与预期产物大小一致的单一、明亮的条带进行回收,详细步骤参照天根公司的普通琼脂糖凝胶DNA纯化回收试剂盒(离心柱型)说明书。
1.2.4 表达载体构建 利用Gateway技术将pre-Vco-miR_n10重组到pDONR207入门载体中,然后转化至DH5α感受态细胞中,筛选阳性单菌落培养于37 ℃环境中,提取质粒并进行测序验证。随后,将阳性质粒构建到表达载体pEarleyGate101上,并将构建好的载体转化到农杆菌GV3101中。
1.2.5 拟南芥的遗传转化及转基因阳性苗的筛选利用蘸花侵染法对长势良好且花蕾较多的拟南芥进行侵染,随后黑暗培养16~24 h。T0代种子经次氯酸钠消毒后播种于蛭石与草炭以1∶3比例混合的基质中。生长期间喷施0.2 mg/mL草胺磷进行筛选,10 d后,将生长状况良好、仍保持绿色的转基因植株定植到新的基质中;收集T1代拟南芥叶片,采用CTAB法提取基因组DNA,经过PCR检测上述筛选出的阳性植株是否存在pre-Vco-miR_n10目的基因,以确定幼苗为阳性转基因植株,繁殖并筛选转基因拟南芥至T3代纯合体。
1.2.6 Vco-miR_n10 转基因拟南芥的干旱胁迫 用甘露醇模拟干旱胁迫处理,将WT种子、35Spro::pre-Vco-miR_n10-24和35Spro::pre-VcomiR_n10-52转基因种子消毒后均匀地播种于分别含有0、300 mmol/L甘露醇的MS培养基上,置于光照培养箱中培养,每天定时统计发芽数和绿苗数,连续统计14 d,分别计算发芽率和绿苗率。
2 结果与分析
2.1 蓝莓pre-Vco-miR_n10的克隆
以蓝莓花组织cDNA为模板,利用Gateway技术对蓝莓pre-Vco-miR_n10基因序列进行克隆,克隆片段大小为224 bp,PCR结果与预期一致(图1A),利用Gateway系统中PDONR207和pEarleyGate101依次进行BP和LR反应后,进行菌落PCR检测,结果如图1B所示。其中条带1、条带3和条带4的大小与目的基因一致,条带2的片段长度略小,应为假阳性克隆。随机选取条带1、条带3菌液送去进一步测序,结果显示均为单峰且峰值较高(图1C),表明测序结果可信度很高,测序结果与预期蓝莓pre-Vco-miR_n10基因序列进行比对,结果显示二者几乎一致,错配率较低(图2),且Vco-miR_n10的成熟体序列完全一致。将构建好的质粒转入农杆菌GV3101中,收取侵染后野生型拟南芥种子,再次播种后,通过草胺磷筛选获得阳性植株(图3),T1代种子继续筛选,进一步繁殖后获取T3代种子用于干旱胁迫分析。

图1 蓝莓Vco-miR_n10的克隆、菌落PCR及测序峰Fig. 1 Cloning, colony PCR and sequencing of blueberry Vco-miR_n10

图2 蓝莓Vco-miR_n10测序结果比对Fig. 2 Comparison of sequencing result of blueberry Vco-miR_n10
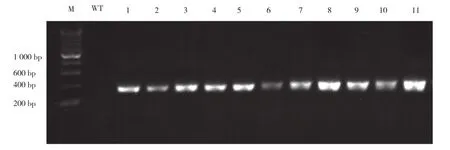
图3 拟南芥阳性转基因株系的鉴定Fig. 3 Identification of positive transgenic lines of Arabidopsis thaliana
2.2 蓝莓pre-Vco-miR_n10的二级结构预测
利用mfold软件对蓝莓pre-Vco-miR_n10的二级结构进行预测,发现其可以形成一个比较典型的茎环结构,且成熟的miRNA序列位于茎环结构的一条臂上、miRNA*落到对应的另一条臂上,两者间仅存在2个错配,同时具有较低的折叠自由能MFE(-75.58),详见图4。

