岷山摩天岭北坡木本植物功能性状的变异特征
2019-03-26何靖田青何桂萍宋玲玲曹雪萍
何靖,田青,何桂萍,宋玲玲,曹雪萍
(甘肃农业大学林学院,甘肃 兰州 730070)
植物功能性状(plant functional traits)是指在植物个体水平上通过影响植物生长、繁殖和存活能力从而间接地影响其适合度的形态、生理以及生活史特征[1].最常见的植物功能性状分类包括:形态性状(morphological traits)和生理性状(physiological traits)、营养性状(vegetative traits)和繁殖性状(regenerative traits)、地上性状(aboveground traits)和地下性状(belowground traits)、影响性状(effect traits)和响应性状(response traits)以及后来被广泛接受的软性状(soft traits)和硬性状(hard traits)[2-3].叶片作为植物的营养器官,与环境接触面积最大、对环境刺激最敏感,其功能性状与植物对资源的利用能力及效率紧密相关[4].比叶面积(SLA)与植物相对生长速率、光合速率、叶氮含量有关,是生理过程权衡的最佳指示者[5-8];叶干物质含量(LDMC)是反映植物生态行为差异的另一个重要指标,常与潜在相对生长速率负相关,是资源获取轴上最稳定的变量;叶氮含量(LNC)和叶磷含量(LPC)是植物生长过程中必须的营养物质,氮含量的高低直接决定着植物叶片光合作用的强弱[9],而植物光合作用同化的碳为植物生理生化过程提供能量和底物[10].近年来,科学家逐渐发现,植物体在生态系统中所起的作用可能是由于该植物体具有的某类功能性状使得它可以获取更多的有效资源;而不同植物体功能性状的相异性使得它们具有不同获取和利用资源的策略,从而减少了生态位重叠,增加了多样性及维持生态系统功能的稳定性[11-12].也有研究表明,同一生态系统过程受功能性状组合的影响,而某一重要功能性状则在多个生态系统过程中起关键作用[13].因此,研究植物性状间的变异特征不仅能够了解植物在不同环境的生态策略差异,而且可以更加深入地探索生态位分化和物种共存的机理,有助于进一步深入揭示功能性状的环境指示意义.近年来,国内外学者从植物功能性状随环境因子如海拔[14]、温度、水分[15-16]和干扰程度[17]等进行了研究,而对植物功能性状的变异与植物科类群、高度、生活型、花序、果型、分布区等的研究较少.
甘肃省白水江国家级自然保护区的主体区域—摩天岭北坡森林植被区,西邻青藏高原、东接秦岭山地、南近横断山脉和华中地区.多方植物区系在此交汇,地带性植物群落组成相当复杂,从山麓到高山,拥有中国亚热带、暖温带、中温带和寒温带的多种代表性群落类型,相对高差较大,具有浓缩的环境梯度和高度异质化的生境,气候和植被垂直分布明显,是岷山至秦岭物种基因交流和过渡的重要通道[18-20].宋玲玲等[21]对岷山摩天岭北坡东南部植物群落α多样性与地形特征的关系研究发现,乔木层植物在中海拔地带多样性较低,在低海拔和高海拔区段内群落多样性较高.张守昊等[20]对摩天岭北坡中段不同海拔梯度草本植物多样性生态系统功能分析研究发现,土壤有机碳和全氮含量对草本群落地上、地下生物量的影响显著[20].目前,摩天岭北坡木本植物功能性状的研究主要侧重于植物叶性状与海拔的关系[19,21-22],而有关植物功能性状间的变异特征和营养性状与繁殖性状间的关系的相关研究较为缺乏.
本研究以白水江自然保护区邱家坝保护站(海拔2 338~2 916 m)37种木本植物为研究对象,对植物叶性状包括比叶面积(SLA)、叶干物质含量(LDMC)、叶氮含量(LNC)、叶碳含量(LCC)、叶磷含量(LPC)的变异特征进行研究,并探讨叶性状变异与植物科类群、高度、生活型、繁殖性状(包括花序、花色、果型)及分布区的关系,旨在揭示摩天岭北坡植物叶性状的种间分异特征及其植物功能性状的变异规律,为植物功能性状的深入研究提供参考依据.
