松嫩平原赖草无性系构件生长的可塑性及其规律
2019-05-31洪雪男杨允菲
洪雪男,杨允菲
(东北师范大学草地科学研究所,植被生态科学教育部重点实验室,吉林 长春 130024)
植物属于构件生物,在漫长的进化过程中,植物普遍形成了营养繁殖习性[1-3]。植物通过营养繁殖能够产生与亲株在遗传上完全一致的独立新株,这样的植物称为无性系植物[4-5]。无性系是在自然条件下,由单个亲株通过世代营养繁殖过程所产生的所有构件的集合[6-7]。无性系不仅有与种群数量相似或相同的增长过程,而且也有与种群相同的复杂结构。因此,通过测定无性系构件结构的变化,可以预测无性系发展方向,并进一步了解在无性系繁殖和生产过程中不同构件的贡献大小。许多无性系植物在整个生长季均能够进行营养繁殖,并不断产生新的分株。因此,无性系构件生长有两个过程,一是通过营养繁殖使构件数量增加或无性系增大过程[8],二是构件形态由小到大及生活史阶段的转变过程[9-10]。生物的性状既受遗传物质决定,也受到外界条件的影响,因此数量性状具有一定的生态可塑性[11],而这种可塑性应该遵循着某些规律[12]。通过对不同构件生长可塑性及其规律的研究,能够了解植物种群生长和繁殖策略等重要过程特征,对于种群生长和调节的研究具有重要的理论和实践意义[13-14]。
赖草(Leymussecalinus)是多年生长根茎型禾草[1,15],是典型的无性系植物。赖草广泛分布于我国北方草地,具有耐寒、耐旱等特性,也能忍耐轻度盐渍化土壤,是饲用价值较高的优良牧草。有关赖草的研究,已有栽培育种[16-18]、遗传特性[19]、休眠与耐旱生理[20-21]、种群结构[1,22]、群落生产[23-24]以及不同环境条件下的无性生长等方面[25-27]的报道。本文在生长季末期通过对移植当年的25个赖草无性系的全体分株连同地下根茎完整挖出,调查分析了无性系构件间关系及构件的生长可塑性和规律,揭示赖草无性系在具有充分生长空间而又没有种间竞争条件下赖草无性系的营养繁殖潜力,为无性系植物种群生态学的进一步研究奠定基础。
1 研究地区与研究方法
1.1 样地自然概况
本研究在松嫩平原南部的吉林省长岭县境内,东北师范大学草地生态研究站(44°38′ N,123°41′ E)进行。该地区属于温带半湿润、半干旱季风气候区[28],其气候为典型的大陆性季风气候。年平均气温为4.6℃~6.4℃,≥10℃年积温为2500℃~3400℃。年平均蒸发量在1500~2000 mm之间,年平均降水量280~450 mm,主要集中在6-9月份。样地设在研究站的试验地,土壤为风沙土。
1.2 研究方法
2018年5月初在试验地建立10 m×20 m的试验区,从松嫩平原自然生长的赖草地中,挖取50株连带根茎的赖草无性系分株(分株高度不超过10 cm,根茎长度不超过±2 cm),移栽至试验小区内。为了使赖草有充分的生长空间,单个分株移植的行距株距均定为2 m,按照5行10列长方形样式分布,共定植50个分株,即在试验小区内建立50个赖草无性系。试验区定期清除杂草,但未做施肥、灌水和防治虫害等小区管理。于2018年10月初赖草生长季末期,采用完整无性系挖掘的方法,对试验区内无性系每间隔一个进行取样,共挖取25个赖草无性系,剩下的具有更大空间做其他进一步研究。取样时将地上分株连同地下根茎一并挖出,注意保持各构件的自然联系,每个无性系样本分别装袋。逐个无性系记录分株、根茎和芽的数量;每条根茎测定从根茎节芽形成分株的间隔子长度,计算根茎累积长度。将分株从根茎节处剪下标记分置,在80℃下烘至恒重后,用1/1000电子天平称量分株和根茎生物量。
1.3 数据处理
