大气CO2倍增条件下冬小麦气体交换对高温干旱及复水过程的响应
2019-05-31郭丽丽张茜茜郝立华乔雅君陈文娜卢云泽李王清涛郑云普
郭丽丽 张茜茜 郝立华 乔雅君 陈文娜 卢云泽李 菲 曹 旭 王清涛 郑云普
1河北工程大学水利水电学院, 河北邯郸 056038; 2河北雄安新区生态环境局, 河北雄安 071700; 3河北工程大学园林与生态工程学院,河北邯郸 056038
自18世纪中后期西方工业革命以来, 大量化石燃料的使用、森林的砍伐以及土地利用方式的转变, 导致大气CO2浓度平均每年增加约2 µmol mol-1[1]。目前, 大气CO2浓度已经超过400 µmol mol-1, 预计21世纪末将继续升高至800µmol mol-1[2]。与此同时, 由于大气CO2浓度升高导致的温室效应引起全球范围内的气候变暖, 到本世纪末全球的平均气温将升高 1.4℃~5.8℃[3]。另外, 气候变暖将进一步影响到全球的降水分配格局, 从而使季节性降水分布不均匀,致使区域性的极端干旱事件频繁发生[4]。以往的气候模型预测结果表明, 大气 CO2浓度升高将增加中高纬度地区的干旱程度和发生极端干旱事件的次数[5], 且高温“热浪”事件的发生频率及范围日益增加[6]。因此, 深入探讨植物对CO2浓度倍增、高温和水分等多个环境因素的协同响应机理有助于提高生态过程模型的模拟精度, 以准确预测未来全球气候变化对不同类型生态系统结构和功能产生的深远影响, 尤其是直接关系到人类粮食安全的农田生态系统对气候变化响应的相关研究备受学者们的广泛关注。
冬小麦(Triticum aestivumL.)为一种广泛分布的C3物种, 也是世界上许多国家或地区最重要的粮食作物之一。作为中国北方地区普遍种植的关键粮食作物, 冬小麦在华北平原区的种植面积高达总耕地面积的 40%[7]。然而,由于华北处于温带季风气候区, 在冬小麦生长关键期的降水量较少, 仅依靠降水远不能满足该区域冬小麦生长的水分供给, 且中国北方水资源相对紧缺, 灌溉用水严重不足[7]。以往研究结果表明, 水分胁迫诱导叶片气孔关闭,限制 CO2扩散过程(气孔限制)和抑制光化学反应过程(非气孔限制)来降低光合速率, 并最终影响植物正常生长和发育[8-9]。通常, 区域干旱伴随着高温天气同时出现, 导致植物气孔导度和蒸腾速率升高, 净光合速率降低, 从而造成农作物的大面积减产[10]。然而, 增加大气CO2浓度可以缓解干旱和高温对植物关键生理生化过程的不利影响,主要是由于提高 CO2浓度对植物产生的“施肥效应”不仅抑制呼吸过程、增加Rubisco酶羧化效率、提高光合速率[11], 同时还能够降低气孔导度、减少水分蒸发, 从而提高作物的水分利用效率和粮食产量[10]。另外, 华北平原粮食产区临时性干旱出现的频率增加, 但干旱期持续时间较短, 水分胁迫时常被阶段性降雨所解除[12]。以往的相关研究还发现, 适度水分胁迫对植物的影响在复水后能够得到一定的恢复, 甚至可能产生超补偿现象[13]。因此, 在CO2浓度倍增和高温条件下研究水分胁迫及复水过程对冬小麦生理生态过程的影响机理有助于为气候变化背景下华北农田的经营和管理提供决策依据。