基于外泌蛋白质组的酿酒酵母信号肽元件的分析及鉴定
2018-08-15田雷瑜曹筠嵩刘忞之杨燕王伟
田雷瑜,曹筠嵩,刘忞之,杨燕,王伟
药用蛋白和工业生物催化剂通常天然含量很低,需要异源重组和表达制备,这需要发展一个稳定的、易操作的和纯化工艺可放大的高效生物表达系统。酿酒酵母细胞相对于大肠杆菌和哺乳动物具有高效的胞外分泌特点和易于遗传操作的整合型或附加体型的表达载体,其已被广泛用于重组蛋白的生物表达系统[1-2]。
酵母细胞自身外泌并转运到细胞表面或发酵液中的蛋白可分为两大类,一类蛋白是经过经典的内质网-高尔基体途径(ER-Golgi)分泌的,这类外泌蛋白的前体通常含有信号肽,翻译后首先被定向地转运到内质网;另一类不含有信号肽的胞质蛋白如糖代谢酶、伴侣蛋白和翻译起始因子等也可以经过非经典的 ER-Golgi 非依赖分泌途径分泌到胞外,其外泌的分子机制还没有阐明[3]。许多外泌蛋白和胞内膜蛋白在合成时都依赖于其 N-端的信号肽功能元件,细胞内膜定位的蛋白尽管也经过ER-Golgi 途径转运,但是其信号肽的下游和分子内部存在穿膜肽或 C-膜定位信号(如核定位信号肽)控制蛋白的亚细胞转位[4]。分泌型的信号肽含有三个区域:N-、H- 和C- 区;N- 区含有碱性正电荷的极性氨基酸残基,H- 区含有 7 ~ 15 个疏水氨基酸残基(如丙氨酸、亮氨酸、异亮氨酸和苯丙氨酸)形成的 α-螺旋,跨越 ER 膜,促进新生肽的转位,它是信号肽的主要功能区;分泌肽的 C-区含有极性结构,作为信号肽酶 I 的识别剪切位点,其中小的和中性氨基酸残基(如丙氨酸和甘氨酸)位于剪切位点的 -3和 -1[5-6]。到目前为止,无论是在酿酒酵母还是在甲醇酵母的蛋白质工程研究中所使用最多的信号肽是酿酒酵母的交配型 α-因子的信号肽功能元件(85-aa prepro 区)[1],其次是酿酒酵母的蔗糖水解酶的信号肽[7]。α-因子的信号肽功能元件不仅含有一个靶向内质网且由信号肽酶剪切的19 个氨基酸残基信号肽,还含有一个 66 个氨基酸残基的前导区[8-9]。因此由 α-因子引导的异源蛋白分泌表达在内质网腔内由信号肽酶剪切后,从内质网出芽并转运进入高尔基体;然后在高尔基体腔内再由内肽酶 Kex2 剪切 2 个保守的碱性氨基酸残基(Lys-Arg 或 Arg-Arg)的羧基端肽键释放前导区,最后再包装成囊泡转运到细胞表面。因此,在蛋白的分泌过程中,如果没有前导区的作用,可能会导致融合蛋白积聚在内质网;而其他信号肽(仅含有 pre 区)与 α-因子的前导区融合形成具有 α-因子野生型相似的功能元件,促进融合蛋白的外泌。可见 α-因子的前导区在由内质网到高尔基体的转位过程中起到非常重要的作用[8]。
