非核糖体肽合成酶基因腺苷酰化结构域序列克隆及分析
2019-03-26薛永常张成锁
薛永常, 张成锁, 李 根
(大连工业大学 生物工程学院,辽宁 大连 116034)
微生物在次级代谢过程中产生丰富的活性天然产物。除以核糖体途径,还能以非核糖体途径合成一系列低分子量的具药用价值的多肽类活性次级代谢产物[1],可被用于抗生素、免疫抑制剂、抗癌和抗病毒因子、铁载体及生物表面活性剂等[2],已成为近几年研究的热点[3]。在微生物中非核糖体肽类次生代谢产物的生物合成是由非核糖体肽合成酶(NRPS)催化的。NRPS属于模块化酶,各相互独立的模块按特定空间、顺序排列,在合成新生肽链的过程中具有不同的功能[4]。NRPSs至少含有腺苷酰化结构域(A)、肽酰基载体蛋白结构域(PCP)和缩合结构域(C) 等3个核心结构域。而A结构域负责合适的底物选择并活化[5], A结构域10个氨基酸残基呈口袋形状,为底物结合处,同时担任底物的特异性识别,是非核糖体多肽合成开始的起始处。NRPS基因常存在于合成次级代谢产物的基因簇中,通过克隆NRPS基因,利用探针或已有生物学软件能够对NRPS基因簇结构域进行分析、结构和功能预测,有助于寻找新型抗生素或生物活性物质[6-9],为相关新药的研发提供新靶点和新思路。本研究是在前期已从海洋链霉菌L1基因组DNA克隆了NRPS缩合结构域(即C结构域)基因片段[10]的基础上,开展NRPS腺苷酰化结构域的基因克隆,通过对所得基因序列及拟翻译氨基酸序列的组成成分、理化性质、结构特征等进行预测和分析,为后续研究A结构域特异性和NRPS生物合成基因簇提供了理论依据,也为寻找新基因簇提供参考。
1 材料与方法
1.1 材料
1.1.1 菌种来源StreptomycesglobisporusL1为本实验室分离鉴定并保存;大肠埃希菌(Escherichiacoli) DH5 为大连工业大学分子实验室保存菌株。
1.1.2 培养基 ①高氏一号培养基:可溶性淀粉20 g,NaCl 0.5 g,K2HPO4·3H2O 0.5 g,KNO31.0 g,MgSO4·7H2O 0.5 g,FeSO4·7H2O 0.01 g,陈海水定容至1 L,重铬酸钾0.025 g,琼脂20 g,pH 7.4~7.6,121 ℃灭菌20 min。②种子液培养基:葡萄糖10 g,蛋白胨1.2 g,酵母提取物2 g,陈海水定容至1 L,pH 7.2,121 ℃灭菌20 min。
1.1.3 试剂与仪器TaqDNA聚合酶、DNA Marker、2×GC-rich PCR MasterMix、细菌基因组DNA提取试剂盒,普通琼脂糖凝胶DNA回收试剂盒均购自天根生化科技有限公司;限制性内切酶Hind Ⅲ和BamH I、pMD 19-T载体购自宝生物工程(大连)有限公司;引物合成及序列测定由北京六合华大基因科技有限公司完成。TaKaRa TP 600 PCR仪,日本TaKaRa公司;Chemi System UVP Bio-imaging System凝胶成像系统, 美国UVP公司; Himac CR-21G高速冰冻离心机, 日本日立公司。
1.2 方法
1.2.1 NRPS基因引物设计和合成 参考Gontang等[11]发布NRPS通用引物设计了用于克隆A结构域基因片段的带有酶切位点的引物:A2:5′-CGGGATCCTATCTACACCTCGGGATC-3′,下划线部分为添加的BamH I酶切位点序列;A4:5′-CCAAGCTTGACGTCCCCCGTCCGGTAC-3′,下划线部分为添加的Hind Ⅲ酶切位点序列。
1.2.2 NRPS基因克隆及测序 以细菌基因组DNA提取试剂盒提取的L1菌株基因组DNA为模板,以A2、A4为特异引物进行目的片段扩增,通过梯度PCR确定最适退火温度。PCR扩增体系为2×GC-rich PCR MasterMix 10 μL,上下游引物各1 μL,模板DNA 1 μL,ddH2O补足20 μL。PCR扩增程序:94 ℃预变性3 min;94 ℃变性35 s,63 ℃退火35 s,72 ℃延伸45 s,扩增反应30个循环;72 ℃延伸10 min,4 ℃保存。扩增产物经1%琼脂糖凝胶电泳检测,回收目的片段与pMD19-T连接,转化DH5 感受态细胞,阳性单克隆进行特异引物PCR和质粒双酶切检测。重组质粒送北京六合华大基因科技有限公司测序。
1.2.3 目的基因生物信息学分析 将测序的扩增序列进行BLAST搜索,确定扩增序列正确性。利用ORF Finder进行阅读框查找与翻译。利用BioEdit和ExPASy ProtParam在线工具预测其理化性质。SOPMA数据库对拟翻译氨基酸序列进行二级结构组成特点分析。利用Swiss-Model工具进行同源建模,分析其三维模拟结构。
2 结果与分析
2.1 NRPS基因克隆及核苷酸序列分析
以L1基因组DNA为模板,以A2、A4为特异引物进行PCR的退火温度优化。当退火温度为63 ℃时能够扩增出一条750 bp左右的单一清晰条带,与预期大小一致,因此确定63 ℃为扩增的最适退火温度。回收目的片段与pMD19-T连接,转化大肠埃希菌DH5 感受态细胞。提取重组质粒,进行特异引物的PCR,在大约750 bp左右能够扩增出一条清晰整齐条带(图1)。
对重组质粒进行Hind III和BamH I双酶切,同样能够得到约750 bp目的片段(图2),酶切片段与克隆目的片段大小相近,证明目的片段插入了pMD19-T克隆载体中。从测序结果得知,插入片段大小为715 bp,反向插入到pMD19-T载体中。在线BLAST比对表明,其与已发布的StreptomycesglobisporusC-1027的NRPS基因序列相似度达93%,证明扩增的基因片段属于NRPS基因的部分序列。