此外, 尽管由大气 CO2浓度升高导致的区域性高温和干旱事件可能会对农田生态系统结构和功能产生协同效应, 但以往大多数研究主要关注 CO2浓度升高、干旱、高温单一或多因素环境因子对作物生理生化过程及产量的影响机理[14-18],以至于目前关于 CO2浓度倍增、高温、干旱及其复水过程对农作物水分利用效率和粮食产量产生交互影响的潜在机理仍不清楚[8,19], 亟待从叶片气孔形态结构特征、气孔空间分布格局、气体交换效率等方面深入剖析农田生态系统对未来气候变化(大气 CO2浓度升高、高温、干旱及其复水过程)的生理响应机制。
本文利用人工气候箱控制不同 CO2浓度、高温和水分条件, 以盆栽冬小麦为试材探讨 CO2浓度倍增条件下高温干旱及复水过程对冬小麦气孔结构特征(气孔密度、气孔面积、气孔形状指数等)及空间分布格局、气体交换参数的影响机制。研究结果不仅为预测未来 CO2浓度升高和高温干旱条件下农业生产力及其服务功能变化提供数据支撑, 而且为农田生态系统气候变化适应性对策的制定提供理论依据。
1 材料与方法
1.1 试验设计
2017年在河北工程大学农业水土资源综合管理与调控实验室进行盆栽试验, 盆栽容器为底部打孔的塑料水桶(桶高27 cm, 桶口面积531 cm2, 桶底面积380 cm2), 且保证桶内土壤水分饱和后能够由桶底顺畅渗出。盆栽土壤为0~20 cm耕层的黄壤土混合营养土(配比2∶1), 土壤容重为1.58 g cm-3, 田间持水量为52%, 含有机质5.26 g kg-1、全氮0.68 g kg-1、全磷0.57 g kg-1、有效磷20 mg kg-1、速效钾58 mg kg-1, 土壤阳离子代换量为15.50 cmol kg-1。冬小麦生长期内不再追施其他肥料。选用华北地区普遍种植的冬小麦(Triticum aestivumL.)品种石麦15, 于3月22日播种, 随后6 d内(3月28日)冬小麦幼苗全部出土。待幼苗生长到三叶一心(4月6日, 即播种后15 d)时定苗, 按大田常规播种密度450万株hm-2的标准折算, 每盆定株24棵。各处理对冬小麦的分蘖影响不大, 且进一步的统计分析结果显示在冬小麦生长期内各处理之间的分蘖数差异不显著(P>0.05)。
采用 4个可以准确控制 CO2浓度的大型人工气候箱(高1830 cm×宽1798 cm×深 675 cm)对冬小麦进行光照培养, 植株冠层的光照强度为1000 µmol m-2s-1, 光照周期为8:00-20:00, 相对湿度控制为60%~70%。各人工气候箱均独立配备一套 CO2浓度自动控制系统, 能够准确控制箱内 CO2浓度(控制精度约为±10 µmol mol-1)。设置对照(CO2浓度为 400 µmol mol-1, 温度为 21℃/16℃, 水分为75%~80%田间持水量)、CO2浓度倍增E (CO2浓度为800µmol mol-1, 温度为 21℃/16℃, 水分为 75%~80%田间持水量)、高温干旱复合处理HD (CO2浓度为400 µmol mol-1,温度为 26℃/21℃, 水分为 45%~50%田间持水量)、CO2浓度倍增下高温干旱复合处理EHD (CO2浓度为800 µmol mol-1, 温度为26℃/21℃, 水分为45%~50%田间持水量)。