用各数量指标的平均数(Mean)反映样本的整体水平;用最大值(Max)和最小值(Min)反映样本指标的实际范围;用标准差(SD)反映无性系间的绝对变异度;用变异系数(CV)反映无性系间的相对变异度[29-31]。在构件数量特征的变化规律的分析中,分别用线性函数、幂函数和指数函数3种函数拟合,采用拟合度最高的作为其定量刻画模型。用Microsoft Excel 2007软件对所得的数据进行统计分析和制图。
2 结果与分析
2.1 无性系构件数量特征的可塑性分析
移植的赖草分蘖苗经过一个完整生长季的生长和营养繁殖,便形成了一个个无性系。经统计,在25个无性系中,根茎累积长度平均为3.53 m,最长的达10.32 m,是最短(0.54 m)的19.11倍;根茎节数平均为54.68节;平均间隔子长度为6.97 cm。最大的无性系形成了95个分株,最少的为4个分株,平均31.72个分株。从各数量指标的相对变异度变异系数上看,以间隔子长度的最小为29.4%,分株数量和生物量的均较大,分别为91.7%和103%,根茎芽和分蘖节芽分别为58.9%和107.9%。由此表明,在具有充分生长空间而又没有杂草竞争的条件下,仅经过一个生长季的生长和营养繁殖,赖草无性系各构件数量特征仍然具有较大生态可塑性。
2.2 分株和根茎构件数量特征之间的关系
经统计分析,赖草无性系分株生物量与分株数量之间、分株数量与根茎累积长之间均呈极显著(P<0.01)幂函数关系;根茎生物量与根茎长之间、分株生物量与根茎长之间、分株生物量与根茎生物量之间以及分株数量与根茎生物量之间均呈极显著(P<0.01)线性函数关系(图1,图2)。由此反映了赖草无性系分株和根茎构件之间,在生长和物质生产或积累过程是极为有序的,其中,在幂函数关系中体现了异速生长,在线性函数关系中体现了同速生长。
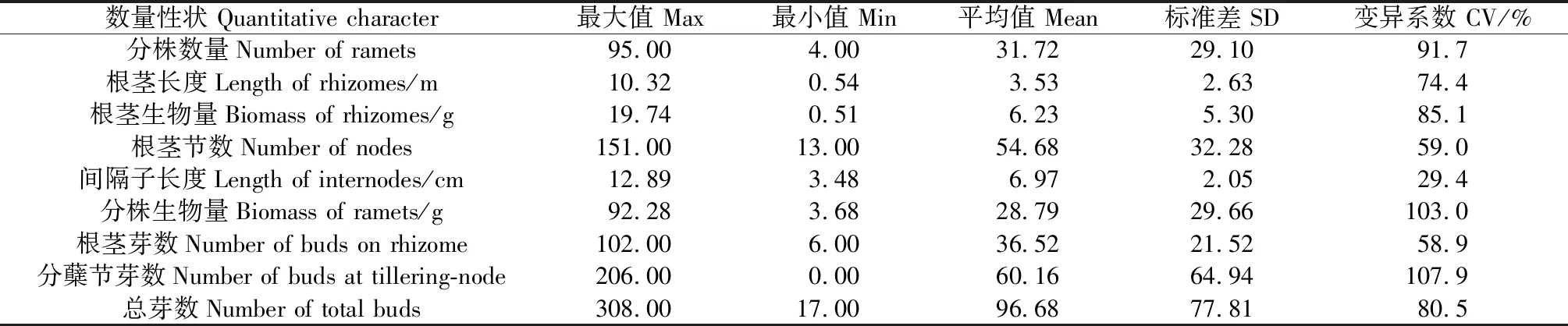
表1 赖草无性系构件的数量特征Table 1 Quantitative characters of the modules in L.secalinus clones (n=25)
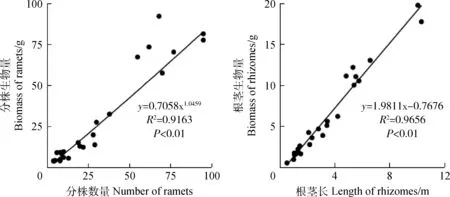
图1 赖草无性系分株数量与分株生物量以及根茎长与根茎生物量之间的关系Fig.1 Relationship between ramet biomass and ramet number as well as rhizome length and rhizome biomass in L. secalinus clones (n=25)
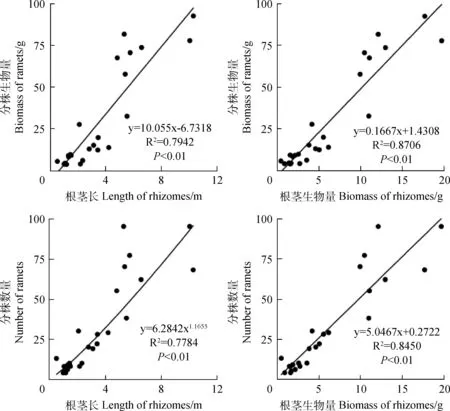
图2 赖草无性系分株与根茎的数量特征之间的关系Fig.2 Relationship between quantitative characters of both ramets and rhizomes in L.secalinus clones (n=25)
2.3 芽数量与分株和根茎构件数量特征的关系
经统计分析,赖草无性系总芽数与分株数量、分株生物量和根茎生物量之间均呈极显著(P<0.01)线性函数关系,而与根茎长度之间呈极显著(P<0.01)幂函数关系(图3);根茎芽数与根茎长和根茎生物量之间均呈极显著(P<0.01)幂函数关系,分蘖节芽数与分株数量和分株生物量均呈极显著(P<0.01)线性函数关系(图4)。由此反映了赖草无性系分株节芽形成的数量与分株构件生长及其物质生产之间体现为同速生长,根茎芽形成的数量与根茎构件生长及其物质积累之间体现为异速生长。
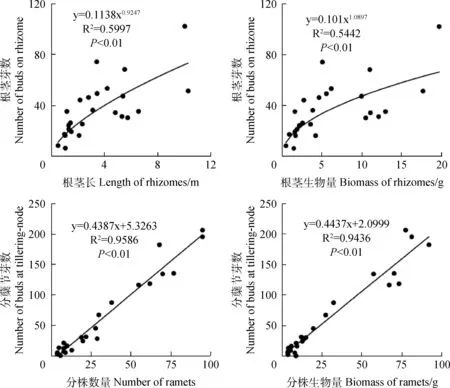
