不产桔霉素高产红曲色素的基因工程红曲菌株构建
2018-07-09高健信陈福生
李 琦,高健信,陈福生,李 牧*
(1.华中农业大学 食品科技学院,湖北 武汉 430070;2.广东科隆生物科技有限公司,广东 江门 529100)
红曲菌(Monascus)是一种丝状真菌,能够合成丰富的次级代谢产物,如红曲色素、莫纳可林K、γ-氨基丁酸等,被广泛应用于食品、医药、化工等行业[1-4]。其中,红曲色素作为一种天然微生物色素,以其稳定性高、着色效果好、生产不受原料与季节限制等优势被用作食品着色剂,在我国与东南亚地区已有着近两千年的应用历史[1]。红曲色素是国内增长最快的食品着色剂品种之一,年产量已超过1万t。
1995年有学者[5]发现,某些红曲菌株可以产生一种毒素—桔霉素,污染红曲色素产品,使红曲色素产品的安全性受到了挑战。根据现行国家标准GB 1886.181—2016《食品安全国家标准食品添加剂红曲红》规定,红曲红单位色价的桔霉素含量≤0.04 mg/kg。因此,控制红曲色素产品中桔霉素的含量成为红曲色素工业中的重要课题。
近年来,国内外学者通过多种方法来控制红曲色素产品中的桔霉素含量,并取得了一些重要进展,主要包括低产桔霉素菌株的筛选、发酵条件优化以及基因工程改造。已有学者通过筛选获得了低产桔霉素的红曲菌株[6],此外,通过优化发酵条件也能够控制桔霉素含量,如控制发酵过程pH值[7]、通氧量[8],以及利用低频磁场也能够抑制桔霉素的产生[9]。
近年来,随着丝状真菌基因组测序以及相关分子生物学技术的发展,红曲菌的分子生物学研究也取得了突破。桔霉素的生物合成途径涉及16个基因[10],其中聚酮合酶(polyketide synthase,PKS)是合成桔霉素的第一个关键酶,有学者将红色红曲菌(Monascusruber)中的该基因敲除,发酵产物中桔霉素的含量降低了99%[11]。因此,通过基因工程构建桔霉素合成基因的敲除菌株,能够有效抑制桔霉素的合成,有利于红曲色素的工业生产。
本研究在前期获得了一株高产红曲色素的紫色红曲菌J01,在马铃薯葡萄糖肉汤(potato dextrose broth,PDB)培养基中培养13 d,色价可达到265 U/mL,高于目前已报道的大多数红曲菌株[12],但该菌株也能产生一定量的桔霉素。因此,本研究首先分析了菌株J01中桔霉素合成基因的转录情况,并采用根癌农杆菌介导的转移脱氧核糖核酸(transfer deoxyribonucleic acid,T-DNA)转化技术敲除紫色红曲菌(Monasucs purpureus)J01的桔霉素合成关键基因pksCT,构建pksCT基因敲除菌株J42,并对其菌落形态、生物量、发酵产物中的桔霉素含量与红曲色素色价进行了分析。期望能够构建具有工业应用意义的不产桔霉素高产红曲色素的菌株,也为通过基因工程来改善丝状真菌中次级代谢产物合成过程提供了借鉴意义。
1 材料与方法
1.1 材料与试剂
1.1.1 菌株
紫色红曲菌(Monascus purpureus)J01、根癌农杆菌(Agrobacterium tumefaciens)EHA105:华中农业大学食品生物技术与安全实验室保藏。
1.1.2 培养基
马铃薯葡萄糖琼脂培养基(potatodextroseagar,PDA):马铃薯200 g,葡萄糖20 g,琼脂15 g,加蒸馏水至1 L,121 ℃湿热灭菌20 min。
PDB培养基:马铃薯200g,葡萄糖20g,加蒸馏水至1L,121℃湿热灭菌20 min。
1.1.3 化学试剂
pSKH质粒、双元载体pCAMBIA3300质粒:华中农业大学食品生物技术与安全实验室保藏;限制性核酸内切酶XhoI(10 U/mL)、SacI(10 U/mL)、HindIII(15 U/mL):宝生物工程(大连)有限公司;TRIZOL:美国Invitrogen有限公司;乙腈(色谱纯):美国Tedia公司;桔霉素标品(纯度98%):阿拉丁试剂(上海)有限公司。