目前,除 α-因子信号肽外,还有很多研究报道了酿酒酵母和甲醇酵母自身分泌蛋白的信号肽应用于异源蛋白的生物表达,如具有 α-因子信号肽相似结构的酿酒酵母 PIR 家族的细胞壁蛋白 CIS3和 HSP150 等多个信号肽用于人白细胞介素-2、脂肪酶、小鼠的唾液酸转移酶等多个蛋白的外泌表达[10]。Massahi 和 Çalık[11]系统地研究了甲醇酵母5 个信号肽对人生长激素表达的影响;此外,还有研究报道了异源的信号肽如菊粉酶的信号肽引导人皮质素蛋白在酿酒酵母中的分泌表达[12]。但综合分析这些研究结果,很容易地发现酿酒酵母的α-因子信号肽依然是促进融合蛋白分泌表达的活性较好的功能元件[1]。为进一步从酿酒酵母中研究发现具有更好促分泌活性的信号肽功能元件,本研究基于酿酒外泌蛋白质组的研究结果,采用生物信息学手段进行计算分析,确定了 16 个定位于细胞壁的外泌蛋白信号肽元件,选取其中编码外切β-(1,3)-葡聚糖酶 1(exo-1,3-beta-glucanase 1,EXG1)的信号肽元件为研究目标,以 α-因子信号肽的促分泌活性作对比分析,结果表明 EXG1 的信号肽功能元件具有较好的促分泌活性。

表 1 本研究使用的 PCR 扩增引物Table 1 PCR primers used in this study
1 材料与方法
1.1 材料
大肠杆菌Escherichia coliTrans1-T1 购自北京全式金生物技术有限公司,用于质粒载体的构建;本实验室保存的酿酒酵母 W303-1b/ΔES(E表示EXG1 基因敲除,S表示SPR1基因敲除)[13]为宿主进行酵母的转化和融合蛋白的外泌表达。
1.2 方法
1.2.1 生物信息学分析 以酿酒酵母外泌蛋白质组(245 个外泌蛋白)为基础数据[14],利用 Uniprot(http://www.uniprot.org/uniprot/?query=&sort=score)数据库下载每个蛋白的氨基酸残基序列;Phobius软件[15](http://phobius.sbc.su.se)能够很好地区分膜蛋白的穿膜螺旋和信号中疏水区,但其预测信号肽的敏感性和精确性稍差,因此先利用此软件对每个外泌蛋白是否具有信号肽进行初步计算筛选;然后利用 