大肠杆菌合成琥珀酸的代谢工程研究进展
2018-11-05白家齐韩北忠陈晶瑜
白家齐,孟 娇,韩北忠,陈晶瑜*
(中国农业大学 食品科学与营养工程学院,北京 100083)
琥珀酸(succinic acid)学名丁二酸,属于四碳二羧酸家族的一员,广泛地存在于动物、植物和微生物中[1]。琥珀酸及其衍生物被广泛地应用于食品,制药和化妆品等领域。琥珀酸的发酵潜力早在1980年就得到认可,在美国能源部公布的12种有潜力可大宗生产的化合物中,琥珀酸居于第一位[2],目前生产琥珀酸的方法主要有化学法和生物法,其中化学法仍然是琥珀酸生产的主要方法[3]。但是化学法生产的过程中污染大、成本高,不符合如今倡导的可持续发展原则,生物发酵法生产琥珀酸因具有资源可再生及吸收CO2等优点而拥有广阔的发展潜力。
琥珀酸盐作为三羧酸循环(tricarboxylicacidcycle,TCA)中的中间体之一,可以由许多微生物合成:产琥珀酸放线杆菌(Actinobacillus succinogenes)[4],产琥珀酸厌氧螺菌(Anaerobiospirillum succiniproducens)[5],谷氨酸棒状杆菌(Corynebacterium glutamicum)[6],酿酒酵母(Saccharomyces cerevisiae)[7],大肠杆菌(Escherichia coli)等,由于生长速度快,基因信息清楚,基因操作简单,营养需要量极小,大肠杆菌已被广泛研究用于琥珀酸的生产[8]。
为了提高大肠杆菌产琥珀酸的能力,研究人员已经探索了多种代谢工程策略[9-11],如激活糖酵解途径(embden meyerhof pathway,EMP)、过表达丙酮酸代谢酶、消除竞争途径、并提供还原当量和能量等[12-15]。本文介绍代谢控制发酵产琥珀酸方面的研究进展,以期对今后的发展提供新思路。
1 消除竞争途径
1.1 敲除竞争性副产物
如图1所不,在厌氧条件下,大肠杆菌进行混合酸的发酵,生成大量的乙酸、乳酸、甲酸和乙醇,乃积累少量的琥珀酸。这些副产物的生物合成不仅消耗碳源,而且还消耗限制琥珀酸生产的烟酰胺腺嘌吟二核苷酸(nicotinamide adeninedinucleotide,NAD+)。因此,大肠杆菌的代谢工程必须关注阻断副产物生物合成途径[6]。JANTAMA K等[16]以KJ091为出发菌株敲除tdcD(编码苏氨酸脱羧酶)和tdcE(编码2-酮丁酸甲酸裂解酶)后,乙酸的含量减少了50%,琥珀酸的得率达到1.3 mol/mol。在此基础上,接着敲除aspC(编码天冬氨酸转氨酶)和sfcA(编码NAD+依赖的苹果酸酶),琥珀酸的产量和得率分别提高至700 mmol/L和1.5 mol/mol。
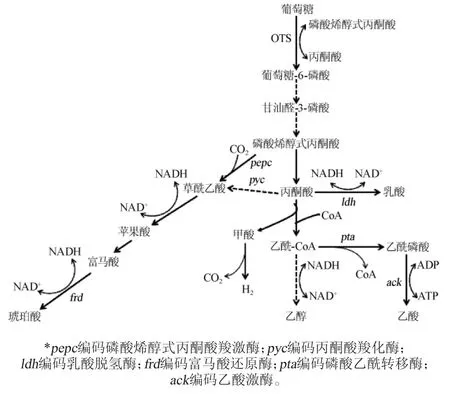
图1 厌氧条件下大肠杆菌生物合成琥珀酸的代谢途径Fig.1 Metabolic pathways of succinate biosynthesis in E.coli under anaerobic conditions
1.2 失活磷酸烯醇式丙酮酸转运系统
磷酸烯醇式丙酮酸(phosphoenolpyruvate,PEP)是合成许多工业化学物(如琥珀酸,苹果酸和芳香族化合物)的重要前体物。然而在大肠杆菌中,葡萄糖主要是通过磷酸烯醇式丙酮酸转运系统(phosphoenolpyruvate transport system,PTS)运输到细胞质中,其中有一半的PEP被用于葡萄糖的摄取和磷酸化。失活PTS系统,提高PEP前体物的供给,是提高代谢流通向目标产品琥珀酸的重要代谢工程策略。然而,PTS突变的菌株在生长和糖耗方面表现得非常缓慢,这样的发酵特点不适用于大规模的工业化生产。ptsG基因突变的菌株PB11通过代谢进化后提高了糖耗的速率,研究发现半乳糖透性酶-葡萄糖激酶(galactose permease-glucokinase,Galp-Glk)转运系统成为了该菌株葡萄糖的摄取和磷酸化的主要途径[17]。TANG J等[18]在菌株XZ-T108(ΔptsI ΔldhAΔpflB,Pck)中组合调控大肠杆菌自身的galp(编码半乳糖透性酶)和glk(编码葡萄糖激酶)后,葡萄糖的利用率得到了提高,琥珀酸的产量从156mmol/L提高至187mmol/L,生产速率从0.19 g/(L·h)提高至0.22 g/(L·h)。