1 材料与方法
1.1 研究区概况
研究区位于甘肃省白水江国家级自然保护区邱家坝保护站(E 104 °19 ′~104 °32′,N 32 °55 ′~32 °94 ′),平均海拔1 475 m,年平均气温15.6 ℃,最热月(7月)25.1 ℃,最冷月(1月)4.4 ℃,年平均降水量为840~950.3 mm,年内分布不均,70%的降水主要集中在6~8月.该区在甘肃省内太阳辐射量最低(4 800~5 000 MJ/(m2·a),日照时数约为1 800 h,是水热组合条件最佳的地区.地带性植物群落组成非常复杂,从山麓到高山,拥有亚热带、暖温带、中温带和寒温带的多种代表性的群落类型.其中海拔2 338~2 916 m区域植被以针阔叶混交林带为主.基带土壤为山地黄棕壤.
1.2 试验材料
以摩天岭北坡邱家坝地区37种植物(隶属15科)为研究对象(表1),选择生长状况良好、病虫害干扰程度较小的枝条,带枝采摘,然后采集完全展叶并充分接受日光照射且没有病虫害的叶片,每1个样点每种30片.
1.3 试验方法
在摩天岭北坡邱家坝地段,采用样线和样地相结合的调查方法,设置1条样线,其选址尽量避开人为干扰较大的地段、迹地和大型林窗,并且选择林相相对整齐的植物群落,样线内按照海拔每升高大约100 m设置1个20 m×20 m的乔木样方,并在每个乔木样方对角线2个角上分别设1个10 m×10 m的灌木样方,共计7个乔木样方,14个灌木样方.记录样方内出现的木本植物种类.
将采集的带枝叶片包入打湿的信封袋中放入封口塑料袋内封口,超过24 h放入温度为2~6 ℃的冰箱内,带回实验室测其SLA、LDMC、LNC、LCC、LPC.对植物高度进行野外实地测量和记录,并将其归为灌木和竹类(0~5 m)、小乔木(5~10 m)、中乔木(10~20 m).根据《中国植物志》对37种植物的科类群、生活型、花序、花色、果型进行检索,并进行分类统计.根据《中国种子植物区系地理》对37种植物的分布区类型进行统计[23].
1.3.1 叶性状测定 比叶面积SLA(m2/kg):利用Delta-T叶面积仪扫描测定叶片面积(A)[24];比叶面积SLA=A/M[24],M为叶片干质量.
叶干物质含量LDMC(g/g):分别称量叶片的饱和鲜质量和干质量[24].LDMC=M/m,m为叶片饱和鲜质量,M为叶片干质量.
叶片全氮含量LNC(g/kg):采用CHNS-O元素分析仪[24]进行测定.
叶片全碳含量LCC(g/kg):采用CHNS-O元素分析仪[24]测定.

表1 37种植物信息统计
叶片全磷含量LPC(g/kg):用酸溶-钼锑抗比色法,用UV2550紫外分光光度仪测定[24].
1.3.2 变量分类 科类群 将37种木本植物归为15个科(表1).
生活型:分为2类,落叶植物;常绿植物.
花序:分为8种,Ⅰ类:柔荑花序;Ⅱ类:圆锥花序;Ⅲ类:聚伞花序;Ⅳ类:伞形花序;Ⅴ类:伞房花序;Ⅵ类:总状花序;Ⅶ类:复伞房花序;Ⅷ类:裸子植物球花.
花色:分为9类, i黄白色;ii黄色;iii黄绿色;iv白色; v紫黄色;vi淡红色;vii紫色;viii绿色;ix红色.
果型:分为10类,(1)坚果;(2)球果;(3)核果;(4)梨果;(5)瘦果;(6)蓇葖果;(7)翅果;(8)蒴果;(9)浆果;(10)颖果.
分布区:根据吴征镒(1991)《中国种子植物区系地理》[23],将37种植物归至 8个分布区,1:北温带分布;2:东亚(东喜马拉雅-日本)分布;3:热带亚洲和热带美洲间断分布;4:北温带和南温带(全温带)间断分布;5:中国-喜马拉雅分布;6:东亚和北美洲间断分布;7:旧世界温带分布;8:中国特有分布.