SignalP4.1[16](http://www.cbs.dtu.dk/services/Signa)lP/对经 Phobius 计算初选的含有信号肽蛋白进行更精确的计算分析,阈值设定为 0.5,计算得分值越高含有信号肽的可信度也越高;最后再分别利用 PSORT II[17](https://psort.hgc.jp/form2.html)和 ProP1.0[18](http://www.cbs.dtu. dk/services/ProP/)对上述筛选获得的蛋白进行亚细胞定位和是否含有高尔基体蛋白酶 Kex2 的加工剪切位点进行预测分析,再利用 NetNGlyc1.0(http://www.cbs.dtu.dk/services/NetNGlyc/)进行糖基化位点预测分析。
1.2.2 表达质粒构建
1.2.2.1 含有EXG1 基因表达载体的构建 以酵母基因组 DNA 为模板,以PDB ID 1H4P 的核酸序列为基础设计合成引物(表 1),经 PCR 扩增获得编码EXG1的 DNA 片段,然后插入到表达载体 pδGAPh,该载体 pδGAPh 含有酿酒酵母三磷酸甘油醛脱氢酶(GAP)基因启动子、酿酒酵母δ 整合位点、酿酒酵母终止子磷酸甘油酸激酶 1(PGK1)和可利用潮霉素 B(HygB)进行筛选的HygB 激酶标记基因,经 CaCl2转化法转化大肠杆菌 Trans1-T1,筛选获得阳性克隆,即得到酵母整合表达质粒 pδGAPh-EXG1。
1.2.2.2 含有 α-Factor 信号肽元件融合基因表达载体的构建 以 pPIC9K(invitrogen)质粒为模板,利用 A_F1/A_F2 引物对(表 1)经 PCR 扩增获得 α-Factor 的信号肽功能元件,然后与 A_F3/A_F4引物对(表 1)所扩增的 EXG1 DNA 片段进行补平反应,最后再以 A_F1/A_F4 引物对扩融合DNA 片段,利用限制性内切酶NdeI 和EcoR I定向置换 EXG1 自身所对应的 DNA片段,即得到 pδGAPh-α-Factor-EXG1。
1.2.3 酵母转化 参照文献[19]报道的方法制备酵母感受态细胞和 DNA 转化,把转化体系涂布于含有相应抗生素的 YPD 平板上筛选,在 30 ℃ 培养箱培养 3 d。将获得的菌株转接于含高浓度抗生素的筛选平板上进行高拷贝整合阳性克隆复筛。