图1 NRPS基因PCR扩增电泳Fig.1 Agarose gel electrophoresis of amplified NRPS gene1:DNA Marker IV;2:扩增片段1: DNA Marker IV;2: amplified fragment from genomic DNA

图2 NRPS重组质粒双酶切电泳图Fig.2 Agarose gel electrophoresis of the recombinant plasmid cut by Hind III and BamH I1: DNA Marker IV; 2: 重组质粒; 3: Hind III和BamH I双酶切质粒1: DNA Marker IV; 2: recombinant plasmid; 3: the recombinant plasmid cut by Hind III and BamH I
2.2 NRPS基因编码氨基酸序列分析
利用ORF Finder对扩增序列进行开放阅读框分析,拟翻译氨基酸序列为IYTSGSTGKPKGV-VTEYAGLTNMLINHQRRIFEPVLAEHGHRTFRIA-HTVSFAFDMSWEELLWLADGHEVHICDEELRRD-APRLVDYCLRHGIDVINVTPTYAQQLVAEGLLED-PERRPALVLLGGEAVTPTLWQRLAETEGTVGYN-LYGPTEYTINTLGVGTFECQDPVVGVAIDNTEVY-VLDPWLRPLPDGVPGELYVSGIGIARGYLGRSAQ-TAHRFVACPFGAPGERMYRTGDV
该序列含有238个氨基酸,属于NRPS腺苷酰化结构域序列的一部分,将此片段命名为A1。利用Blast比对发现,该氨基酸序列中含有一个A结构域特有的AFD_class_I superfamily核心结合区,此家族包括酰基-、酰基-CoA连接酶、非核糖体肽合成酶腺苷酰化结构域和萤火虫荧光素酶(图3),在非核糖体肽合成酶腺苷酰化结构域中,催化ATP依赖反应。
选取与A1同源性较高的9个氨基酸序列构建NJ进化树,结果见图4。结果显示A1与Streptomycessp. EN27含有的NRPS处于同一分支,证明该序列为NRPS基因序列。
将该拟翻译氨基酸序列与Streptomycessp. EN27的非核糖体肽合成酶进行序列比对,同源性达到99%以上,证明扩增序列的翻译产物即为NRPS(图5)。
利用在线网站PKS/NRPS Analysis Web-site (http://nrps.igs.umaryland.edu/)分析得出,该氨基酸片段A1属于NRPS的A结构域,这与在NCBI中Blast结果相一致,说明A1片段确实属于NRPS的A结构域(图6)。
通过模拟在其绑定位点中心区域,发现有8个氨基酸残基发挥催化作用,分别为DMWNLGLI。利用NRPS Predictive Blast工具(http://nrps.igs.umaryland.edu/blast.html)搜索具有该活性位点的NRPS,发现A1与含有SyrB-M1-Thr活性位点的NRPS相似度达到62%,说明扩增的基因片段为NRPS的A结构域部分序列。