2 过表达自身或外源的关键酶
2.1 过表达内源羧化酶
在PEP下游有两种代谢途径:通过固定一个分子的CO2,PEP(C3)可以转化为草酰乙酸(oxaloacetate,OAA)(C4),并且草酰乙酸进一步转化为琥珀酸;PEP也可用于生产丙酮酸盐,导致副产物乙酸盐,甲酸盐,乙醇和乳酸盐的形成。因此,C4化合物的合成对琥珀酸生产是重要的。大肠杆菌自身有4种羧化酶:磷酸烯醇式丙酮酸羧化酶(phosphoenolpyruvate carboxylase,PPC)、磷酸烯醇式丙酮酸羧激酶(phosphoenolpyruvate carboxykinase,PCK)和两种苹果酸酶(malic enzyme,Mae)A和B。MILLARDCS等[19]在菌株JCL1208的基础上,采用诱导型质粒过表达大肠杆菌自身的PPC,PPC酶活力提高了8.57倍,琥珀酸的转化率和产量分别提高了3.75和3.50倍;相反,过表达大肠杆菌自身的PCK则对琥珀酸产量没有明显的提高。STOLSL等[20]在NZN111菌株中过表达大肠杆菌本身的MaeA,琥珀酸的产量较出发菌提高了6倍。在菌株K-12中过表达自身MaeB,使得重组大肠杆菌的琥珀酸产量提高至15.4mmol/L,比出发菌株增强了1.4倍[21]。TAN Z等[22]通过组合优化PPC和PCK两个酶的表达提高了琥珀酸的产量。研究发现,PCK的活力和琥珀酸的产量有正相关性,而PPC乃有在特定的酶活力范围内PPC活力和琥珀酸的合成呈正相关性,超过这个界定范围,细胞的生长和琥珀酸的形成就会显著下降。通过对PPC和PCK的组合优化,最优菌株琥珀酸的产量提高了近70倍,琥珀酸的得率达到了1.12 mol/mol。
2.2 过表达外源羧化酶
现有的文献报道中,被研究最多的外源羧化酶就是丙酮酸羧化酶(pyruvate carboxylase,PYC)和磷酸烯醇式丙酮酸羧激酶(phosphoenolpyruvate carboxykinase,PCK)。虽然在大肠杆菌中PCK催化的反应主要是糖异生,但是在一些高产琥珀酸的瘤胃细菌里PCK却是主要的羧化酶。厌氧条件下,葡萄糖经EMP途径再经TCA还原臂生成琥珀酸的途径中是没有净ATP产生的,为了解决厌氧条件下菌体的维持能,引入ATP依赖性的磷酸烯醇式丙酮酸羧激酶(PCK)成为大肠杆菌主要的羧化酶是一个重要的代谢工程策略。KIM P等[23]在大肠杆菌K-12中过表达来自产琥珀酸放线杆菌的PCK后,琥珀酸的产量并没有明显的变化,这可能是由于大肠杆菌自身PPC的高活力所导致的。然而,在PPC突变的菌株中过表达产琥珀酸放线杆菌的PCK后,琥珀酸的产量较出发菌株提高了6.5倍。
3 提高NADH的供给
在厌氧的条件下,一分子葡萄糖经过EMP途径能产生两分子的还原型辅酶Ⅰ(nicotinamideadeninedinucleotide,NADH)和两分子磷酸烯醇式丙酮酸(PEP),然而经TCA还原臂生成一分子琥珀酸则需要一分子的PEP和两分子的NADH,所以葡萄糖经这条途径的理论得率乃有1 mol/mol葡萄糖,为了提高NADH的供给,目前以甘油为底物产琥珀酸、激活乙醛酸途径和表达外源的甲酸脱氢酶已经得到了应用。
3.1 利用甘油为底物产琥珀酸