1.4 数据处理
用描述性统计对植物科类群、高度、生活型、花序、花色、果型、分布区进行统计;以相关性分析(correlation analysis)评价植物功能性状间的相关性;用方差分析(one-way ANOVA)检测植物科类群、高度、生活型、花序、花色、果型、分布区对叶性状变异的影响;以一般线性模型( general linear model)进一步检测单因子作用的效应.统计分析使用 SPSS 19.0软件和Excel统计软件.
2 结果与分析
2.1 叶性状的变异特征
如图1所示,37种禾本植物SLA的平均值为14.22 m2/kg,SLA最大是陕甘花楸(Sorbuskoehneana)62.45 m2/kg,最小是冬瓜杨(Populuspurdomii,为2.16 m2/kg,变异系数是103.45%;LDMC平均为0.43 g/g,最大的是藏刺榛(Corylusferox),为1.11 g/g,最小的是华北落叶松(Larixgmelinii),为0.01 g/g,变异系数为61.61%;LNC平均为23.69 g/kg,最大的是牛奶子(Elaeagnusumbellate),为39.34 g/kg,最小的是岷江冷杉,为12.21 g/kg,变异系数是20.38%;LCC平均为458.94 g/kg,最大的是岷江冷杉(Abiesfargesii),为526.97 g/kg,最小是绣球(Hydrangeamacrophylla),为427.23 g/kg,变异系数是4.86%;LPC平均为36.30 g/kg,最大的是华北落叶松(Larixgmelinii),为52.19 g/kg,最小是陕甘花楸(Sorbuskoehneana),为21.40 g/kg,变异系数是20.76%(表2).37种植物的SLA和LDMC为单峰偏左分布,SLA集中在1~10 m2/kg,LDMC集中在0.25~0.5 g/g;LNC、LCC、LPC均为正态分布(图1).

图1 植物叶性状分布特征Figure 1 Distribution characteristics of plant leaf traits
2.2 性状间的相关性
Pearson相关分析结果显示,37种植物的LCC与植物高度呈极显著正相关关系(P<0.01),其线性关系为y=2.51x+438.93(表3和图2所示);LPC与LNC呈极显著正相关关系(P<0.001),其线性关系为y=0.852 2x+16.109(表3和图3所示);植物LCC与LDMC呈显著负相关(P<0.05),其线性关系为y= -27.915x+470.91(表3和图4所示);其他各性状之间无显著相关性(P>0.05).
2.3 叶性状变异与植物科类群、高度、生活型之间的关系
方差分析显示,不同科类群植物之间的LCC差异极显著(P<0.01),其间LNC差异显著(P<0.05),SLA、LDMC、LPC差异不显著(P>0.05).LCC最大为松科(502.84 g/kg),最小为禾本科(434 g/kg);LNC最大为胡颓子科(39.34 g/kg),最小为壳斗科(15.23 g/kg)(表4和图5).不同高度的植物LDMC差异显著(P<0.05),其间LCC差异极显著(P<0.01),SLA、LNC、LPC差异不显著(P>0.05).不同生活型的植物LNC和LPC差异极显著(P<0.01).不同花序类型植物之间的LDMC(P<0.01)和LCC(P<0.001)差异极显著.不同花色的植物LNC差异显著(P<0.05).不同果实类型的植物间SLA(P<0.001)和LCC(P<0.01)差异极显著,其间LDMC差异显著(P<0.05).植物的分布区对叶性状无显著影响(P>0.05,表4).
方差分析显示,科类群、植物高度和生活型分别解释SLA变异的42.4%、5.3%、0.3%,LDMC变异的50%、17.2%、9.4%,LNC变异的63.3%、3.3%、21.6%,LCC变异的74.1%、34.2%、27.7%,LPC变异的32.4%、5.1%、2%.花序、花色和果型分别解释SLA变异的39.8%、18%、83.3%,LDMC变异的45.9%、29.1%、43.8%,LNC变异的16.5%、42.6%、22.8%,LCC变异的60.8%、38.7%、54.4%,LPC变异的18.7%、31.5%、21.3%(表4).

