南海鸢乌贼微型群体的摄食习性研究
2018-08-07龚玉艳孔啸兰杨玉滔詹凤娉江艳娥陈作志
龚玉艳,孔啸兰,杨玉滔,詹凤娉,张 鹏,江艳娥,陈作志
(中国水产科学研究院南海水产研究所,农业部外海渔业开发重点实验室,广东广州 510300)
鸢乌贼(Sthenoteuthis oualaniensis)是广泛分布于印度洋、太平洋的赤道和亚热带等海域的经济种类,俗称深海红鱿鱼,在中国南海和印度洋西北部海域资源量较大[1-2]。近年来,国内外学者[3-7]有对鸢乌贼摄食习性做过一些研究,但研究对象基本为笼统的鸢乌贼,并没有对鸢乌贼的不同群体具体细分。我国南海海域内普遍存在鸢乌贼中型和微型两个群体,笔者在2016年已研究了鸢乌贼中型群体的食性[8],本文对鸢乌贼微型群体的食性再做研究。
水生生物食性研究从传统的胃含物分析法逐步发展到稳定同位素技术、脂肪酸标记技术等,并开始出现分子生物学手段[9-12],但目前食性分析应用最广泛的仍然是胃含物分析法。鸢乌贼具有发达的颚齿,摄食的大个体饵料生物均呈碎块状,仅采用胃含物分析法很难鉴别到最低分类阶元。BRALEY等[13]结合形态学和分子手段研究澳洲双柔鱼(Nototodarus gouldi)的食性发现,形态学方法鉴定出的饵料种类占所有已鉴定种类的50%,DNA条形码技术鉴定出的饵料种类占所有已鉴定种类的84%,且两种方法的结合优于单独使用任一种方法。因此,本研究在胃含物分析法的基础上尝试引入了新的技术手段——DNA条形码技术,以探讨分子技术在鸢乌贼食性分析中的可行性。目前国内运用分子手段研究头足类食性的较少,仅见于鸢乌贼中型群体食性的探索性研究[8]。
本研究根据2012年及2015年对南海海域进行灯光罩网采集的鸢乌贼微型群体样本,结合胃含物分析方法及DNA条形码技术,研究其食物组成、摄食强度、摄食习性与胴长的关系等,以期为南海鸢乌贼资源的合理开发和可持续利用提供理论依据。
1 材料与方法
1.1 调查海域及采集方法
鸢乌贼微型群样品分两次采集于南海外海海域,其中,2012年9-10月,在南海中、南部海域(110°34′~117°27′E、6°31′~14°53′N)选取 5个站位进行灯光罩网采集鸢乌贼样品,每站位随机挑选50尾保存。回实验室后根据LABE[14]对菲律宾西部海域和张鹏等[2]对南沙海域鸢乌贼群体的辨别方法,即胴背发光器官的有无和性腺发育的快慢,将采集的鸢乌贼分为中型和微型2个群体,其中微型群体样品共52尾(占总量的23.11%)。此外,2015年3月在南海北部海域(114°53′E、19°34′N)随机采集鸢乌贼微型样品169尾。