为避免人工气候箱自身差异对研究结果的影响, 在整个培养期内每周对人工气候箱和冬小麦植株进行随机调换。将盆栽塑料桶内盛满混合营养土, 放入水盆中浸泡5 min,以确保桶内盆栽土壤充分吸收水分, 利用重量法控制土壤水分进行干旱处理。对于干旱处理, 约1周时间土壤水分达到目标含水量即 45%~50%田间持水量。另外, 试验进行水分处理后, 每2 d称重一次, 土壤水分达到下限时,补足到上限。由于本研究主要探讨 CO2浓度和高温干旱复合处理对冬小麦营养生长阶段叶片结构和功能的影响,故播种前并未对冬小麦的种子进行春化处理, 且幼苗在整个营养生长期内没有经历冬眠过程。待冬小麦在不同CO2浓度和高温干旱复合处理下60 d后进行复水处理, 保证每个处理的土壤含水量均达到田间持水量的 100%, 随后进行气孔特征和气体交换参数的测量及数据分析。
1.2 印记法气孔取样及测量
随机从每株上选取 3个叶片, 利用无色透明的指甲油涂于冬小麦叶片远轴面和近轴面的中部, 采集气孔印记样品。在装备有照相机的显微镜(DM2500, Leica Corp, Germany)下观察叶片印迹并照相。随机选择3个不同的显微视野, 每个视野下拍4张照片, 即得到12张气孔的显微照片, 再选取5张来计算气孔密度。利用Auto CAD 2010软件分别测量气孔的长度、气孔宽度、气孔周长、气孔面积以及气孔形状指数。气孔形状指数是指通过计算单一气孔形状与相同面积的圆之间的偏离程度, 当气孔的形状为圆形时, 形状指数为1; 即气孔的形状越扁长, 则气孔形状指数就越大。
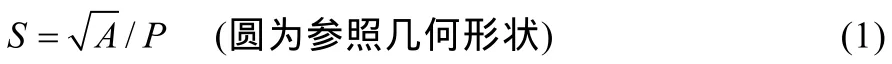
式中,S为叶片气孔形状指数,A表示气孔面积,P表示气孔周长。
1.3 气孔的空间分布格局分析
随机选取4个光学显微照片(放大5倍)用于分析不同处理条件对叶片气孔空间分布格局的影响。在本项分析中,认为每个气孔都是叶片表面上分布的单点, 气孔开口的最中间位置为该单点的位置。首先利用空间分布软件Arc GIS 10.0将照片进行数字化处理, 得到每个气孔的坐标值。再运用空间统计分析方法Ripley’sK方程对表征气孔分布状况的点进行空间分析[16]。Ripley’sK方程是一个分布累加的函数, 该函数利用所有单点距离的二阶矩阵探究这些点在不同尺度上的二维空间分布格局。分析结果由Lhat(d)值来表达。

式中, Lhat(d)表示最小邻域距离;K(d)为任何个体在一定空间尺度内对其他个体的期望值;d为空间尺度。
当分布格局为随机分布时, 所有的d值到 Lhat(d)距离均相等。为了确定95%的可信任区间, 采用蒙特卡洛算法模拟随机分布点1000次。假如叶片表面的气孔在给定尺度d下为随机分布, 则计算出来的值应该位于95%可信任区间之内。假如值大于95%可信任区间, 则气孔在该尺度簇状分布, 当该值小于95%可信任区间时, 气孔在该尺度下为规则分布[16]。