图3 扩增序列拟翻译氨基酸结构域分析Fig.3 The putative amino acid domain analysis from amplified sequences

图4 A结构域进化树构建Fig.4 Construction of Adenylation domain evolutionary tree

图5 A1拟翻译氨基酸序列与Streptomyces sp. EN27 NRPS的比对Fig.5 The alignment of A1 amino acid sequence with Streptomyces sp. EN27 NRPS
2.3 跨膜结构域的预测和分析
跨膜结构域一般由20个左右的疏水氨基酸残基构成,在膜中形成α-螺旋,其外部疏水侧链通过范德华力与脂双层分子脂肪酸链相互作用,是膜中蛋白与膜脂相结合的主要部位。对跨膜结构域的预测和分析,对于推断其在细胞中的作用部位具有重要意义。本研究应用在线工具TMPred对A1进行跨膜结构域分析(图7),推断出A1不存在跨膜结构域。
2.4 疏水性亲水性的预测和分析
采用Bioedit软件预测NRPS基因编码氨基酸序列理化性质,得出该片段为亲水性蛋白质(图8)。
根据ExPASy ProtParam在线分析得出:A1的氨基酸组成个数为238,理论分子量为26.36 kDa,等电点pI为5.06,带负电荷的氨基酸残基数(Asp+Glu) 31,带正点荷的氨基酸残基数(Arg+Lys) 19。在280 nm波长下,测得其水溶液消光系数为38 640 M-1cm-1,0.1%浓度的Abs为1.465。当成熟肽N端为Ile时,半衰期在酵母体内约为30 min,在大肠埃希菌体内大于10 h。该蛋白不稳定系数为42.43,脂肪指数91.30;平均亲水性系数(GRVAY)为-0.132。综上,A1的不稳定指数达到42.43,大于阈值40,表明A1为不稳定蛋白。其负电荷氨基酸残基数较多,故判断为酸性蛋白质。根据GRVAY数值-0.132推测A1为亲水性蛋白质,这与BioEdit软件预测结构一致。

图8 A1亲水性/疏水性分析Fig.8 Hydrophilicity/Hydrophobicity analysis results of A1
2.5 二级结构的预测与分析
蛋白质的二级结构是多肽链中相邻多个氨基酸残基形成的局部肽链空间,包括α-螺旋、β-转角、β-折叠片、延伸链及无规则卷曲等二级结构元件组成。
A1氨基酸序列的SOPMA二级结构预测显示其α-螺旋占26.89%,β-转角占10.50%,无规则卷曲占31.51%,伸展链占31.09%。可知α-螺旋和无规则卷曲是A1二级结构的主要元件,而β-转角和伸展链则散布于整个蛋白质中(图9)。