1.2 仪器与设备
LC-20AT高效液相色谱仪:日本岛津公司;UV-1700紫外可见分光光度计:日本Shimadzu公司;ACQUITY UPLCXevo TQ MS高效液相串联四级杆质谱联用仪:美国Waters公司;SHD-250生化培养箱:上海精宏实验设备有限公司;HZ200LB型摇床:武汉瑞华仪器设备有限责任公司。
1.3 方法
1.3.1 桔霉素合成相关基因的克隆
参考红曲菌公布的桔霉素合成基因的序列信息,采用Oligo 6软件设计引物(见表1),由于pksCT基因有7 000 bp,直接扩增错配率较高,因此将其分3段扩增。

表1 桔霉素基因扩增引物信息Table 1 Primers for citrinin gene amplification
按照参考文献[13]报道的方法提取紫色红曲菌J01的总核糖核酸(ribonucleicacid,RNA),以总RNA为模板,反转录获得互补脱氧核糖核酸(complementarydeoxyribonucleic acid,cDNA)。以cDNA为模板对桔霉素合成的相关基因进行克隆,并对序列进行分析。
1.3.2 桔霉素合成关键基因pksCT的序列分析
克隆得到pksCT基因送去测序,根据测序结果对其基因结构进行分析。采用Softberry软件预测紫色红曲菌J01桔霉素合成基因簇中pksCT基因的结构及其编码的氨基酸序列。采用美国国家生物技术信息中心(national center of biotechnology information,NCBI)中的Blast功能对预测的基因的氨基酸序列进行比对分析。采用蛋白数据库Pfam预测基因所编码蛋白的活性结构域。
1.3.3 桔霉素合成关键基因pksCT的敲除
通过聚合酶链式反应(polymerase chain reaction,PCR)扩增技术,使用表2中的引物对HygF/HygR克隆质粒pSKH上的潮霉素B抗性基因hph,分别使用引物对CITF1/CITR1和CITF4/CITR4克隆紫色红曲菌J01基因组中桔霉素编码区的815bp上游同源臂(5'-UTR)和850 bp下游同源臂(3'-UTR)。通过重叠延伸PCR[14]技术,将3个片段进行融合,即得到敲除盒。采用SacI和HindIII分别双酶切敲除盒和双元载体pCAMBIA3300,纯化后的酶切产物经T4DNA连接酶连接,得到敲除载体PC-CIT,将敲除载体转入根癌农杆菌(Agrobacterium tumefaciens)EHA105中,与红曲菌J01孢子在28℃共培养3~10 d。
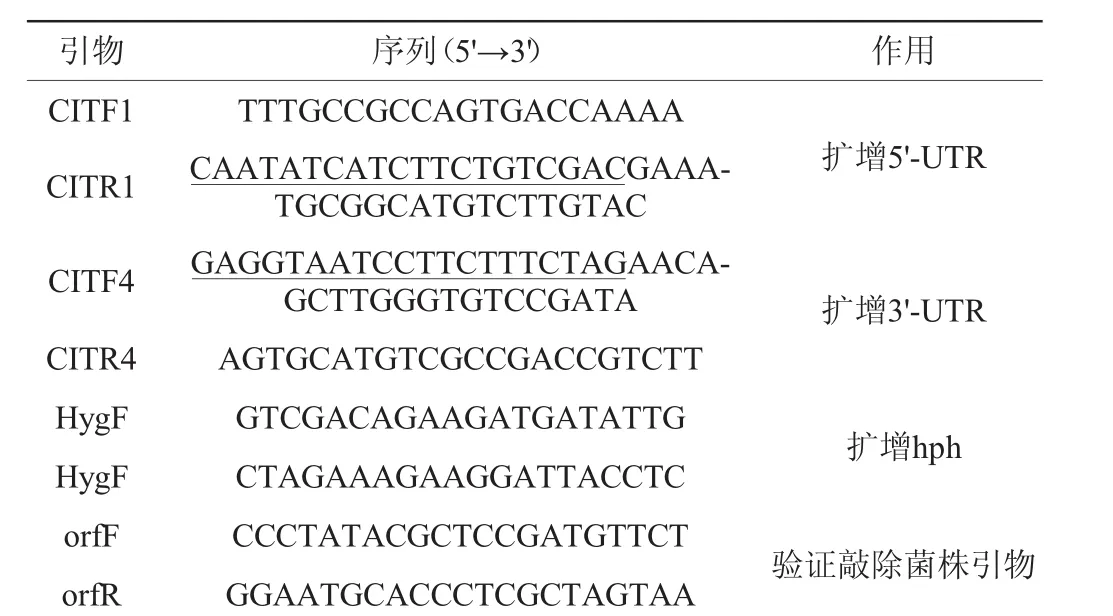
表2 本实验使用的引物Table 2 Primers used in this experiments
1.3.4 基因敲除菌株的筛选与验证
将基因敲除菌株接种到含有30 mg/L潮霉素的PDA平板上,28℃培养5 d,对菌株进行初步筛选。提取初步筛选到的基因敲除菌株的基因组DNA,以基因组DNA为模板,分别采用引物对HygF/HygR和orfF/orfRPCR扩增潮霉素抗性基因和orf基因片段,对基因敲除菌株进行验证。
1.3.5 pksCT基因的Southern杂交验证
在37℃条件下,利用限制性核酸内切酶XhoI酶切菌株J01和J42的基因组12 h,采用0.8%的琼脂糖凝胶电泳10 h后转移到Hybond+尼龙膜上,分别以hph基因(Probe 1)、pksCT基因的orf(Probe 2)作为探针在42℃条件下杂交16 h,进行免疫检测与显色。
1.3.6 菌落形态观察
将红曲菌J01和菌株J42接种到PDA斜面上,7d之后洗下孢子,制备孢子悬浮液并且将孢子悬浮液浓缩到106个/mL,接种1μL到PDA培养基上,28℃培养15d,观察其菌落形态。