1.4 光合气体交换参数的测定
利用 Li-6400XT便携式光合测定系统(Li-COR Inc.Lincoln, Nebraska, USA)分别测定冬小麦复水前(0 h)及复水后2、4、6、8、10和12 h的叶片气体交换参数即净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(Tr)。与光合测定系统配套的2 cm × 3 cm标准气室可以独立控制光合光量子通量密度(PPFD)、CO2浓度、叶片温度以及气室湿度, 测量时标准气室参数设定为叶室内PPFD 1000 μmol m-2s-1PAR, CO2浓度为 400 μmol mol-1, 叶片温度25℃。利用公式WUE =Pn/Tr计算叶片尺度上的水分利用效率。
1.5 统计分析
不同处理对冬小麦产生影响的各个指标利用单因素或多因素方差分析的统计方法, 再使用Duncan’s Multiple Range Test比较不同处理间的显著性差异(P<0.05)。本研究的统计分析均利用SPSS 13.0 (Chicago, IL)软件完成。
2 结果与分析
2.1 气孔形态特征及其空间分布格局
与对照相比, E显著减小近轴面的气孔宽度和气孔形状指数(P<0.05); 同样, HD也导致近轴面气孔长度、宽度、周长、面积和形状指数的降低(P<0.05), 但却显著增加气孔密度(P<0.05)。高CO2浓度导致高温干旱下叶片近轴面气孔密度、宽度和形状指数的降低(P<0.05), 但气孔长度、周长和面积显著增加(P<0.05)。与对照相比, EHD导致近轴面气孔密度显著增加(P<0.05), 而气孔宽度、面积和形状指数显著减小(P<0.05)。E显著增加远轴面气孔密度和形状指数(P<0.05), 但并未对气孔长度、宽度、周长和面积产生明显影响。然而, HD导致远轴面气孔密度显著增加 33%, 气孔周长减少 8% (P<0.05)。E显著提高气孔长度 13%, 从而增加气孔周长和面积 16%和 20% (P<0.05),暗示E可能缓解高温干旱对气孔造成的影响。与对照相比,EHD提高近轴面的气孔密度45% (P<0.05), 对气孔长度、宽度、周长、面积和形状指数没有产生影响。
对冬小麦气孔空间分布格局分析的结果显示, 无论是对照、E还是HD处理下冬小麦近轴面和远轴面气孔均在小尺度范围内为规则分布(近轴面< 130 μm, 远轴面<170 μm), 而在大尺度范围内(近轴面> 200 μm, 远轴面>230 μm)呈现随机分布的特征(图1)。同时, 在小尺度范围内冬小麦远轴面上的气孔分布比其在近轴面的分布更规则, 即不同处理条件时相同空间尺度上气孔在远轴面分布的 Lhat(d)(均值 -5.3)比近轴面更小(均值 -3.9), 尤其是 E下叶片远轴面和近轴面的 Lhat(d)分别为-8.0和-5.4(图1)。尽管气孔最规则分布的空间尺度在近轴面(90 μm)和远轴面(60 μm)存在差异, 但不同处理并没有对冬小麦叶片近轴面和远轴面气孔最规则分布的空间尺度产生明显影响(图 1)。另外, 不同处理条件时气孔分布格局由规则分布转变为随机分布的空间尺度在近轴面(120 μm)和远轴面(170 μm)之间也存在明显的差异。此外, 与对照相比, E增加了气孔近轴面规则程度, 而HD对气孔规则程度没有产生显著影响, 同时, E并未对高温干旱下气孔规则产生影响。相反, 与对照相比, E导致远轴面气孔规则程度降低, 而远轴面气孔规则程度未受 HD的影响, 但EHD的叶片远轴面气孔规则程度降低。