图1 南海鸢乌贼微型群体采样站位示意图Fig.1 Sampling sites of dwarf-form S.oualaniensis in the South China Sea注:S1~S5:2012年采样站位;SA:2015年采样站位Note:S1~S5:Sampling stations in 2012;SA:Sampling station in 2015
1.2 样品处理与分析
两次调查共采集鸢乌贼微型样品221尾,均带回实验室后测定其胴长、体质量、纯体质量、摄食等级、胃含物等生物学参数。摄食等级采用5级标准[15]。用滤纸吸干食物团表面水份,在精度为0.001 g的电子分析天平称量食物团总重,并在体视显微镜(Olympus SZ61)和光学显微镜(Olympus CX22)下鉴别饵料生物种类,并分别计数和称重。
鸢乌贼颚齿发达、消化能力强,胃内食物均为块状或半糜状,形态学上的分析只能从动物残肢及未消化的鱼类耳石进行比对鉴定,而用于耳石比对的鱼类均采自于南海海域。只针对数量较多的桡足类,以及体积较大的虾类、鱼类和头足类等饵料生物进行称重。
在分析鸢乌贼中型群体食性中发现,糜烂程度较大的胃含物通过DNA条形码技术也难以鉴定到最低分类阶元[8],因此该次实验选取了糜烂程度较小的食物块,共25份,均来自2015年采集的微型样品胃含物,其中24份在形态学上已鉴定为头足类不可辨认种类,另1份为虾类不可辨别种类。25份饵料生物样品均置于95%乙醇中保存,超纯水复水后提取基因组DNA,利用COⅠ基因通用引物扩增目的片段,进行COⅠ基因序列分析,进一步鉴定种类。
1.3 数据处理与分析
应用非参数 Kolmogorov-Smirnov Z检验分析南海中、南部鸢乌贼微型群体与北部微型群体的胴长分布是否具有显著性差异;
采用物种个数百分比(N%),出现频率(F%),饵料质量百分比(W%)和相对重要性指数百分比(%IRI)等指标,评价各种(类)饵料的重要性[16-17]:
IRI=(N%+W%)×F%
%IRI=(IRI/∑IRI)×100
用胃饱满指数(RI,%)和空胃率(VI,%)来评估鸢乌贼的摄食强度[3]:
RI(%)=饵料质量/纯体质量×100
VI(%)=空胃数/总胃数×100
2 结果与分析
2.1 胴长分布
经检查,南海中、南部鸢乌贼微型群体的胴长范围为56~123 mm,平均胴长为89.5 mm;而南海北部鸢乌贼微型群体的胴长范围为74~127 mm,平均胴长为100.0 mm,两群体的胴长分布有重叠,且南海北部群体的胴长分布范围和平均胴长均大于中、南部群体。非参数 Kolmogorov-Smirnov Z检验分析表明,南海中、南部鸢乌贼微型群体与北部微型群体的胴长分布存在显著性差异(P<0.05)。