1.2.4 阳性克隆的鉴定 目的基因转化酵母细胞,筛选获得阳性克隆,然后采用酵母基因组 DNA提取试剂盒提取基因组 DNA。通过 PCR 鉴定确定目的基因是否整合进入宿主的基因组中,最后再以各基因的特异引物对纯化的 PCR 扩增片段进行测序验证。
1.2.5 菌体生物量测定 测定OD600光密度值:取 1 ml 发酵液,稀释一定倍数至OD600值在 0.2 ~0.8 之间,测定其OD600值,通过计算得到菌体生物量。
1.2.6 重组菌株外泌蛋白酶的活性测定 采用4-硝基苯-β-D-吡喃葡萄糖苷(pNPG)法测定 EXG1酶活[20]。配置不同浓度的对硝基苯酚,于 400 nm处测定吸光值并绘制标准曲线。酶活定义:每分钟释放 1 nmol 的对硝基苯酚所需要的细胞干重(mg)。挑取单克隆于 10 ml 液体 YPD 培养基中,30 ℃、220 r/min 培养 15 ~ 20 h,以起始OD600= 0.2 转接到 50 ml 液体 YPD 培养基中,30 ℃、220 r/min培养 24 h 后取适量发酵液置于灭菌的 1.5 ml EP管中,8000 r/min 离心 3 min,收集上清液。取上清液 50 μl,加入 50 μl 的 20 mmol/LpNPG(溶于 pH 6.0 的醋酸钠缓冲液中),混匀,30 ℃ 反应15 min,立即加入 100 μl 的 10%(W/V)Na2CO3终止反应,反应过程中释放出对硝基苯酚,利用PerkinElmer 酶标仪于 400 nm 测定吸光值。
2 结果
2.1 酿酒酵母外泌蛋白质组的生物信息学分析
2.1.1 外泌蛋白信号肽的计算确定 酿酒酵母与哺乳动物细胞一样具有依赖内质网-高尔基体分泌的经典和非经典的分泌途径[3],外泌蛋白如果经过经典途径分泌,则发生在内质网腔内信号肽酶或高尔基体腔内切蛋白酶 Kex2 的剪切,导致外泌蛋白质组的质谱分析检测不到信号肽的整个功能元件区。因此,本研究利用 Uniprot 对文献报道的245 个外泌蛋白所对应全长氨基酸残基序列进行下载整理,这些蛋白质氨基酸序列首先利用计算程序 Phobius 预测分泌信号肽和信号肽酶的剪切位点,然后再利用计算程序 SignalP4.1和 PSORT II进行验证,经计算分析总计获得 65 个含有信号肽结构的外泌蛋白,其中含有 prepro 区的蛋白16 个(表 2),仅含有信号肽 pre 区的外泌蛋白49 个,计算程序 Phobius 和 PSORT II 预测的不同剪切位点在表 2 中已标注。