图4 赖草无性系根茎芽和分蘖节芽的数量与分株和根茎构件数量特征之间的关系Fig.4 Relationship between numbers of buds on rhizome and tillering-node and the quantitative characters of modules of both ramets and rhizomes in L.secalinus clones (n=25)
3 讨论
在松嫩平原环境相对匀质的条件下,虽然移植的分蘖苗起始数量相同,但在生长季末期,所形成的赖草无性系构件的数量特征却出现较大差异,如果把数量特征最大值和最小值的差异状况和变异系数作为衡量可塑性的指标,分株和根茎构件的数量特征视为现实无性系生长,芽数量视为潜在无性系大小,本试验中分蘖节芽的可塑性最高,分株和根茎均以生物量的可塑性高于其数量。芽的数量是衡量营养繁殖力的重要指标[32],也是无性系和种群持续更新的基础[33]。据报道,在优势种为多年生植物的草原上,来源于种子的实生苗生长成植株所占比例很少[34],而无性系植物在种群形成过程中有性繁殖并不能起到很大的作用[35],芽数量及变异程度预示着下一个生长季种群的兴衰[36]。依此也可以推测,在下一个生长季,试验区内赖草无性系构件的生长和营养繁殖将产生更大的差异。植物的根茎既是物质的贮藏器官,也是重要的营养繁殖器官[31]。而分株作为物质生产的唯一构件,对环境条件非常敏感,往往采取非常灵活的生长策略。有研究指出,赖草无性系具有以芽优先,分株次之,最后是根茎的构件生长原则[15]。此外,在赖草无性系构件的全部数量特征中,间隔子长度的变异系数最低,仅为29.4%。这可以理解为赖草无性系在营养繁殖和扩展过程中,对其分株的空间分布具有一定的调控功能,其调控机制有待进一步研究。
赖草无性系根茎、分株和芽3种构件间均有着较强的规律性。根据函数性质,通常把线性关系视为同速增长过程,把从加速到缓速的幂函数、从缓速到加速的指数函数等非线性关系视为异速增长过程[29]。在赖草无性系的生长过程中,根茎生物量与根茎长之间呈现为同速生长过程,分株生物量与分株数量之间呈现为加速到缓速的异速生长,这是由于根茎主要受到土壤环境的影响,在相对均一的条件下,单位长度根茎生物量(根茎贮藏力)保持相对稳定的状态,可以保证子代分株的旺盛生活力,尽量减少子代分株的竞争,更有利于种群的空间扩展[37]。而分株除了受到非生物环境等影响外,还与生物环境有关,即随着无性系植物的扩展,生长空间逐渐受限,地上部分的资源利用可能发生改变,从而导致平均单株生物量(分株生产力)发生变化。在分株生物量和分株数量与根茎长的关系中,分株生物量与根茎长之间表现为同速生长,分株数量与根茎长之间表现为异速生长,在无性系扩展过程中,当分株数量增加到一定程度,分株间会存在竞争关系,分株的增长便从加速到缓速。随着根茎的扩展和生长,总芽和根茎芽的数量呈现出从加速到缓速的异速增长规律,随着分株构件的增加,分蘖节芽数呈现出同速增长过程,这可能与赖草的生长繁殖策略有关。
4 结论
在松嫩平原沙地具有充分生长空间的同质园栽培条件下,经过一个生长季的生长,在赖草无性系构件的9个数量特征中,如果用变异系数来衡量其可塑性,间隔子长度的变异系数最小,为29.4%,可塑性最小;分蘖节芽的变异系数最大,为107.9%,可塑性最大;相对而言,分株和根茎构件均以其生物量的可塑性高于其数量。
在赖草无性系中,随着根茎长度的增加,分株数量、总芽数和根茎芽数均呈极显著(P<0.01)幂函数异速生长,而根茎和分株的生物量均呈极显著(P<0.01)线性函数同速生长,体现了赖草无性系具有分株数量优先于根茎伸长的构件生长原则。