2.2 气体交换参数
冬小麦的净光合速率(Pn)在E (0 h)条件下比对照提高18%, 但HD (0 h)处理导致Pn下降69%; 同时, EHD处理下, 提高 CO2浓度能够轻度缓解高温干旱对Pn的影响,但仍然较对照低54% (图2)。复水后, 随着时间的推移, E和HD处理的Pn均迅速升高, 而EHD条件下冬小麦叶片Pn升高较缓慢(图2)。另外, 对照组在复水后2 h快速上升,6 h后降低, 但随后呈上升趋势, 并在12 h后叶片的Pn达到最大; E处理下的叶片Pn在复水后4 h达到28 μmol m-2s-1, 6 h后Pn迅速下降, 但随着时间的推移, 叶片的Pn整体呈上升趋势, 且都显著高于复水前。HD处理下Pn与对照组复水结果相似, 也在2 h后迅速上升, 但在6 h降低,其后随时间的延长而上升, 然而复水过程并没有使HD出现补偿效应(图2); 此外, 在EHD处理下Pn缓慢上升, 并在12 h达到最大值, 并高于HD处理, 表明提高CO2浓度能缓解高温干旱对Pn的伤害(图2)。
气孔导度(Gs)在E (0 h)出现降低趋势, 但较对照差异并不显著; 而HD (0 h)导致Gs显著下降,Gs值接近0, 下降幅度为82% (图3)。EHD (0 h)处理下的Gs增加了83%,但仍较对照减小67% (图3)。复水后, 对照组叶片Gs在复水2 h后迅速增大, 其后波动上升, 并在复水12 h后达到最大值0.87 mmol m-2s-1, 而E导致Gs在4 h达到最大值0.97 mmol m-2s-1(图3)。在HD处理下, 复水2 h后叶片Gs升高, 并随着复水时间的推移整体呈上升趋势。
与对照相比, 冬小麦的蒸腾速率(Tr)在E (0 h)条件下降低, 但差异并不显著; 而HD使Tr显著降低69% (图4)。在EHD处理下, CO2浓度倍增导致高温干旱复合处理下叶片的Tr值增加, 增幅为57%, 但仍显著低于对照51% (图4)。另外, 复水2 h后, 对照组叶片Tr迅速增大, 随着时间的推移,Tr始终在缓慢上升, 并在复水后12 h达到最大值。而E导致叶片的Tr在复水4 h后达到最大值, 且整体呈现上升趋势, 并在复水12 h后与对照组相近(图4)。此外, 复水后HD处理叶片的Tr值上升, 6 h后呈下降趋势, 但其后随着时间的推移,Tr值上升(图4)。在EHD处理下, 复水后CO2浓度倍增使高温干旱复合处理叶片的Tr值迅速升高, 并随着时间的推移,Tr值始终呈上升趋势, 在12 h后达到最大值。
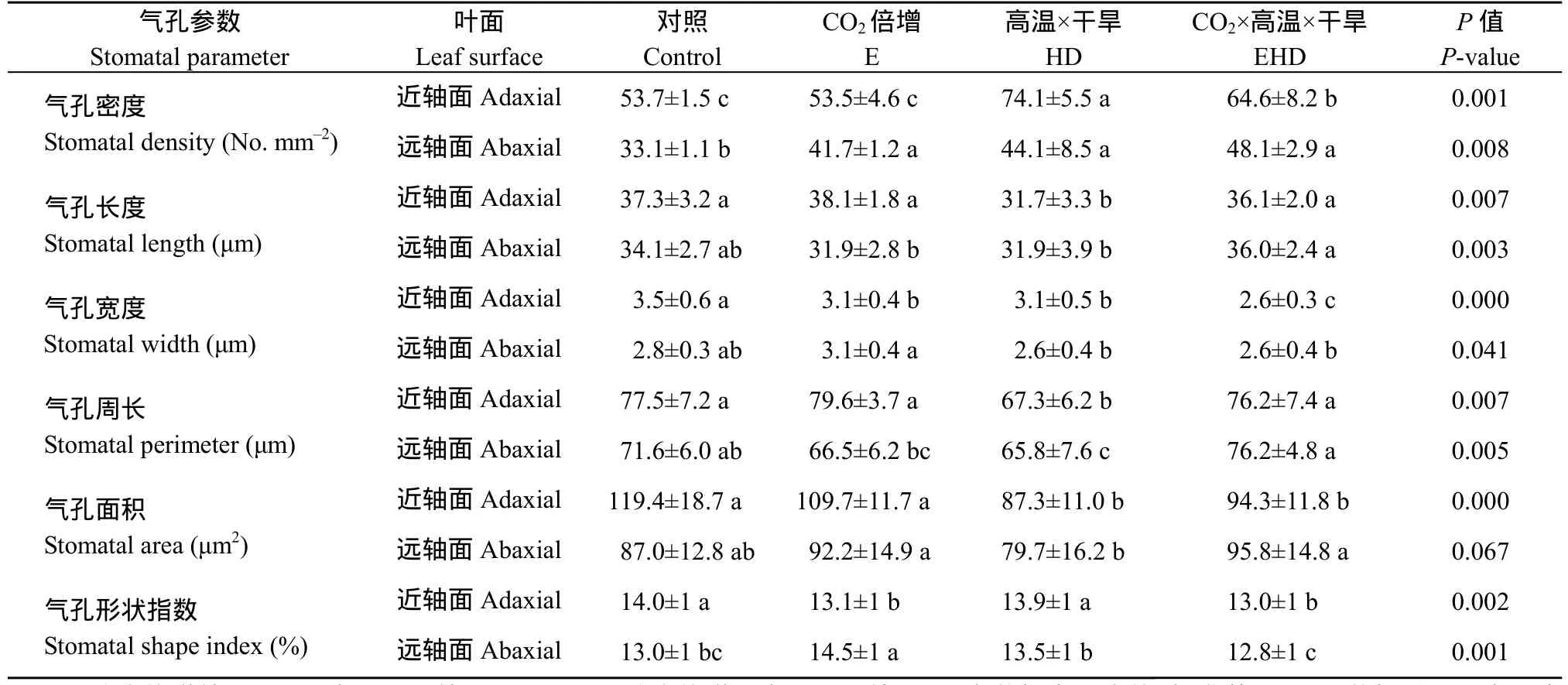