2.2 饵料组成
在胃含物分析法的基础上,利用DNA条形码技术,对24个头足类不可辨认种类再作进一步鉴定,结果鉴定出其中13个头足类原不可辨认种类,共3种,其中2种已鉴定到种,分别为安达曼钩腕乌贼(Abralia andamanica)和菱鳍乌贼(Thysanoteuthis rhombus),另一种仅鉴定到科,为柔鱼科Ommastrephidae,检出率为52%。此外,还有1个虾类样品未通过此方法鉴定出来。
结合胃含物分析法和DNA条形码技术的鉴定结果可以看出(表1),南海鸢乌贼微型群体主要以小型游泳生物为食,并兼食浮游动物,其饵料组成包括头足类、虾类、桡足类、鱼类(主要为灯笼鱼科)等7大类41种(包括未鉴定到种及不可辨认种类)。
从饵料类群的出现频率看,鸢乌贼微型群体的饵料组成以头足类的F%最高,为79.90,占出现频率百分组成的36.38%;其次是虾类,F%为50.25,占出现频率百分组成的22.88%;桡足类和鱼类也经常被鸢乌贼微型群体所摄食,F%分别为27.14和14.57,分别占出现频率百分组成的12.36%和6.64%,其中丽隆水蚤(Oncae venusta)的F%为19.60,是桡足类出现频率最高的种类,占出现频率百分组成的8.92%;其它介形类、等足类和端足类则是鸢乌贼微型群体偶然摄食的对象,其F%和占出现频率百分组成比例较小,均不超过5.00和2.00%。
从饵料类群的个数百分比看,桡足类的N%最高(51.95),其中丽隆水蚤为37.39,在众多已鉴定到种的饵料生物中N%最高。其次是头足类(28.10);虾类为 11.47;鱼类为 5.50;其余类群的N%均较低。
从饵料类群的质量百分比来看,头足类的W%最高(60.91);其次是虾类,为32.97;鱼类为5.43;桡足类仅为0.69。
在鸢乌贼微型群体的各饵料生物类群中,头足类的%IRI最高(65.05);其次是虾类(20.43);桡足类为13.07;鱼类最低,为1.46。显然,头足类是南海鸢乌贼微型群体的优势饵料生物类群。
2.3 摄食强度及随胴长的变化
南海中、南部鸢乌贼微型群体的摄食等级1~4级占群体实胃数的比例分别为20.83%、27.08%、29.17%和22.92%,而南海北部鸢乌贼微型群体的摄食等级1~4级占群体实胃数比例分别为11.92%、35.10%、37.09%和15.89%。从图2鸢乌贼微型群体各胴长组与摄食等级百分比的关系可看出,南海中、南部群体在胴长≤100 mm时,摄食等级以2~4级为主,101~110 mm时以1、2级为主,≥111 mm时以3、4级为主;南海北部群体在胴长≤80 mm时,以空胃个体居多,≥81 mm时摄食等级以2、3级为主,摄食等级比例随胴长增加呈现总体上升趋势。