表 2 酿酒酵母含有 prepro 区的信号肽Table 2 List of endogenous and exogenous signal peptides with prepro-region for Saccharomyces cerevisiae
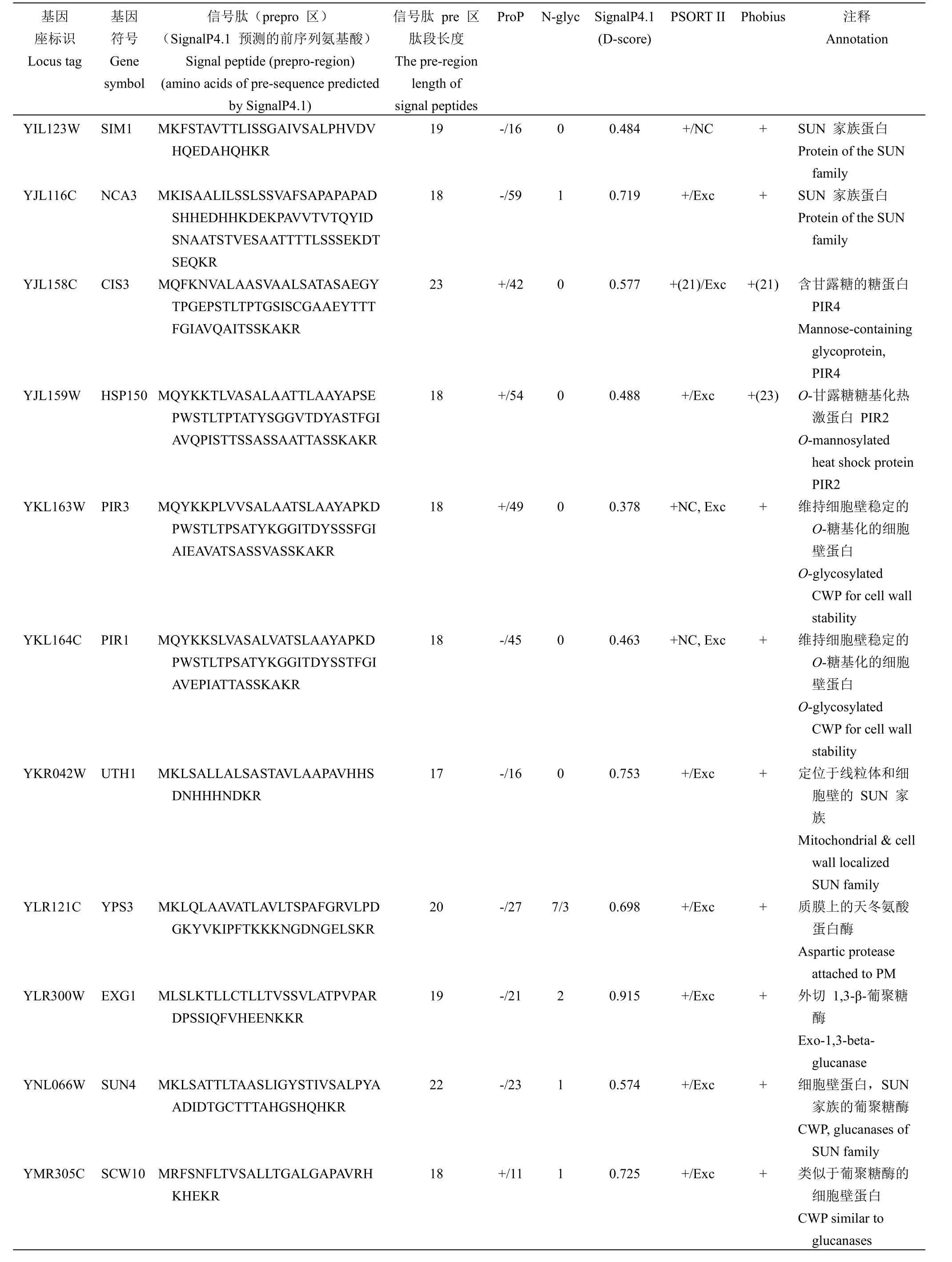
续表 2
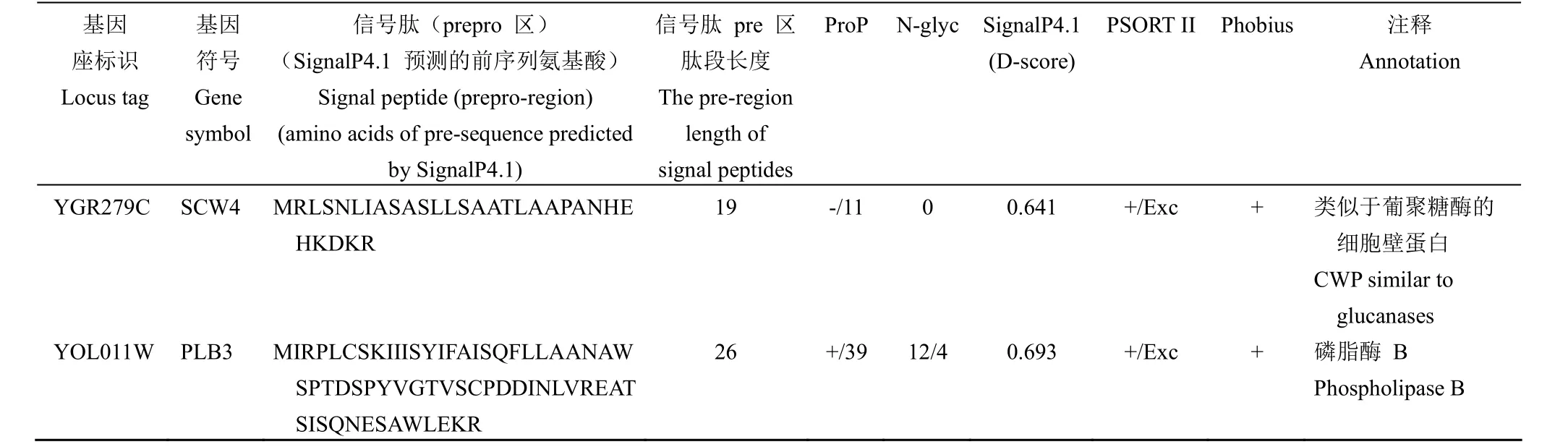
续表 2

图 1 外泌蛋白信号肽酶剪切位点的统计分析(P1 表示信号肽酶剪切位点-1)Figure 1 Statistical sequence analysis of predicted signal peptidase cleavage sites (P1 of the logos corresponds to the position -1 of signal peptidase cleavage site in the substrate)
2.1.2 信号肽酶剪切位点的结构 针对 65 个信号肽功能元件的计算分析结果可以看出,分泌型的信号肽含有明确的 N-、H- 和 C- 区,图 1 是对信号肽酶剪切位点 -6 到 -1 和 +1 到 +6 位氨基酸残基的分析结果。-3 和 -1 是信号肽酶特异识别的位点,-1 位置氨基酸残基的频次分别为 Ala(51)、Gly(6)、Ser(5);-3 位置氨基酸残基分别为高度保守的小的氨基酸残基 Val(26)、Ala(22)、Ile(7)、Ser(4);从图 1 也可以明显看出+1 位置多为小的疏水性的氨基酸残基 Ala、Leu、Ser;另一个重要而显著的结果为 Pro 在 +2 和 +4位置出现频次最高,再结合 -1 或 -6 位置出现Gly,可以得出在信号肽酶识别位点区形成一个β-转角或 Loop 区是一个必需的活性结构。
2.1.3 信号肽功能元件中前导区的糖基化位点 已有研究证实酿酒酵母 α-因子信号肽的前导区有 3 个糖基化位点,其位点突变导致融合外泌蛋白的分泌降低[21]。本研究利用 NetNGlyc1.0 计算程序对每个外泌蛋白进行糖基化位点预测分析,预测的糖基化位点与文献实际测定分析到的位点存在很大的差异[14],可能原因是外泌蛋白丰度本来就较低,而相应的肽段很难检测。