甘油是生物柴油生产的主要副产品。随着生物柴油产业的发展,如何将甘油转化为有价值的产品已成为亟待解决的问题。作为糖异生碳源之一,甘油具有高还原性,因此有利于产生琥珀酸等还原产物。其天然pck基因在双重突变体大肠杆菌菌株MLB(ΔldhA和ΔpflB)中的过表达显著提高了甘油利用率和琥珀酸产量。与对照相比,甘油消耗量从2.2 g/L增加至35.8 g/L,琥珀酸产量从2.5 g/L增加至42.5 g/L[8]。BLANKSCHIEN M D等[24]通过代谢工程使得大肠杆菌在微氧下利用甘油产琥珀酸。首先失活竞争途径的几个关键基因ldhA、adhE和pflB,随后引入来自乳酸菌的丙酮酸羧化酶(PYC),琥珀酸的产率和得率分别达到了400 mg/(g·h)和0.69 g/g甘油。ZHANGX等[25]在已构建好的产琥珀酸的菌株上,通过失活pflB和ptsI基因,强化pck基因的表达,重组后的菌株利用甘油为底物产琥珀酸,琥珀酸的得率达到了理论最大得率的80%。
3.2 激活乙醛酸途径
在大肠杆菌的代谢途径中,有两条路径能形成琥珀酸。一条是在完全厌氧的条件下较为活跃的TCA还原臂,另一条则是在好氧条件下较为活跃的乙醛酸支路。乙醛酸循环催化一分子的草酰乙酸和两分子的乙酰辅酶A形成一分子的琥珀酸和一分子的苹果酸,生成的这一分子苹果酸再与一分子的NADH形成一分子的琥珀酸,总得来说,消耗一分子的NADH能形成两分子的琥珀酸。研究表明,当71.4%的碳流量通向TCA还原臂,28.6%的碳流量通向乙醛酸支路,琥珀酸得率能达到最大理论得率1.714 mol/mol[26]。研究发现,在厌氧条件下敲除乙酸合成基因ackA和pta对激活乙醛酸途径起到了关键的作用。敲除ldhA、ptsG和ackA-pta基因,重组菌株YJ003琥珀酸的产量能达到150.78 mmol/L,相比对照菌株提高了5倍[27]。
3.3 表达外源的甲酸脱氢酶
甲酸脱氢酶能催化一分子的甲酸形成一分子的CO2和一分子的NADH,引入外源的甲酸脱氢酶不仅能为琥珀酸的合成提供额外的还原力,加强琥珀酸的合成能力,同时还能降低副产物甲酸的积累,有助于下游产品的分离纯化。BALZERGJ等[28]将假丝酵母中NAD+依赖型的甲酸脱氢酶基因fdh1引入大肠杆菌SBS550MG中,重组后的菌株琥珀酸的产率从1.4 g/(L·h)提高至2 g/(L·h),甲酸从17mmol/L减少至0~3 mmol/L。LITSANOV B等[29]将来自分支杆菌的甲酸脱氢酶基因fdh引入产琥珀酸的谷氨酸棒杆菌中,重组菌株BOL-3/pAN6-gap在以葡萄糖为单一碳源的培养基中发酵培养,琥珀酸的得率达到1.07 mol/mol,当发酵液中添加一定量的甲酸后,琥珀酸的得率提高到1.41 mol/mol。
4 扰动磷酸戊糖途径(PPP途径)
磷酸戊糖途径(pentosephosphatepathway,PPP)途径产生的还原力是烟酰胺腺嘌吟二核苷酸磷酸(nicotinamide adeninedinucleotidephosphate,NADPH),而琥珀酸的合成则需要烟酰胺腺嘌吟二核苷酸(NADH),通过扰动PPP途径来提高琥珀酸的生物合成还需要体内转氢酶的参与,进而实现辅因子NADPH和NADH的相互转化。在大肠杆菌中存在两种类型的转氢酶,它们分别是能量依赖型的膜定位嘧啶核苷酸转氢酶(PntAB)和非能量依赖型的可溶性转氢酶(UdhA)。当胞内NADPH的浓度较低时,PntAB催化NADH和NADP+生成NADPH和NAD+,反之,当胞内NADPH的浓度较高时,UdhA则催化NADPH和NAD+生成NADH和NADP+[30]。MENG J等[14]将参与谷氨酸棒状杆菌产生NADPH的解调基因zwf243(编码葡萄糖-6-磷酸脱氢酶)和gnd361(编码6-磷酸葡萄糖酸脱氢酶)导入大肠杆菌以产生琥珀酸,有益于突变脱氢酶的共表达,其去除了PPP途径氧化部分中的反馈抑制,将琥珀酸产量从1.01 mol/mol葡萄糖增加至1.16 mol/mol葡萄糖。然后过表达了3个关键基因,pgl(编码6-磷酸葡萄糖酸内酯酶)、tktA(编码转酮酶)和talB(编码转醛酶)以重定向更多的碳流向PPP途径,并进一步将琥珀酸得率提高至1.21 mol/mol。