1.3.7 生物量测定
将菌株J01和J42按照1%(V/V)的接种量分别接种于40 mL PDB培养基中,于28℃,120 r/min条件下振荡培养,发酵第15天时,将菌丝体和发酵液用滤布分离,并用蒸馏水多次洗涤菌丝体后将其转移至已称质量的离心管中,冷冻干燥至恒质量,计算菌丝体干质量。
1.3.8 桔霉素含量的测定
菌株的培养:按照1.3.7的方法进行培养。
桔霉素含量的测定:桔霉素的测定参考国标GB/T 5009.222—2016《红曲类产品中桔青霉素的测定》中的免疫亲和柱净化-高效液相色谱法(high performance liquid chromatography,HPLC)进行测定分析。为使测定结果准确,将上述样品按照文献[15]报道的方法进行高效液相色谱串联质谱(liquid chromatography tandem mass spectrometry,LC-MS/MS)分析。
1.3.9 红曲色素色价的测定
菌株的培养:按照1.3.7的方法进行培养。
红曲色素色价的测定:
(1)测定波长的选择:红曲色素主要由黄、橙、红三种类型的色素组成,各类色素对应的最大光吸收波长在380nm、470 nm和520 nm附近,因此,分别选取380 nm、470 nm和520 nm波长处的吸光度值计算黄、橙、红色素的色价。
(2)发酵液色素含量测定:用PDB培养基稀释发酵液,使其吸光度值在0.15~0.80范围内,分别测定波长380 nm、470 nm、520 nm处的吸光度值,按色价计算公式分别计算发酵液中黄、橙、红色素的色价。
(3)菌丝体色素含量测定:取冷冻干燥的菌丝体0.01g,采用液氮研磨破碎后装入2mL的离心管中,加入1mL体积分数为70%的乙醇水溶液,于60℃条件下水浴2h后,10000r/min离心5 min,取上清。用体积分数为70%的乙醇水溶液稀释上清液,使其吸光度值在0.15~0.80范围内,分别测定上清液在波长380 nm、470 nm、520 nm处的吸光度值,按色价计算公式分别计算菌丝体中黄、橙、红色素的色价,其计算公式如下:
色价=A×总稀释倍数。
式中:黄色素色价A为波长380 nm处的吸光度值;橙色素色价A为波长470 nm处的吸光度值;红色素色价A为波长520 nm处的吸光度值。
(5)总色价:参考国标GB 1886.19—2015《食品安全国家标准食品添加剂红曲米》中方法进行。
2 结果与分析
2.1 桔霉素合成相关基因的转录水平分析
在基因敲除之前,首先需要验证潜在的目标基因是否能转录。据报道,红曲菌桔霉素合成基因簇包括16个基因,分别为MRL7、MRL6、MRL5、MRL4、MRL3、MRL2、MRL1、pksCT、MRR1、MRR2、MRR3、MRR4、MRR5、MRR6、MRR7、MRR8,这些基因依次参与桔霉素的生物合成过程,且该合成过程已经基本清晰[10],因此,需要对所有与桔霉素合成相关的基因进行PCR扩增,分析其是否处于转录水平。以反转录得到的紫色红曲菌J01的cDNA为模板,对紫色红曲菌J01的桔霉素合成相关基因进行PCR扩增,结果见图1。
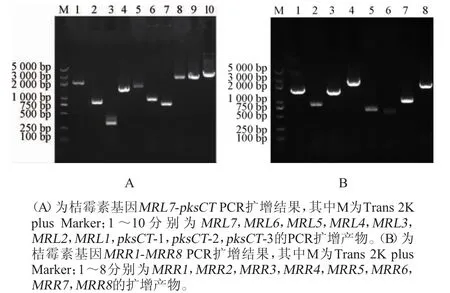
图1 紫色红曲菌J01桔霉素合成相关基因的PCR扩增结果Fig.1 PCR amplification results of citrinin-related syntetic genes fromM.purpureusJ01
由图1可知,从紫色红曲菌J01中克隆得到16个与桔霉素合成相关的基因,并且电泳条带大小与表1预测结果基本一致,说明这些桔霉素合成相关基因处于转录水平。PCR产物送去(测序公司)测序,经序列分析,结果表明,紫色红曲菌J01来源的16个与桔霉素合成相关基因与目前公布的桔霉素合成基因序列基本一致,进一步说明这些基因正常的参与桔霉素的合成,可被选择为基因敲除对象。