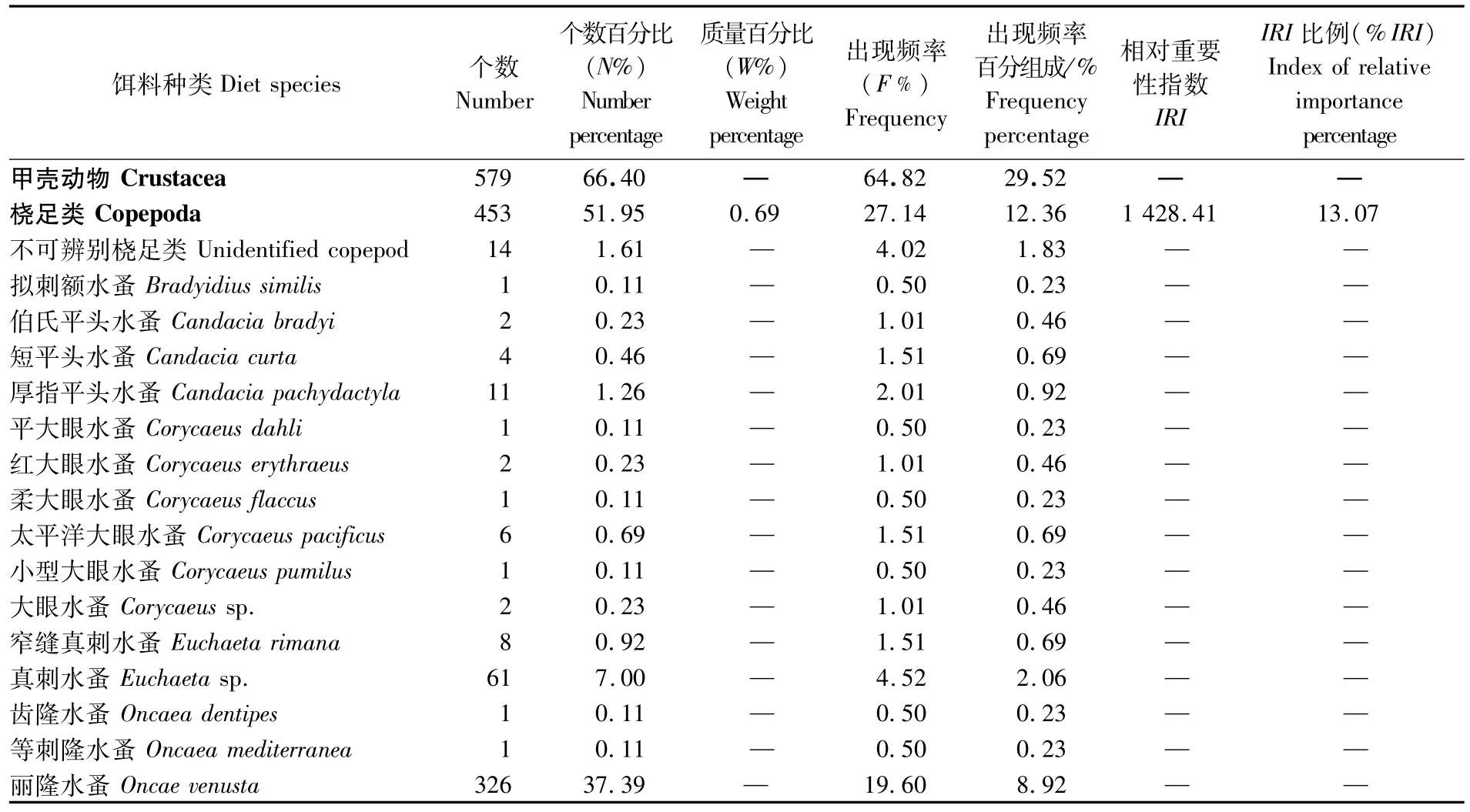
表1 南海鸢乌贼微型群体的饵料组成Tab.1 Diet composition of dwarf-form S.oualaniensis
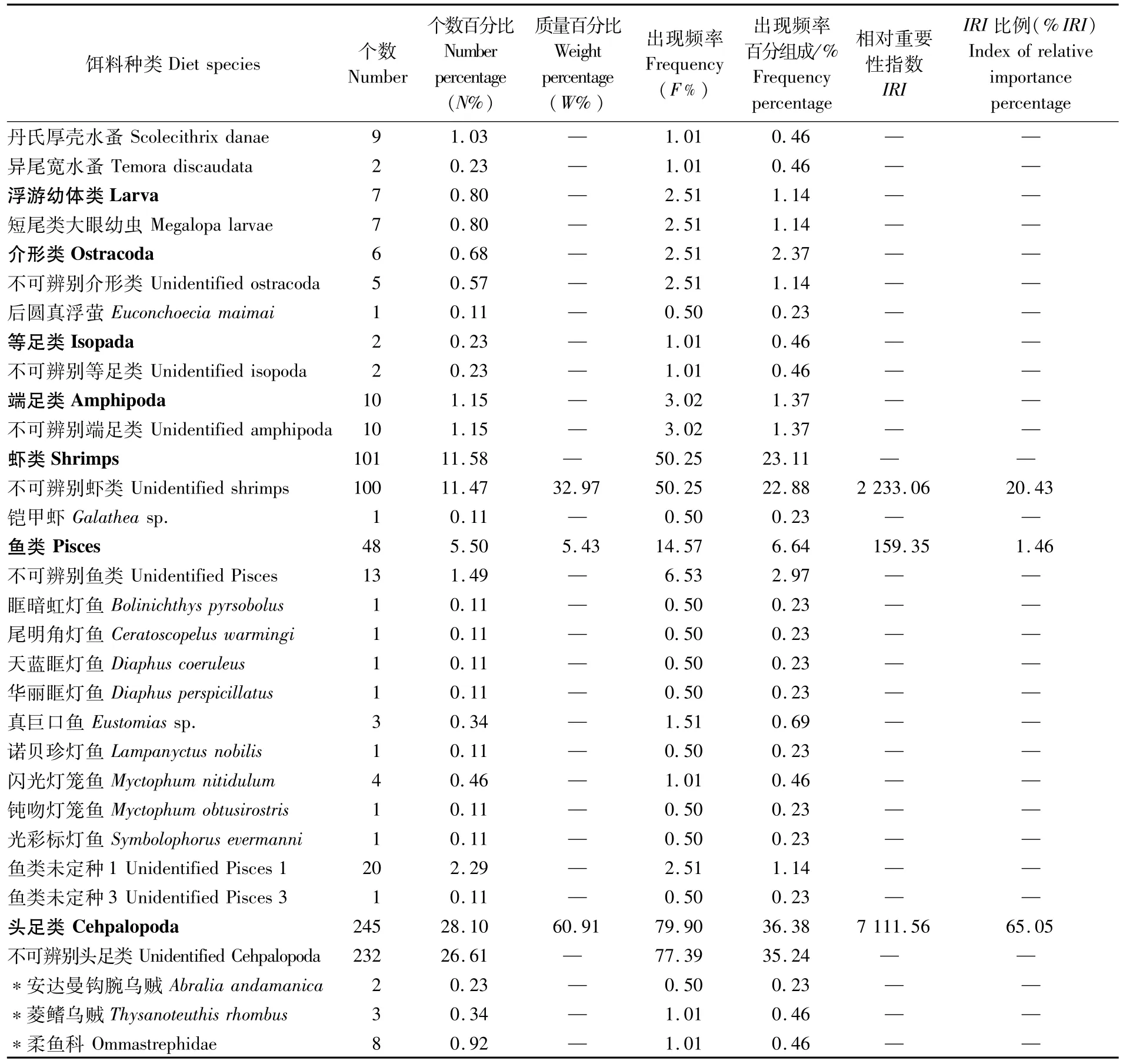
·续表1·
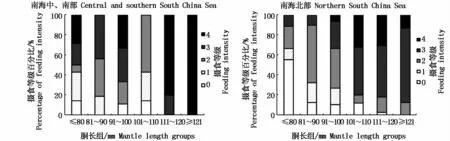
图2 不同胴长组鸢乌贼微型群体的摄食等级百分比Fig.2 Percentage of feeding intensity of dwarf-form S.oualaniensis in different mantle length groups
南海中、南部鸢乌贼微型群体的胃饱满指数(RI)最高值为7.70%,均值为1.73%,而空胃率(VI)均值为8.33%;南海北部群体的RI最高值为9.58%,均值为2.08%,VI均值为11.92%,高于南海中、南部群体。总体上,南海中、南部和北部群体的RI值均表现出高胴长组高于低胴长组,而VI值则相反(图3)。

图3 不同胴长组鸢乌贼微型群体的胃饱满指数(RI)和空胃率(VI)Fig.3 Stomach fullness index(RI)and percentage of empty stomach(VI)of dwarf-form S.oualaniensis in different mantle length groups
2.4 不同胴长组的摄食习性
鸢乌贼微型群体各胴长组主要食物类群的N%和F%分布见图4。南海中、南部鸢乌贼微型群体饵料类群的N%在胴长小于100 mm以前基本以头足类、鱼类和虾类为主,胴长在101~110 mm之间,以桡足类、头足类和鱼类为主,胴长大于111 mm时,头足类则占有绝对优势;头足类在各胴长组的F%均较高,虾类在胴长为81~90 mm和91~100 mm时,出现频率最高,均高于60.00,而在101~110 mm时,虾类F%迅速下降,鱼类和桡足类的F%上升,在胴长大于等于111 mm时,仅头足类F%最高,达100.00。南海北部鸢乌贼微型群体的N%在胴长小于90 mm时,以桡足类占绝对优势,胴长大于91 mm后,依然以桡足类为主,但其比例下降,而头足类比例升高;南海北部鸢乌贼微型群体的F%在各胴长组中,均以头足类为主,其次是虾类以及桡足类,鱼类在这一群体的食物中较少出现。