图4 蓝莓pre-Vco-miR_n10的二级结构预测图Fig. 4 Secondary structure diagram of blueberry pre-Vco-miR_n10
2.3 蓝莓Vco-miR_n10的功能分析
为了更好地了解蓝莓Vco-miR_n10基因在拟南芥中发挥的功能,我们将筛选得到的种子在加入甘露醇的MS固体培养基中进行预试验,根据预试验结果,我们挑选了其中最显著的2个株系(Vco-miR_n10-24和 Vco-miR_n10-52) 及 1个甘露醇浓度(300 mmol/L),对这2个株系在此条件下的生长状况进行了分析。结果表明,生长在含有0 mmol/L甘露醇MS培养基中的野生型拟南芥和转基因拟南芥没有差别,0 mmol/L甘露醇下的拟南芥长势明显优于300 mmol/L甘露醇下的拟南芥(图5)。在蓝莓Vco-miR_n10基因的作用下,拟南芥对干旱条件的抵抗能力明显降低(图5)。300 mmol/L甘露醇下的野生型和转基因拟南芥均从播种后第4天开始萌发、第5天萌发速率提高,且从第5天晚上起野生型拟南芥发芽率显著高于转基因株系发芽率,直至第9天发芽率基本趋于稳定。野生型拟南芥的发芽率高达67.72%,相同条件下转基因株系Vco-miR_n10-24的发芽率为33.53%,比野生型拟南芥低34.19%。同样,转基因株系Vco-miR_n10-52的发芽率为37.69%,比野生型拟南芥低30.03%(图6);此外,在300 mmol/L甘露醇MS固体培养基中,野生型和转基因拟南芥从第7天开始绿苗,以一个比较稳定的速率持续增长,从第10天开始野生型和转基因拟南芥开始出现显著差异,到第14天绿苗率达到最大,之后不再变化。转基因株系Vco-miR_n10-24和Vco-miR_n10-52的绿苗率分别达到33.53%和37.69%,同样处理下野生型拟南芥的绿苗率为66.33%,比转基因株系绿苗率分别高出32.80%和28.64%(图7)。上述结果表明蓝莓Vco-miR_n10基因在拟南芥中的过表达会降低拟南芥植株对干旱的抵抗能力。

图5 0 、300 mmol/L甘露醇条件下野生型拟南芥和转基因株系间发芽率比较Fig. 5 Comparison of germination rate between wild type Arabidopsis thaliana and transgenic lines treated with 0, 300 mmol/L mannitol
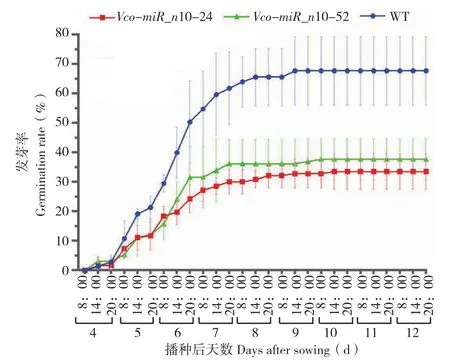
图6 300 mmol/L甘露醇处理下野生型拟南芥和转基因株系的发芽率Fig. 6 Germination rate of wild type Arabidopsis thaliana and transgenic lines treated with 300 mmol/L mannitol