2.2 紫色红曲菌J01中pksCT基因的结构分析
根据目前报道的桔霉素生物合成途径,pksCT基因编码的蛋白为一种聚酮合酶(PKS),是合成桔霉素的关键酶,因此,分析了紫色红曲菌J01中的pksCT基因。从J01菌株的cDNA中克隆得到pksCT基因,全长共7 521 bp,编码2 435个氨基酸,其氨基酸序列与文献报道的紫色红曲菌中控制桔霉素生物合成的PKS(GenBank:BAD44749.1)序列覆盖度达100%,相似度为95%。
通过Pfam软件预测pksCT蛋白的结构域,结果如图2所示。由图2可知,pksCT蛋白包含多个关键的结构域,如起始单元酰基载体蛋白质转酰基酶(starter unit acyl carrier protein transacylase,SAT)、β-酮基合成酶(β-ketosynthase,KS)、酰基转移酶(acyltransferase,AT)、产物模板(product template,PT)、甲基转移酶(methyltransferase,CMeT)、酰基载体蛋白(acyl carrier protein,ACP)、还原酶(reductase,R)。聚酮合酶(PKS)中这些结构域的功能各不相同,如:KS主要负责催化前体的缩合反应,使得聚酮链得以延长;AT主要负责前体的选择;ACP主要负责前体和中间体的共价结合。聚酮合酶中KS、AT、ACP三个结构域是绝大部分PKS功能所必须的[16]。因此,敲除的目标基因片段至少要包括这三个部分,才能够确保pksCT基因不再发挥功能。
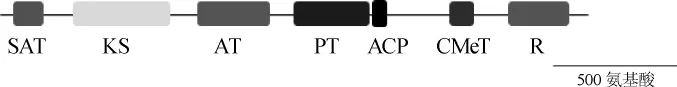
图2 pksCT蛋白结构域分析Fig.2 Analysis of pksCT protein structural domain
2.3 桔霉素合成关键基因pksCT的敲除
采用根癌农杆菌介导的T-DNA转化技术,利用红曲菌自身的同源重组系统实现pksCT基因的敲除。首先通过PCR扩增验证敲除载体是否转化至根癌农杆菌,验证结果见图3A。由图3A可知,在农杆菌中,存在5'-同源臂、hph抗性基因、3'-同源臂以及敲除盒的片段,并且条带大小符合预期,说明敲除载体成功转化到根瘤农杆菌中。
图3 pksCT基因敲除载体扩增验证(A)及菌株J42的PCR扩增结果(B)和Southern blot验证(C)Fig.3 Verification ofpksCTgene deletion cassette amplication(A)and PCR amplication results of strain J42(B)and Southern blot verification(C)
然后通PCR扩增验证紫色红曲菌中的pksCT基因是否被敲除,PCR扩增结果见图3B,由图3B可知,PCR扩增得到了2.1 kb的唯一条带,未扩增到缺失的orf基因,因此,可以将该菌株初步鉴定为敲除菌株,命名为J42。
最后,进一步对该基因敲除菌株进行Southern杂交验证,结果如图3C所示。由图3C可知,以hph基因(Probe1)为探针,在敲除菌株中出现了单一杂交带,而以orf基因(Probe2)为探针,则未出现杂交带,说明pksCT基因已被抗性基因hph替换,且hph以单拷贝的形式存在于菌株J42中,说明菌株J42成功敲除pksCT基因。
2.4 基因敲除菌株与原始菌株的生长情况比较
菌落形态比较:将原始红曲菌株J01和基因工程菌株J42点接到PDA培养基上,28℃培养15 d,观察菌落形态,结果如图4所示。由图4可知,菌株J42的菌落颜色呈橙黄色,菌落直径为6.8 cm,与原始菌株J01相比,无明显差异,说明其生长情况较为一致。