表1 CO2浓度倍增下高温干旱对冬小麦叶片气孔参数的影响Table 1 Effects of the combined stress of high temperature and drought on stomatal parameters of winter wheat under doubling CO2 concentration
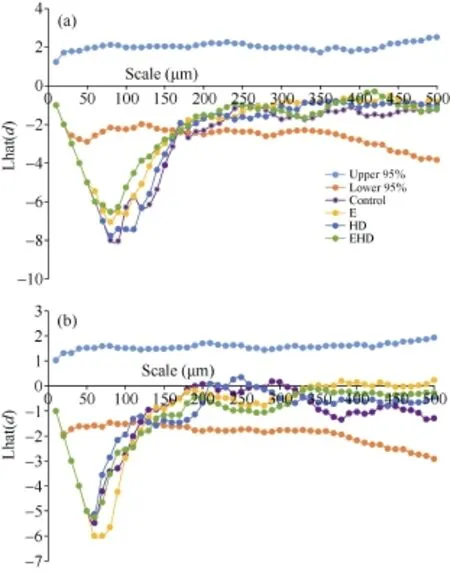
图1 大气CO2浓度倍增下高温干旱处理对冬小麦叶片近轴面(a)和远轴面(b)气孔空间分布格局影响Fig. 1 Effects of the combined stress of high temperature and drought on spatial distribution pattern of stomata on the adaxial (a) and abaxial (b) leaf surface of winter wheat under doubling CO2 concentration
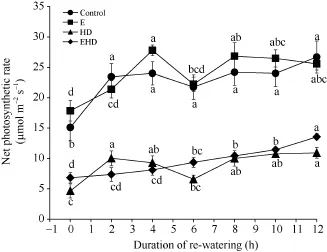
图 2 大气 CO2浓度倍增下高温干旱及复水对冬小麦净光合速率的影响Fig. 2 Effects of the combined stress of warming and drought as well as re-watering on the net photosynthetic rate of winter wheat under doubling CO2 concentration
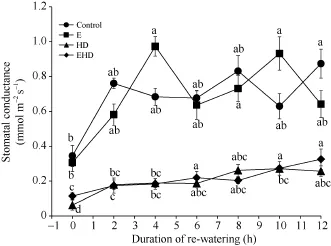
图 3 大气 CO2浓度倍增下高温干旱及复水对冬小麦气孔导度的影响Fig. 3 Effects of the combined stress of high temperature and drought as well as re-watering on stomatal conductance of winter wheat under doubling CO2 concentration
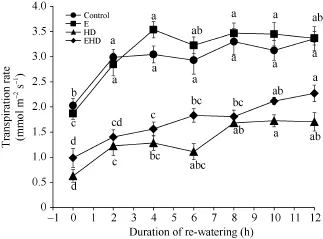
图 4 大气 CO2浓度倍增下高温干旱及复水对冬小麦蒸腾速率的影响Fig. 4 Effects of the combined stress of high temperature and drought as well as re-watering on transpiration rate of winter wheat under doubling CO2 concentration