5 强化琥珀酸的输出
产物的输出对于提高目标产品的产量和得率是非常重要的。在厌氧条件下,E.coli四碳二羧酸的摄取,运输和输出主要是依靠体内的Dcu转运系统。Dcu转运系统主要包含4种运输蛋白,它们分别是DcuA、DcuB、DcuC、DcuD。DcuA和DcuB是两个同源蛋白,它们能够独立地或相互地作用大肠杆菌体内冗余的四碳二羧酸的输出,研究表明DcuB运输蛋白的主要功能是催化四碳二羧酸的交换,例如催化延胡索酸转变为琥珀酸[31],然而DcuA确切的生理功能至今还未见报道。尽管DcuC蛋白能协助DcuA和DcuB蛋白去完成延胡索酸的摄取和延胡索酸与琥珀酸的交换,但是已有报道指出DcuC蛋白的主要功能却是协助在厌氧条件下以葡萄糖为碳源时琥珀酸的胞内到胞外的输出[32]。DcuD虽然是DcuC的同源蛋白,但是DcuD在体内大多情况下是不被表达的,所以DcuD的主要生理功能至今也不清楚[33]。CHEN J等[34]以大肠杆菌Suc-T110为出发菌株,首先分别敲除编码Dcu运输蛋白的基因,dcuA、dcuB、dcuC、dcuD,结果发现敲除dcuA和dcuD基因后,琥珀酸的产量和得率几乎没有变化。敲除dcuB和dcuC基因后,琥珀酸的产量分别下降了15%和11%。然而,同时敲除dcuB和dcuC这两个基因后,琥珀酸的产量下降了90%。该研究表明当以葡萄糖为单一碳源时,DcuB和DcuC是大肠杆菌体内琥珀酸胞内到胞外输出的两个关键蛋白。随后,对DcuB和DcuC这两个运输蛋白的RBS进行组合优化,琥珀酸的产量从273 mmol/L提高至366 mmol/L,琥珀酸的得率从1.12 mol/mol提高至1.41 mol/mol。
6 代谢进化
代谢进化作为一种有价值的方法已经被广泛用于菌株发育和优化,这不仅可以导致潜在途径的激活,而且还会导致突变株的期望表型和环境适应性的改善[35]。JANTAMA K等[36]在野生型的E.coli ATCC 8739的基础上,结合代谢工程和代谢进化,筛选到琥珀酸生产最优的两株菌KJ060(ΔldhAΔadhEΔackAΔfocA ΔpflB)和KJ073(ΔldhAΔadhE ΔackAΔfocAΔpflBΔmgsAΔpoxB),在以葡萄糖为单一碳源时,琥珀酸的产量达到622~733 mmol/L,得率达到1.2~1.6 mol/mol。ZHU X等[37]通过代谢进化得到一株高产琥珀酸的菌株HX024,在以基本盐为培养基时该菌株琥珀酸的产量达到了813 mmol/L,琥珀酸的得率达到1.36 mol/mol。通过全基因组测序,转录组学及酶学的分析,lpdA基因中有三个核苷酸的突变,该基因的突变解除了丙酮酸脱氢酶对NADH的抑制作用,丙酮酸脱氢酶的活力提高,提供更多NADH用于琥珀酸的合成。除此之外,磷酸戊糖途径中的转酮酶(TktA)以及转氢酶(SthA)的活力也得到了相应的提高。结合逆向代谢工程,在野生型的大肠杆菌中过表达磷酸戊糖途径中的tktA及sthA,琥珀酸的得率从1.12mol/mol提高到1.33 mol/mol。
7 展望
控制微生物代谢发酵来生产琥珀酸有着不可替代的优势,另外产品更加适用于食品、医药、农药等与人类健康密切相关的行业,具有巨大的生产潜力,目前为止通过多种基因策略来提高琥珀酸生物合成的尝试是成功的,但是仍然达不到工业化大生产所需要的效率和强度,为了能够实现琥珀酸生物合成工业化,依然需要深入研究大肠杆菌琥珀酸代谢途径、过表达关键酶、强化输出、代谢进化等,综合利用强化琥珀酸合成途径来构建新的产琥珀酸途径,开发出具有产业化前景的产琥珀酸基因工程菌株,同时随着系统生物学、分子生物学技术等的不断发展,琥珀酸的生物合成定会充分体现出环境和经济效益,也将推动发酵产琥珀酸的产业化,生产出更具优势的琥珀酸产品。
