PPP2R2A在乳腺癌细胞中结合GFPT2并导致其去磷酸化
2018-06-26李笑荣张进马端
李笑荣,张进,马端
复旦大学 基础医学院,上海 200012
PP2A是一种重要的丝氨酸/苏氨酸磷酸酶,是由调节亚基 (B)、支架亚基 (A) 和催化亚基(C) 组成的异三聚体,其中调节亚基B决定PP2A磷酸酶复合物的底物特异性和亚细胞定位[1-2]。目前在人类细胞中共发现有12种PP2A调节亚基,PPP2R2A是PP2A中调节亚基之一[3]。以往的研究表明,PPP2R2A参与肿瘤的发生发展,促肿瘤细胞生长[4-7]。例如,胰腺癌细胞中 PPP2R2A的表达显著增加,可激活多条促肿瘤发生的信号通路 (AKT-,ERK-,Wnt-)[8];肿瘤细胞中的PPP2R2A/PP2A可催化将c-Jun的T239位点去磷酸化,促进其与 DNA结合从而促进细胞的增殖和迁移[4];此外,肿瘤组织谷氨酰胺的缺乏可诱导PPP2R2A表达的上调,并抑制P53通路,从而促进肿瘤细胞生存[9]。
本课题组前期研究也发现,在乳腺癌细胞中下调PPP2R2A后,细胞增殖和迁移的能力显著下降。由于PPP2R2A是磷酸酶复合物的调节亚基,决定了PPP2R2A-PP2A磷酸酶复合物的底物特异性,因此寻找PPP2R2A的相互作用蛋白质,有助于阐明 PPP2R2A的作用机制。本研究通过SBP-HIS8串联纯化分离了PPP2R2A蛋白复合物,进而通过 HPLC-Chip-ESI/MS/MS分析其蛋白组分。蛋白质质谱结果显示 PPP2R2A与 GFPT1/GFPT2结合。GFPT1/GFPT2是己糖胺途径的限速酶,催化谷氨酰胺水解为谷氨酸,6-磷酸-果糖转化为 6-磷酸-葡萄糖胺。然后经一系列反应产生UDP-GlcNAC,而UDP-GlcNAC是蛋白质O-连接丝氨酸-苏氨酸的糖基化修饰以及糖原、糖脂修饰的糖基供体[10-11]。在本研究中,我们将进一步确认 PPP2R2A与 GFPT1/2间的相互作用,以及对GFPT1/2磷酸化和下游蛋白糖基化的影响。
1 材料与方法
1.1 材料
人胚肾细胞株HEK293T、HEK293和人乳腺癌细胞 MDA-MB-231购自 ATCC;大肠杆菌DH5α由本实验室扩增提供;慢病毒包装用辅助质粒 (TAT/GAG/VSVG/REC) 购自 SBI公司;Streptavidin-beads购自 GE公司;Protein A/G beads购自 Roche公司;GST beads购自 Sigma公司;单克隆兔源抗 PPP2R2A抗体和单克隆兔源抗GAPDH抗体购自Bioworld公司;单克隆鼠源FLAG抗体购自Abmart;单克隆鼠源 GST抗体和单克隆兔源HA抗体购自Sigma 公司;兔源PKA substrate购自CST公司;Dylight 488以及Dylight 594购于Jackson Immuno Research。
1.2 方法
1.2.1 重组质粒构建