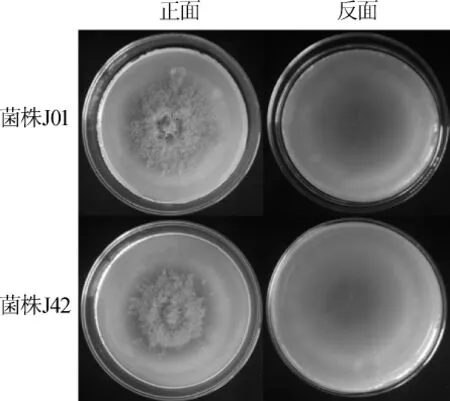
图4 紫色红曲菌株J01和J42菌落形态Fig.4 Colony morphology ofM.purpureusJ01 and J42
生物量比较:红曲菌株J01和J42经PDB培养基培养15d,对菌丝体的生物量进行测定。菌株J42生物量为4.43g/L,菌株J01的生物量为4.12g/L,两者之间无显著性差异(P>0.05),进一步说明pksCT基因的缺失红曲菌株的生长无显著性影响。
2.5 桔霉素含量及红曲色素色价的测定
2.5.1 桔霉素含量的测定
采用免疫亲和柱净化-高效液相色谱法测定菌株J01和J42菌丝体的桔霉素含量,结果见图5。由图5可知,紫色红曲菌J01的桔霉素产量为5.1mg/kg。pksCT基因敲除后,菌株J42敲除菌株未检出桔霉素,为进一步验证实验结果,对样品进行了LC-MS/MS分析,其对桔霉素的检出限为1.0 μg/kg,仍未检测到桔霉素,并且在发酵液中也未检测到桔霉素,说明pksCT基因的缺失阻断了桔霉素的合成,这与SHIMIZUT等[17-18]的研究结果一致。
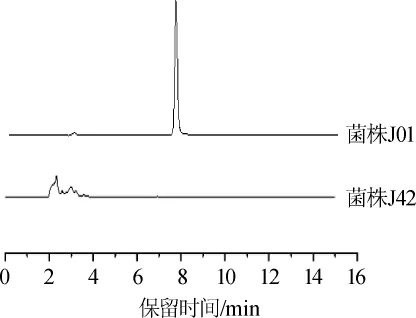
图5 紫色红曲菌J01和J42桔霉素产量的HPLC分析结果Fig.5 HPLC analysis results of citrinin yield ofM.purpureusJ01 and J42
2.5.2 红曲色素色价的测定
采用紫外分光光度法,分别测定了紫色红曲菌J01和J42发酵液和菌丝体色价,结果如图6所示。由图6可知,在发酵液中(图6A、6C、6E),菌株J42的色价分别为0.93 U/mL、0.31U/mL、0.24U/mL,较J01相比无显著性差异(P>0.05),在菌丝体中(图6B、6D、6F),菌株J42的黄色素、橙色素和红色素色价分别为1 877 U/g、773 U/g、1 068 U/g,显著高于菌株J01(P<0.01),说明pksCT基因的缺失不会降低菌株的产色素能力。从总色价来看,敲除菌株J42的总色价为415 U/mL,是原始菌株的1.56倍,分析原因可能是pksCT基因敲除后,用于合成桔霉素的大部分能量供给了色素合成,促进了色素产量的提高。

图6 紫色红曲菌J01和J42菌株发酵液(A,C,E)和菌丝体(B,D,F)色价的比较Fig.6 Comparison of the color value in fermentation broth(A,C,E)and mycelium(B,D,F)ofM.purpureusJ01 and J42
3 结论
本研究在前期发现的一株高产红曲色素的紫色红曲菌J01基础上,基于桔霉素合成基因序列测序与分析,依据桔霉素生物合成途径,选择聚酮合酶pksCT基因为敲除对象,通过基因工程方法成功将紫色红曲菌J01基因组上的pksCT基因敲除,构建工程菌株J42。通过菌落形态观察和生物量的测定,得出菌株J42与原始菌株J01的菌落形态及生物量无显著性差异(P>0.05)。通过HPLC和LC-MS/MS检测得出,菌株J42不产桔霉素,且红曲色素总色价为415 U/mL,较原始菌株J01提高1.56倍。所以,本研究成功构建一株不产桔霉素高产红曲色素的基因工程红曲菌株J42。
[1]CHEN W,HE Y,ZHOU Y,et al.Edible filamentous fungi from the speciesMonascus:early traditional fermentations,modern molecular biology,and future genomics[J].Compreh Rev Food Sci Food Safety,2015,14(5):555-567.