在预测的含有前导区的 16 个信号肽功能元件仅有磷脂酶 B(PLB3)含有一个糖基化位点,已得到实验证实的信号肽功能元件如 CIS3、HSP150 与 α-因子信号肽的结构相似,能够引导融合异源蛋白的外泌表达,说明信号肽前导区的糖基化不是必需条件。图 2是含有前导区的信号肽功能元件的对比分析,可以看出有些信号肽功能元件如 EXG1、SCW4、SCW10和 SUN4 等含有较短的前导区,但已有研究结果证实 EXG1 被定位于高尔基体腔中的内切酶 Kex2剪切[22]。
2.1.4 信号肽功能元件中 Kex2 剪切位点的分析 外泌蛋白的信号肽如含有前导区,其分泌表达需要在高尔基体腔内被 Kex2 剪切加工,本研究利用计算程序 ProP1.0 进行预测分析,该软件的精确性较差,使用已得到广泛证实的 α-因子信号肽作对照进行分析,其预测分数为 0.374(阈值为 0.5),通过氨基酸序列的比对分析 16 个外泌蛋白含有前导区的信号肽,仅有 6 个信号肽的预测值大于0.5(表 2),从图 2 可以看出前导区 Kex2 剪切位点非常保守(2 个碱性的氨基酸残基 Lys-Arg)。
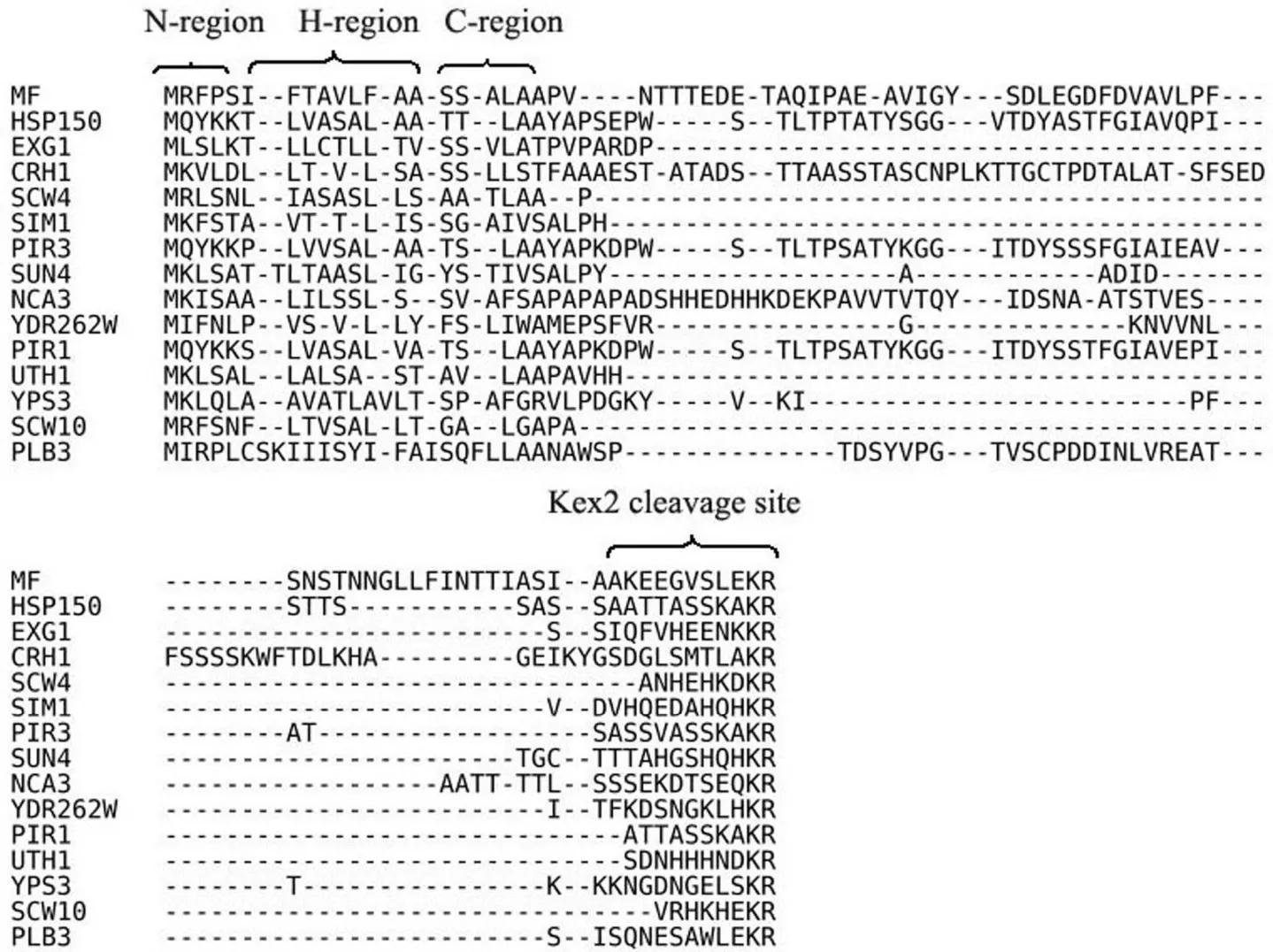
图 2 预测含有前导区的信号肽的比对分析Figure 2 Alignment of predicted signal peptidase and endopeptidase Kex2 cleavage sites

图 3 工程酵母基因组整合的目的基因的 PCR 鉴定(A:EXG1 DNA 片段;B:α-Factor-EXG1 DNA 片段)Figure 3 PCR verification results of genetically engineered strains with integration of two heterologous genes (A: EXG1 DNA fragments amplified by PCR; B: α-Factor-EXG1 DNA fragments amplified by PCR)
2.2 EXG1 信号肽功能元件的促分泌活性分析