表2 植物叶性状分布及其变异
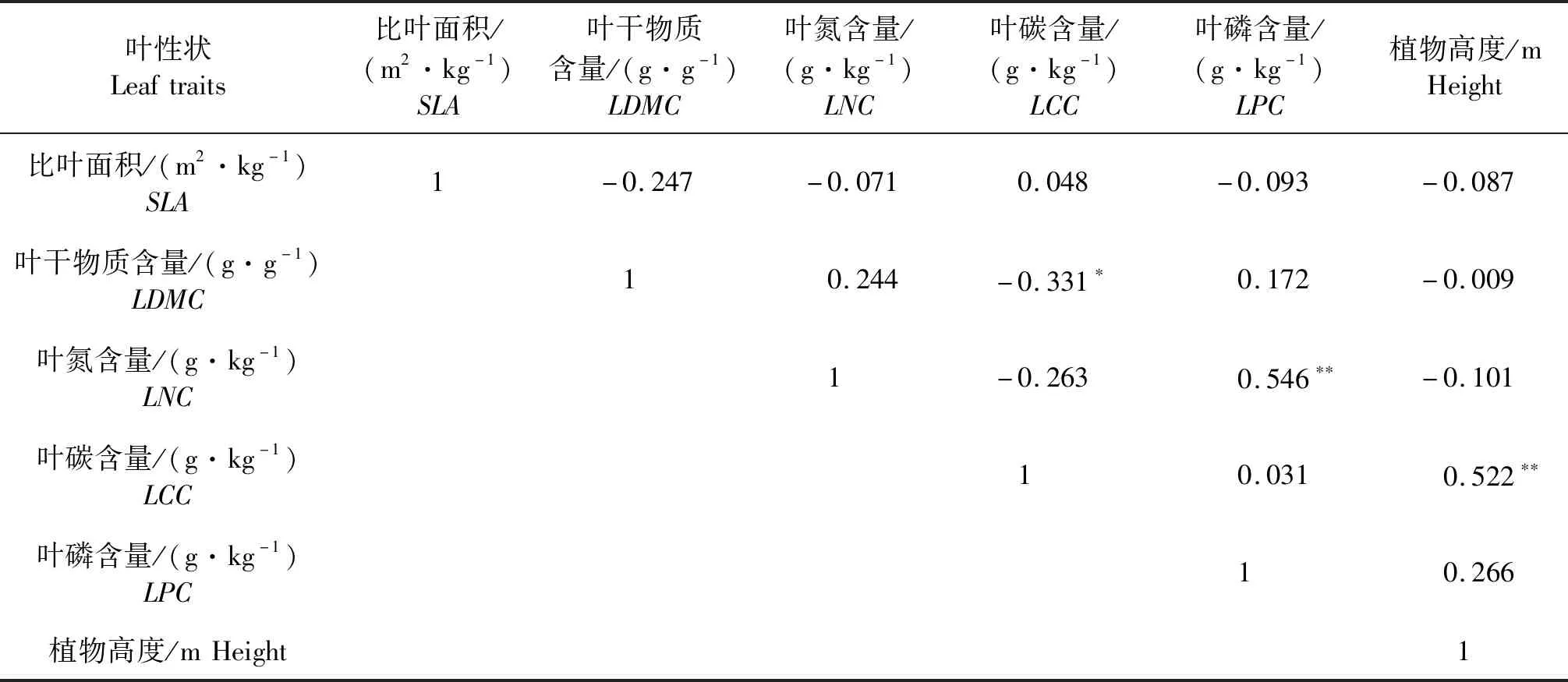
表3 植物功能性状间的Pearson相关性检验
**表示差异极显著(P<0.01);*表示差异显著(P<0.05).
** indicates the highly significance difference(P<0.01);* indicates significance difference(P<0.05).