与对照(0 h)相比, E(0 h)导致叶片的水分利用效率(WUE)增加30%; HD (0 h)减少了叶片WUE, 但差异不显著(图 5); 然而, 提高 CO2浓度对高温干旱处理(0 h)的叶片WUE并没有产生显著影响(图5)。复水后, 对照组的叶片WUE在4 h后迅速上升, 随着时间的推移, 叶片WUE整体呈上升趋势。然而, E在复水后2 h, 叶片WUE迅速下降, 并随着时间的推移, 叶片WUE呈下降趋势(图5)。复水2 h后, HD使叶片WUE上升, 然而, 随着时间的推移, 叶片WUE迅速下降, 并在复水6 h后叶片WUE达到最小值(图5)。复水后, CO2浓度倍增使高温干旱复合处理下的叶片WUE显著下降, 并在复水后6 h达到最低, 随着时间的推移, 叶片WUE呈上升趋势(图5)。
3 讨论
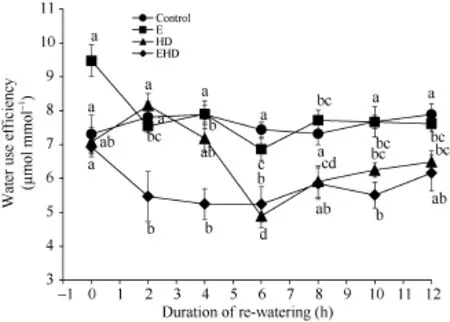
图 5 大气 CO2浓度倍增下高温干旱及复水对冬小麦水分利用效率的影响Fig. 5 Effects of the combined stress of high temperature and drought as well as re-watering on water use efficiency of winter wheat under doubling CO2 concentration
气孔是植物同外界环境进行水分和气体交换的重要门户, 叶片上气孔对开口大小、数量及其分布状况的调节功能是植物适应环境、抵御外界胁迫的一项重要机制[16]。本研究结果显示, 增加 CO2浓度导致冬小麦远轴面气孔密度增加, 气孔长度减小。然而, 近轴面气孔密度却没有显著影响, 但气孔宽度显著减少从而导致气孔形状指数降低, 表明冬小麦叶片的远轴面和近轴面对 CO2浓度的响应并不完全一致, 存在明显的差异。另外, HD导致近轴面和远轴面气孔密度均显著增加, 但仅近轴面气孔长度、宽度、面积和周长均显著减少, 远轴面没有显著变化, 表明冬小麦叶片近轴面气孔对高温干旱复合处理的响应可能比远轴面更加敏感。在EHD处理下CO2浓度倍增导致高温干旱复合处理下冬小麦远/近轴面的气孔长度、周长和面积均显著增加, 表明提高 CO2浓度在一定程度上缓解了高温干旱复合处理对气孔特征的负效应[20-23]。此外,植物叶片的气体交换过程不仅受气孔大小和气孔密度的影响, 而且还与气孔的空间分布格局紧密相关[24]。以往的相关研究发现, 陆生植物叶片的气孔起初并不是随机分布而是规则分布, 从而确保水分丢失和碳同化之间的最佳平衡状态[25]。更为有趣的是, 许多植物在面临干旱、高温和CO2浓度升高等外界环境胁迫时会将气孔的分布格局调整成簇状分布, 从而降低叶片的气体交换效率[18,25]。本研究结果显示, E导致近轴面气孔分布更加规则, 但远轴面气孔分布规则程度降低, 表明冬小麦近轴面比远轴面具有更高的气体交换效率, 即近轴面的气孔空间分布格局对提高叶片气体交换效率贡献更大。另外, 提高 CO2浓度对高温干旱复合处理下近轴面气孔分布没有显著影响, 反而降低远轴面的气孔分布的规则程度, 表明在高CO2浓度条件下叶片远轴面的气孔分布状况对高温干旱的响应更为敏感。