2.2.1 含 EXG1 信号肽功能元件融合表达载体的构建和酵母转化 通过生物信息学分析获得的16 个含有前导区的信号肽,其中 CIS3、HSP150、PIR3、PIR1 为同一个家族的外泌蛋白,结构与α-因子信号肽相似,已有研究结果证实 HSP150 的信号肽功能元件具有与 α-因子信号肽相同的促进小鼠唾液酸转移酶分泌的表达水平[10];基于其他信号肽的前导区都较短的分析结果,于是推测其他的前导区是否具有较 α-因子信号肽更强的促分泌活性。本研究选取 EXG1 研究目标进行探索。首先利用 PCR 方法直接从酵母基因组中扩增获得EXG1 整个基因,亚克隆到酿酒酵母整合型表达载体 pδGAPh,即得到 pδGAPh-EXG1;同时利用α-因子信号肽功能元件作为参照,利用 SOE-PCR获得编码 α-因子信号肽 DNA 片段,直接置换EXG1 自身的信号肽编码区,获得整合型表达载体pδGAPh-α-Factor-EXG1。然后转化已基因敲除EXG1 的宿主菌 W303-1b/ΔES,通过潮霉素 B 抗性筛选标记进行阳性克隆的筛选,然后通过 PCR鉴定确定 EXG1 和 α-Factor-EXG1 是否整合进入宿主的基因组中,结果如图 3 所示阳性克隆都有预期的整合基因片段,进而纯化 DNA 片段并测序验证。
2.2.2 EXG1 信号肽功能元件的促分泌活性 EXG1 编码的外切 β-(1,3)-葡聚糖酶 1 具有水解pNPG 的 β-葡萄糖苷键的活性,利用对硝基苯酚绘制标准曲线(图 4),标准曲线线性良好;测定发酵上清中糖苷水解酶活性,结果表明使用EXG1 自身的信号肽比 α-因子信号肽引导的 β-葡萄糖苷酶水解活性更高,约是后者的 3 倍,同时也观察到 EXG1 的高表达对细胞没有生长抑制作用。因此,EXG1 的信号肽功能元件具有较好的促进融合蛋白分泌的活性(图 5)。

图 4 对硝基苯酚的浓度标准曲线Figure 4 Standard curve of p-nitrophenyl measured by PerkinElmer
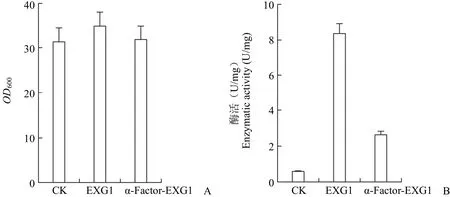
图 5 工程菌的生物量(A)和胞外 β-葡萄糖苷酶活性的测定(B)Figure 5 The determinations of biomass (A) and the supernatant enzymatic activities of the recombinant strains (B)
3 讨论