2.5 不同海域的摄食习性
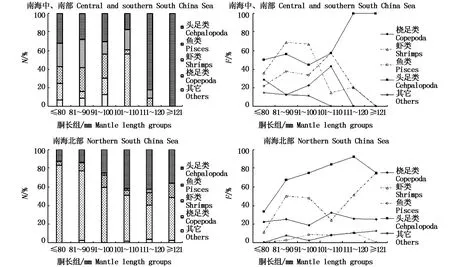
图4 不同胴长组鸢乌贼微型群体的个数百分比(N%)和出现频率(F%)Fig.4 N%and F%for major prey groups in diets of dwarf-form S.oualaniensis in different mantle length groups
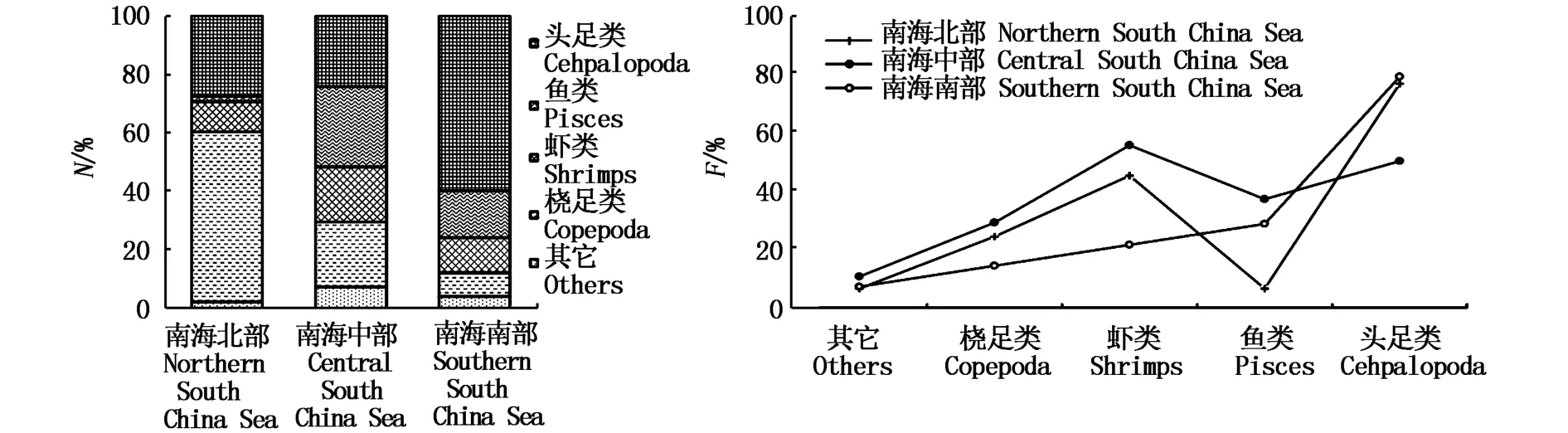
图5 不同海域鸢乌贼微型群体个数百分比(N%)和出现频率(F%)Fig.5 N%and F%for major prey groups in diets of dwarf-form S.oualaniensis in different survey areas
SA位于南海北部海域,S1~S3位于南海中部海域,S4和S5位于南海南部海域,鸢乌贼微型群体在3个海域有着不同的摄食习性,详细见图5。鸢乌贼微型群体在南海北部的饵料类群N%主要以小个体的桡足类为主,其次是头足类,与南海中、南部海域明显不同;南海中部海域鸢乌贼微型群体对各饵料类群均有摄食,各N%差别不大;而南海南部海域鸢乌贼微型群体的N%则以头足类最高,主要摄食大个体饵料。鸢乌贼微型群体的头足类饵料类群F%在各海域均表现较高,虾类F%在南海中部海域最高,北部和南部海域也占有一定比例;而鱼类F%在北部海域极低,桡足类和其它饵料类群的F%在3个海域均不高。
3 讨论
3.1 饵料组成
胃含物分析结果表明,南海鸢乌贼微型群体以头足类为主要摄食对象,%IRI达65.05;其次是甲壳动物虾类和桡足类,%IRI分别为20.43和13.07,属游泳动物和浮游动物食性类型。与南海鸢乌贼中型群体饵料组成相比[8],两者存在一定差异,南海鸢乌贼中型群体主要摄食头足类和鱼类,%IRI共达95.77,属游泳动物食性类型。
鸢乌贼主要摄食头足类、鱼类和甲壳动物,但随着栖息环境的不同和时间的推移,各饵料生物类群在食物中的比重存在明显的时空差异[8]。印度洋西北部鸢乌贼在2003年[18]的调查中主要以鱿鱼为食,在2004年[19]则主要以糠虾类和桡足类等甲壳动物为食。鸢乌贼在夏威夷主要摄食鱼类,其次是头足类[7],而在夏威夷群岛东北部其雌性群体则主要摄食甲壳类[20]。冲绳群岛鸢乌贼也主要摄食甲壳类[21]。张宇美等[12]逐月对西沙群岛鸢乌贼的研究发现,鸢乌贼在不同月份对鱼类和头足类的摄食偏好也不同。