图9 A1二级结构预测Fig.9 The secondary structure prediction results of A1
2.6 三级结构的预测与分析
应用SWISS-MODEL在线对其三级结构进行同源建模(图10)。A1三维空间结构中α-螺旋、β-转角和无规则卷曲与二级结构预测基本一致。其中5t3d.1.A模板与靶蛋白的相似性评估得分为35.22%,靶蛋白三维模型与数据库中Enterobactin synthase component F的结构相似。GMQE分值为0.71,QMEAN分值为-3.04,说明靶蛋白模型与模板拟合度较高,靶蛋白模型可靠。靶蛋白与模板氨基酸的对应序列(图11)。初步判断扩增的腺苷酰化结构域片段与Enterobactin synthase component F同源。为今后研究A1特异性与功能提供了重要参考,也为获取相应NRPS基因全长提供了实验依据。
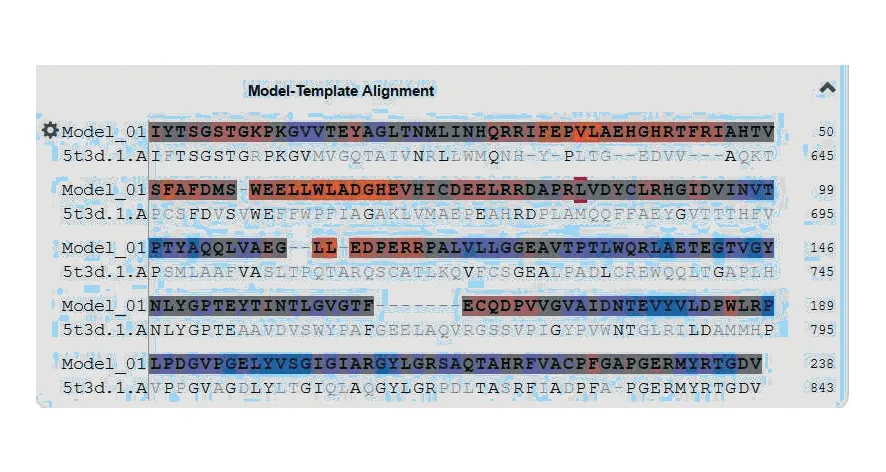
图10 A1氨基酸片段3D结构预测Fig.10 3D structure prediction of A1 amino acid fragment

图11 A1三维建模序列Fig.11 The 3D modeling sequence of A1
3 讨 论
A结构域对于整个NRPS是不可或缺的部分,在底物激活期间,A结构域的小亚结构域靠近大亚结构域,形成闭合状态[12],旋转140°为PCP硫醇化作用暴露出活性位点[13]。经1/3催化周期完成后,A结构域由闭合状态转变为开放状态,此时允许底物进行绑定[14]。Miller等[15]证明PCP结构域与A结构域相连区域,保守基序LPxP与A小亚结构域形成稳定的相互作用,从而缩短了连接区域的有效长度。构象改变驱使A结构域运动到PCP结构域。在PCP结构域的存在下,A结构域催化效率大大提高[16-17]。而A结构域在催化同时进一步引导PCP结构域移动到A和C结构域活性位点上。这个周期运动由A结构域更迭周期所调节。有证据表明在一个完整的NRPS模块中,A结构域能够催化2个活化周期,其中一个加载到PCP结构域,另一个用于激活下一个氨基酸。当上游缺少供体底物时,催化即刻停止[14]。结合观察A结构域状态变化,如果非核糖体肽在C结构域供体位点不可用,PCP结构域与上游C结构域相互作用的同时,会“冻结”A结构域的腺苷酰化作用。该机制有很重要的调节意义,因为被绑定的底物从PCP结构域释放之前,下一周期PCP是不可能成为巯基化状态的[18]。鉴于A结构域所具有的保守结构,克隆NRPS的A结构域序列对于探测NRPS基因的存在,并根据A结构域序列信息预测肽骨架的结构具有重要意义。本研究克隆了非核糖体肽合成酶A结构域基因,为研究NRPS全序列乃至整个基因簇功能结构提供参考。