图7 300 mmol/L甘露醇处理下野生型拟南芥和转基因株系的绿苗率Fig. 7 Cotyledon greening rate of wild type Arabidopsis thaliana and transgenic lines treated with 300 mmol/L mannitol
3 讨论
随着高通量测序技术与生物信息学不断的结合与发展,越来越多的miRNA在不同植物中被预测与鉴定。近年来,国内外研究表明成熟miRNAs作为有效调控因子在转录后水平可通过引导靶基因mRNA降解、介导翻译抑制以及DNA甲基化等途径调控靶标基因的表达,从而参与植物的各项生命过程。目前关于蓝莓miRNA的研究报道还很少,本研究中的研究对象Vco-miR_n10是通过蓝莓果实高通量测序获得的一个新型miRNA,关于其调控功能目前尚未见任何报道。为了解Vco-miR_n10的基本功能,第一个关键问题是鉴定该序列是否为真正的miRNA。本研究从两个方面来验证:首先,其前体序列可以形成一个“茎-环”的二级结构,且其成熟的microRNA序列在“茎”的其中一条臂(3′臂或5′臂)上,这与植物miRNA的生物合成机制相一致[30-31];其次,DCL1和HYL1进一步切割pre-miRNA形成miRNA/miRNA*复合体,随后于细胞质中分离为不同长度的miRNA成熟体[32],且该miRNA均为典型的18~24 nt切割产物,而本研究中的Vco-miR_n10成熟体序列大小符合这个标准,即长度约为20个核苷酸,由此表明本研究所克隆的序列确实是miRNA。
在前期研究中,我们对Vco-miR_n10在5个蓝莓果实发育关键时期(绿果初期、绿果末期、白果期、转色期和蓝果期)的表达模式进行了分析,结果显示该新型miRNA在蓝莓蓝果期呈现了最高的表达量[29],说明Vco-miR_n10极有可能在果实成熟过程中发挥重要调控作用。因此,我们利用课题组的蓝莓转录组数据资源结合近缘物种越橘的相关基因组序列,分析并克隆了pre-Vco-miR_n10,并将其转入野生型拟南芥,以期对蓝莓Vco-miR_n10的功能进行研究,除了对获得的T3代转基因株系进行表型观察,我们同时观察拟南芥35S::pre-Vco-miR_n10在甘露醇处理条件下的发芽率和绿苗率变化,结果显示Vco-miR_n10过表达的拟南芥抗干旱能力显著变弱。同样的,Li等[24]研究已表明miR169a和miR169c在植物中过表达会导致植物对干旱更敏感,这与本研究结果一致。与之相似的是,苜蓿缺水时,miR169在根中的表达量下降[33];在拟南芥中,miR169则会通过调控NFYA5来提高干旱胁迫的耐受能力[24];玉米miR168通过靶标基因MAPK在种子中表达,参与种子新陈代谢,进而参与对干旱胁迫的调控[26]。然而,水稻miR169g、miR169n/o[26], 苜 蓿 miR398、miR408[33]与 番茄miR169c[26]在干旱胁迫条件下,表达量均会升高。大豆中OSA-miR319和miR394的过表达会提高植株对干旱的抵抗能力[26,34-35]。此外,gma-miR172c在拟南芥中过表达会提高植株在甘露醇处理下的发芽率和绿苗率[15]。这些研究结果与本文相反,可能是因为不同miRNA的表达水平随着胁迫处理过程而变化,而其靶标基因的不同也会导致调控机制存在差异。本研究从抗干旱胁迫的角度阐释了蓝莓Vco-miR_n10的功能,为蓝莓Vco-miR_n10的进一步研究提供了重要信息和研究思路。
遗憾的是,在整个转基因拟南芥植株生长期间,我们没有观察到转基因植株明显异于野生型植株的表型变化,这可能是因为Vco-miR_n10在拟南芥正常生长状态下的形态发生过程中存在功能冗余,单一提高了该miRNA的表达量并不能引起任何外部形态上的显著变化。后续还将进一步挖掘蓝莓Vco-miR_n10基因在植物包括蓝莓中的靶标基因及其下游基因,以期分析蓝莓Vco-miR_n10基因在植物生长过程中及胁迫条件下的功能调控机制。
4 结论
本研究利用蓝莓转录组、小RNA数据库资源结合近缘物种越橘的相关基因组序列,筛选到蓝莓新型miRNA(Vco-miR_n10),其具有较为典型的“茎-环”二级结构且符合miRNA鉴定的标准。将克隆到的pre-Vco-miR_n10转化到拟南芥中使其异位表达,并观察不同生长环境条件(0 mmol/L和300 mmol/L甘露醇)下的表型变化。结果显示,转基因拟南芥与野生型拟南芥相比,在利用300 mmol/L甘露醇进行的模拟干旱条件下,发芽率和绿苗率呈现比较明显的下降趋势,即蓝莓Vco-miR_n10在拟南芥中过表达降低了拟南芥对干旱胁迫的抵抗能力,使种子在干旱条件下的萌发能力和绿苗率均降低,这可能是由于该miRNA所调控的靶标基因的表达量变化引起的,该结果对提高植物抗旱能力、提升植物品质有着重要的理论指导意义。