[2]FENG Y L,SHAO Y C,ZHOU Y X,et al.Effects of glycerol on pigments and monacolin K production by the high-monacolin K-producing but citrinin-free strain,Monascus pilosusMS-1[J].Eur Food Res Technol,2015,240(3):635-643.
[3]杨 洋,陈 冬,达文燕,等.红曲、红曲霉和红曲色素[J].生物学通报,2017,52(7):1-3.
[4]周文斌,贾瑞博,李 燕,等.红曲色素组分、功效活性及其应用研究进展[J].中国酿造,2016,35(7):6-10.
[5]BLANC P J,LAUSSAC J P,BARS J L,et al.Characterization of monascidin A fromMonascusas citrinin[J].Int J Food Microbiol,1995,27(2-3):201-213.
[6]蒋冬花,冯青青,任 浩,等.高产莫纳可林K低产桔霉素红曲霉菌株的筛选和发酵条件初步优化[J].微生物学杂志,2016,36(6):10-16.
[7]XIONG X,ZHANG X,WU Z,et al.Optimal selection of agricultural products to inhibit citrinin production during submerged culture of Monascus anka[J].Biotechnol Bioproc Eng,2014,19(6):1005-1013.
[8]YANGJ,CHENQ,WANGW,et al.Effect ofoxygen supplyonMonascus pigments and citrinin production in submerged fermentation[J].J Biosci Bioeng,2015,119(5):564-569.
[9]WAN Y,ZHANG J,HAN H,et al.Citrinin-producing capacity of Monascus purpureusin response to low-frequency magnetic fields[J].Process Biochem,2017,53:25-29.
[10]HE Y,COX R J.The molecular steps of citrinin biosynthesis in fungi[J].Chem Sci,2016,7(3):2119-2127.
[11]季宏飞.红色红曲菌pksCT基因缺失株的构建及其固液态发酵生产红曲色素的研究[D].南昌:南昌大学,2008.
[12]许楚旋,任 浩,章 婷,等.高产红曲色素低产桔霉素紫色红曲霉转化子筛选与代谢产物分析[J].山西农业科学,2016,44(3):318-322.
[13]丁晓莉.红色红曲菌M7聚酮合酶与氧化还原酶在米曲霉中的异源共表达研究[D].武汉:华中农业大学,2015.
[14]YU J H,HAMARI Z,HAN K H,et al.Double-joint PCR:a PCR-based molecular tool for gene manipulations in filamentous fungi[J].Fung Genet Biol,2004,41(11):973-981.
[15]JI X,XU J,WANG X,et al.Citrinin determination in red fermented rice products by optimized extraction method coupled to liquid chromatography tandem mass spectrometry(LC-MS/MS)[J].J Food Sci,2015,80(6):1438-1444.
[16]ALICE Y C,NATHAN A S,KIM C Y,et al.Extender unit and acyl carrier protein specificity of ketosynthase domains of the 6-deoxyerythronolide B synthase[J].J Am Chem Soc,2006,128(9):3067-3074.
[17]SHIMIZU T,KINOSHITA H,ISHIHARA S,et al.Polyketide synthase gene responsible for citrinin biosynthesis inMonascus purpureus[J].Appl Environ Microbiol,2005,71(7):3453-3457.
[18]FU G,XU Y,LI Y,et al.Construction of a replacement vector to disrupt pksCTgene for the mycotoxin citrinin biosynthesis inMonascus aurantiacus and maintain food red pigment production[J].Asia Pac J Clin Nutr,2007,16(S1):137-142.