酿酒酵母之所以能成为众多药用蛋白和工业催化酶的生产表达系统,不仅仅是其具有遗传操作便捷、培养简单和繁殖迅速等优点,更重要的是其自身具有较强的 ER-Golgi 依赖的分泌途径[3]。外泌蛋白进行分泌表达的第一个关键步骤是外源蛋白的信号肽对分泌途径的选择性,依据信号肽识别体转运新合成的外泌蛋白到内质网的分子机制把外泌蛋白靶向内质网的转运过程区分为翻译共转移途径和翻译后转移途径。目前在酵母表达系统中应用最广泛的信号肽是 α-因子信号肽功能元件,利用翻译后转移途径驱动融合蛋白的外泌表达,为提高外源蛋白的表达量提供了一个靶点。
目前文献已报道了多种用于提高异源蛋白在酵母细胞高效表达的技术策略,主要包括对分泌途径中相关基因的敲除或高表达、优化表达载体系统和发酵过程及其调节伴侣蛋白促进内质网膜上的蛋白折叠和二硫键的形成[1]。但是如何提高异源蛋白在胞内从内质网到高尔基体的内膜系统囊腔内的转位依然是影响融合蛋白外泌效率的决定因素,其中信号肽功能元件是在该膜系统内转位的关键分子结构。过去很多研究集中在不同信号肽(实质上是信号肽的前导区)的发现鉴定研究,而对信号肽包括前导区的下游元件对外泌蛋白的影响研究相对较少[8]。本研究直接基于酿酒酵母细胞外泌蛋白质组的研究结果,采用生物信息学计算方法,快速发现具有促进蛋白外泌表达的信号肽功能元件。经计算分析获得了 65 个具有潜在分泌表达的信号肽,而这些蛋白绝大多数定位于细胞壁或胞外发酵液;进一步分析确定了含有前导区的 16 个信号肽功能元件,这与 Bae 等[10]利用翻译融合伴侣筛选系统分离鉴定具有较强促进人白细胞介素-2 表达的多个信号肽功能元件(如 ECM33、HSP150、CSI3 和 SCW4)得以相互印证,而后者还融合表达了信号肽功能元件下游更长的结构蛋白肽段。通过计算分析和结构预测可以看出在信号肽酶剪切位点下游的 +1 到 +6 的肽段含有导致 α-螺旋断开的 Gly 或 Pro 的氨基酸残基,推测可能形成易于信号肽酶识别剪切的 β-转角或环状结构,该结构是影响信号肽酶剪切的重要分子结构。
为验证通过计算方法获得的信号肽功能元件的可靠性和促进融合蛋白分泌的效率,选取其中与α-因子信号肽结构不同的 EXG1 的信号肽功能元件为研究目标,利用基因敲除 EXG1 的酵母为表达宿主消除内在 EXG1 的潜在影响因素。研究结果表明 EXG1 信号肽功能元件比常用的 α-因子信号肽功能元件具有更强的促蛋白分泌活性。总之,随着蛋白质组和转录组等系统生物学技术的发展,利用这些组学的研究结果并结合计算分析,可以快速地分离鉴定促进蛋白外泌表达的功能元件,这些元件的发现也将促进蛋白质工程研究和药用蛋白的生物合成。