表4 科类群、高度、生活型、花序、花色、果型及分布区对叶功能性状的影响
*** 表示P<0.001;**表示P<0.01;* 表示P<0.05;ns表示P>0.05.R2表示被每一个因子所解释的变异量.
*** indicatesP<0.001;** indicatesP<0.01,* indicatesP<0.05,ns:P>0.05.R2represents the variance explained by each factor.

图2 LCC与高度的关系Figure 2 The relationship between LCC and height

图3 LPC与LNC的关系Figure 3 The relationship between LPC and LNC

图4 LCC与LDMC之间的关系Figure 4 The relationship between LCC and LDMC
3 讨论
3.1 叶性状的变异特征
摩天岭北坡木本植物各叶性状属性值的变异程度存在很大的差异,反映了不同植物对环境的适应具有差异性.本研究发现,LCC属于弱变异(变异系数为4.86%),LDMC、LNC、LPC属于中等变异(变异系数为61.61%,20.38%、20.76%),而SLA属于强变异(变异系数为103.45%),进一步证实了群落内或相似生境的叶性状或生理特征存在种间差异[25].SLA的变异系数最大,这与何桂萍等[19]对木本植物叶性状的研究结果相一致,表明该研究区木本植物单位重量的叶面积变化较大,植物叶片投入的单位质量干物质所能截取光能的能力差异较大.碳是构成植物体内干物质的基本元素,LCC最小的是落叶植物绣球(Hydrangeamacrophylla),为427.23 g/kg,最大的是常绿植物岷江冷杉(Abiesfargesii),为526.97 g/kg,这说明不同生活型的物种叶片碳含量存在较大差异.37种植物的LDMC具有较低变异幅度,该结果同赵红洋等[26]对沙漠地带植物的研究结果相一致,说明LDMC是资源获取轴上较稳定的变量.

图5 不同科类群植物的LNC和LCCFigure 5 LNC and LCC of plants of different families and groups
本研究中植物的高度与LCC呈极显著正相关关系,C是植物体内的生命骨架元素和结构性物质[27],这说明有机物含量越高,C含量越高,植物生长越快.LNC与LPC呈极显著正相关关系(P<0.01),这与刘超等[28]对内蒙古灌木叶性状关系的研究,张奇平[29]对天目山主要树种叶性状的研究,以及张月强[30]对泰山木本植物功能性状间关系的研究结果一致,即叶片N与P呈显著正相关关系,这说明氮与磷元素之间存在协同作用.
3.2 叶性状的种间变异
植物性状的变异程度受生境过滤和生物竞争共同影响[31],生境过滤作用能够降低性状的变异程度,使得具有相似性状的物种在局域生境中共存,而这些共存物种会通过增加性状变异降低竞争强度.另有研究发现,植物类群和生活型对其性状分异的影响要大于环境因子如降雨、温度和经纬度[32].本研究通过对 37种植物功能性状在科类群和生活型上的变异分析发现叶氮含量和叶碳含量在科类群和生活型水平上有较大变异,科类群能解释63.3%的LNC变异和74.1%的LCC变异,生活型能解释21.6%的LNC变异和27.7%的LCC变异,说明该地带植物叶性状的种间变异在较大程度上受物种系统发育史的影响,这与陈文等[33]对粤东植物叶性状变异特征的研究结果相一致.生活型主要反映植物的资源获取和水分利用策略[34],因此不同生活型的植物的叶性状差异较大.
植物的营养性状和繁殖性状间也存在密切联系[25].植物需要通过控制生长、开花、传粉和结果的时间来合理分配资源[35].本研究显示,叶性状的种间变异与植物的高度、花序、花色和果型的关联度较大,与分布区无显著关联,其中高度能解释17.2%的LDMC变异和34.2%的LCC变异,花序能解释45%的LDMC变异和60.8%的LCC变异,花色能解释42.6%的LNC变异,果型能解释83.3%的SLA变异、43.8%的LDMC变异和54.4%的LCC变异,说明该地带植物叶性状的种间变异还受到植物生活史的影响,与植物个体在一生中所经历的生长发育的全过程紧密相关.
4 结论
植物功能性状能够响应生存环境的变化并对生态系统功能有一定影响[2],它反映了植物在表征生态系统功能方面的生态指示作用,强调其与生态系统过程和功能的关系[25],也反映了植物在特定环境中生长、繁殖和生存的能力.本研究结果显示:37种木本植物的植物功能性状在科类群和生活型上的变异分析发现,叶氮含量和叶碳含量在科类群和生活型水平上有较大变异,说明该地带植物叶性状的种间变异在较大程度上受物种系统发育史的影响.叶性状的种间变异与植物的高度、花序、花色和果型的关联度较大,与植物的分布区无显著关联,说明该地带植物叶性状的种间变异还受到植物生活史的影响.植物功能性状并不是孤立发挥作用的,植物在长期适应环境过程中,其形态和功能性状的特征不仅受环境的影响,也可能受植物生活史和物种系统发育史的影响.