鸢乌贼中型群体的饵料类群在南海中、南部的比例也存在差异,头足类饵料类群的N%和F%在南海中部最高,鱼类饵料类群的N%和F%在南海南部最高,甲壳动物饵料类群的N%和F%在两海域的比例均较低[8]。在本次调查中,南海北部渔获的鸢乌贼微型样品胃含物里极少发现鱼类饵料生物,而桡足类发现较多,其N%达60.00,与南海中、南部以大个体饵料生物为主差别较大,这主要是因为两批样品采自不同年份不同季节,以及不同海域,同时受到时间与空间的双重作用影响;南海中部鸢乌贼微型群体对头足类、鱼类和虾类等较大个体饵料生物均有摄食,且各类饵料生物的比重差别不大;而南海南部则以头足类饵料生物占主要优势,与南海中部鸢乌贼微型群体的摄食偏好也出现了一些差异,但与南海北部相比,差异相对较小,主要受空间变化的影响。鱼类的摄食也普遍存在时空差异,韩东燕等[22]研究了胶州湾六丝钝尾虾虎鱼(Amblychaeturichthys hexanema)的摄食生态,发现六丝钝尾虾虎鱼的食物组成存在明显的季节变化,与胶州湾饵料生物种类和数量的季节性变化有关,而不同经、纬度海区的摄食差异也与胶州湾饵料生物分布的空间异质性有关。饵料生物的时空分布差异会影响鱼类的摄食差异,而鱼类摄食的时空变化在一定程度上也能反映饵料生物种类和数量的时空变化[22-23]。虽然目前对鸢乌贼摄食生态的研究未有鱼类深入,但鸢乌贼是一种广食性动物,其摄食的时空差异必然与饵料生物种类和数量的变动有关。钱卫国等[19]在研究印度洋西北部鸢乌贼渔场时发现,饵料生物如尖尾海萤(Cypridina acuminata)与中心渔场分布无关,而箭虫类、桡足类和糠虾类与中心渔场分布关系显著,可作为渔场分布的指示种类。
菱鳍乌贼是大型鱿鱼,成体一般比鸢乌贼大,被鸢乌贼摄食的机率理应较小,但利用DNA条形码技术在鸢乌贼微型群体的胃含物中检测到有菱鳍乌贼的DNA,且在同一胃含物发现的残留角质颚个体较小,因此我们推断鸢乌贼微型群体有可能摄食了菱鳍乌贼的仔鱼。DNA条形码技术对鸢乌贼微型群体胃含物的检出率为52%,这说明该技术在鸢乌贼的食性分析应用上具有一定可行性。但从该研究中也发现,乌贼的通用引物较差,且难以设计特异性引物,这可能是导致DNA条形码技术难以鉴定鸢乌贼微型群体胃含物到最低分类阶元的主要原因。此外,胃含物中糜烂程度较大的饵料生物样品之间容易互相污染,也有可能降低扩增结果的准确性。
3.2 食性变化