以往大量研究结果表明, CO2浓度升高可以增加净光合速率, 同时降低气孔导度和蒸腾速率[26], 从而提高水分利用效率[27-28]。本研究结果表明, E不仅提高冬小麦的光合速率, 而且降低气孔导度和蒸腾速率, 从而提高叶片尺度的水分利用效率。本研究中 CO2浓度增加导致冬小麦气孔开度的减小, 直接解释了降低气孔导度和蒸腾速率, 提高水分利用效率的原因。此外, 高温和干旱两个环境因子对植物造成的伤害具有叠加效应, 且植物基因和代谢途径对高温干旱处理的响应也不同于单一胁迫[29-30]。干旱或高温胁迫通常降低光合反应效率的原因主要由部分气孔关闭而导致的气孔限制和光合反应位点同化能力的下降所引起的非气孔限制[26]。本研究结果显示, 高温干旱(HD)导致叶片光合速率严重下降, 气孔导度和蒸腾速率均显著降低, 表明HD处理下气孔限制是光合速率下降的主导因素。同时, 在HD处理下冬小麦叶片的气孔长度、宽度、面积和周长均显著减少, 表明高温干旱复合处理下的气孔限制主要是由气孔开度的减小而引起的。另外, 以往研究结果表明提高 CO2浓度会导致植物气孔导度降低,蒸腾速率减小, 随之减少水分消耗[31]。因此, 提高CO2浓度可以缓解干旱胁迫, 提高植物对干旱的适应能力[32]。然而, 值得注意的是升高温度会增加气孔导度和蒸腾速率,增加土壤蒸散, 从而导致总耗水量增加[15]。本研究结果显示, 高温干旱处理下提高CO2浓度(EHD)导致气孔导度增大, 从而增加冬小麦的净光合速率, 表明高CO2浓度在一定程度上缓解高温干旱对冬小麦的伤害程度。本研究中,EHD处理下CO2浓度倍增导致高温干旱时冬小麦远/近轴面的气孔长度、周长和面积均显著增加, 但气孔在近轴面的分布状况没有变化, 甚至远轴面的气孔分布规则程度降低, 暗示提高 CO2浓度对高温干旱处理的补偿效应主要通过调整气孔的形态特征来实现, 而并非由气孔在叶片上空间分布格局的变化而决定。
植物在高温或干旱胁迫下其体内的水分运输系统会通过自身的调节来保障植物正常的生长发育[33]。以往的研究发现, 植物在长期的适应和进化过程中, 不仅逐渐形成了对干旱、缺氧、冷、热等各种逆境的抵抗能力, 并且在逆境状况得以改善时其气体交换功能和生长发育过程还能在一定程度上得到恢复, 甚至还可以达到或超过未经胁迫或伤害下的情形, 从而弥补逆境造成的伤害, 即表现出明显的补偿或超补偿效应[10]。本研究结果显示, 复水后不同处理条件下冬小麦叶片光合气体交换参数均有所升高, 尤其是在E处理下, 净光合速率、气孔导度和蒸腾速率在复水 4 h后均高于对照, 表明在水分充足条件下,提高 CO2浓度对小麦叶片光合速率产生一定的“施肥”效应, 该结论与Wu 等[34]的研究结果一致。然而, 在复水2 h后, HD处理导致Pn、Gs和Tr增加, 但随后气体交换参数的变化均不明显, 这可能是由于高温干旱处理已经导致叶片光合反应位点的结构和功能遭到破坏[35], 故复水并不能对冬小麦结构和功能的恢复产生补偿效应, 这可能是由于复水时间较短(仅几个小时), 叶片的气孔密度、气孔长度及其空间分布格局不可能在较短时间做出调整,而只能依靠气孔开口宽度的变化响应环境的短期变化;由此看来, 气孔通过调整开口宽度提高叶片气体交换效率的能力非常有限。此外, 本研究中发现 EHD处理在复水后叶片光合气体交换参数没有显著变化, 这也暗示高温干旱处理导致植物光合反应位点同化能力的丧失[36]。