鸢乌贼随着生长发育,其摄食习性发生了一些变化。南海鸢乌贼中型群体随其胴长的增加,饵料组成中的小个体饵料类群比例降低,大个体饵料类群比例升高,符合“最佳摄食理论”[8]。除南海中、南部101~110 mm胴长组的大个体饵料类群在饵料组成中的比例明显偏低外,本次调查的鸢乌贼微型群体也存在相似的食性变化趋势。鸢乌贼中型群体偏好摄食个体较大的头足类和鱼类饵料生物,而胴长相对较小的微型群体还摄食一定量的小个体饵料生物,两群体饵料组成的差异显然与胴长大小有关。按照鸢乌贼胴长与食性的变化关系,南海北部微型群体的胴长较中、南部大,其食物中的大个体饵料生物比重理应也比中、南部大,但调查结果发现,南海北部微型群体饵料中的桡足类占比很高,大个体饵料生物占比低于中、南部。显然,胴长大小并不是导致南海北部和中、南部鸢乌贼微型群体出现摄食差异的主要原因。该研究的鸢乌贼微型样品采于不同海域及不同季节,胴长分布具有显著性差异,而饵料生物的时空分布差异会影响鱼类的摄食差异,南海北部和中、南部鸢乌贼微型群体出现的摄食差异很可能受海域饵料生物分布差异的影响,而两群体的摄食差异是否还受其他因素影响,饵料生物分布的差异对两海域微型群体摄食差异的影响程度大小,还需要以后再做深入研究。
3.3 摄食强度
南海鸢乌贼微型群体的摄食等级大部分以2、3级为主,低于中型群体的摄食等级[8]。除南海中、南部101~110 mm胴长组摄食等级偏低外,两海区的鸢乌贼微型群体摄食等级比例总体随胴长增加而升高,与中型群体摄食等级比例分布趋势相似,胴长越大,摄食等级越高,这也是南海鸢乌贼微型群体摄食等级普遍低于中型群体的主要原因。与以往对南海鸢乌贼研究的结果相似[3-4]。相对于较低胴长组,南海中、南部101~110 mm胴长组的摄食等级及大个体饵料类群在饵料组成中的比例均出现偏低的现象,这可能与该胴长组样本相对较少产生的偶然误差有关,其原因尚待进一步分析。
该研究的鸢乌贼微型群体样本来自不同年份和不同海域,其摄食习性可能受不同因素影响,从而降低两群体之间摄食习性的可比性。此外,两次采集的数量也存在较大差异,2012年仅采集了52尾,鸢乌贼微型群体食性随胴长变化过程中出现的异常,很可能就与胴长组样本量偏少有关;而该研究也考虑到样本量较少,故未进一步分析鸢乌贼不同性别的摄食差异。在分析方法上,该研究还运用了DNA条形码技术鉴定胃含物,虽然鉴定种类不多,但足以证明是一种可行的方法,值得改进并加以应用。张宇美[12]在2013年也结合胃含物分析法和碳氮稳定同位素方法研究了南海鸢乌贼摄食生态并估算出鸢乌贼在南海食物链中的大致营养位置。因此,在以后的研究中,有必要结合胃含物分析法、DNA条形码技术以及碳氮稳定同位素等方法,深入研究南海鸢乌贼不同种群、性别的摄食习性的时空变化、营养级及其主要影响因素,以期为南海鸢乌贼资源的评估、合理开发和可持续利用提供有力的理论依据。
致谢:感谢张俊、张魁、李玉芳、陈颖瑶等同事在鸢乌贼样品采集和生物学测定过程中给予了大力帮助,特致谢忱!