构建 PCDH-FLAG3-HIS8-SBP-Puro重组质粒,首先设计SBP引物,并添加连续编码8个HIS氨基酸序列标,PCR扩增得到目的片段,纯化后酶切,与酶切处理的PCDH-FLAG3-Puro质粒进行连接,质粒经转化,然后菌落经PCR鉴定扩大培养阳性克隆。最后重组质粒测序鉴定。重组质粒PCDH-FLAG3-hyg-GFPT1/PCDH-FLAG3-hyg-GFPT 2/PCDH-FLAG3-HIS8-SBP-PPP2R2A-Puro/mGSTHIS-GFPT1/mGST-HIS-GFPT2/PCDH-FLAG3-hyg-PP P2R2A以及mGST-HIS-PPP2R2A构建方法同上,其中引物如表1所示。
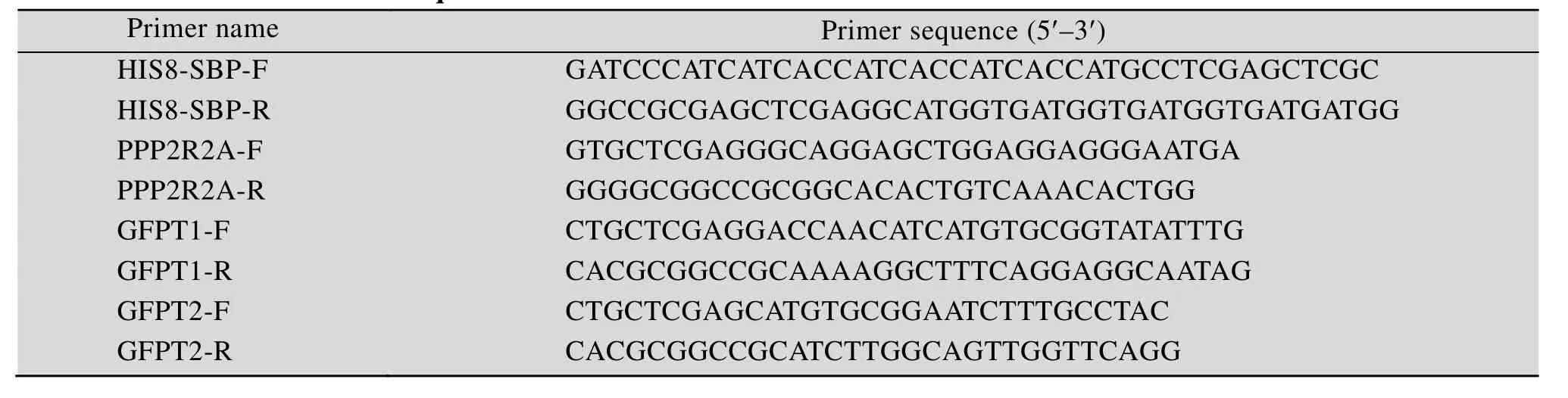
表1 引物名称和序列Table 1 Primer names and sequences
1.2.2 细胞稳转株构建
以HEK293T为慢病毒包装细胞,按照说明,将4种辅助质粒按照和基因表达质粒等比例混合即 Rec∶VsVg∶GAG∶TAT∶目标质粒=1∶1∶1∶1∶4,脂质体 Lipofectamine介导转染,24 h后收集病毒上清,2 000 r/min离心,上清中含有组装好的慢病毒加入细胞中,同时加入2.5 ng/mL 聚凝胺 (Polybrene) 促进感染,同上分别收集 48 h以及72 h的病毒并感染细胞。最后一次病毒感染48 h后,使用潮霉素加压筛选细胞稳转株。
1.2.3 串联亲和纯化联合HPLC-Chip-ESI/MS/MS分析
收集稳定表达 SBP-HIS-FLAG-PPP2R2A的细胞稳转株MDA-MB-231,加入15 mL细胞裂解液 (含1% Triton X-100,1 mol/L巯基乙醇) 冰上裂解20 min,离心收集全部细胞上清。第一次链霉柔和素 (Streptavidin) 纯化,蛋白上清加入Streptavidin磁珠,4 ℃过夜;弃上清,用细胞裂解液 (含0.1% Triton X-100,1 mol/L巯基乙醇) 洗涤5遍,每次3 min,最后保留1 mL左右含有沉淀复合物混悬液,加入400 μL 生物素洗脱液 (无EDTA/EGTA,含2 mmol/L biotin,20 mmol/L 咪唑),4 ℃旋转4 h后,收集上清。第二次Ni磁珠纯化,蛋白上清加入40 μL PBS平衡后的Ni磁珠,4 ℃旋转2 h;离心弃上清,加入0.05 mmol/L的NH4HCO3(pH 8.0),4 ℃ 静置5 min,离心弃上清,重复2次;加入10 mmol/L的NH4HCO3,用2 μg胰酶酶切过夜,取出酶切产物,2 000 r/min离心1 min (4 ℃),收集上清加入2 μg胰酶继续上述酶切4 h (37 ℃),2 000 r/min离心1 min,收集上清。最后HPLC- Chip-ESI/MS/MS分析,上清样品经过色谱分离后,肽段先后进入一级质谱和二级质谱,从而进行分析和鉴定。
1.2.4 细胞免疫荧光
将高压灭菌的盖玻片蘸于 0.1%明胶中后放至12孔板中;HA-PPP2R2A和PCDH-hyg-FLAGGFPT1/2重组质粒依据产品说明书进行脂质体Lipofectamine@2000瞬时转染,转染6 h后消化细胞,将其接种在明胶上生长,24 h后用4%多聚甲醛固定细胞10 min,PBST (0.2% Triton X-100)洗涤3次,5 min/次,3% BSA封闭1 h;取出玻片加入兔源抗HA一抗和鼠源抗FLAG一抗混合液 (1∶100),湿盒中4 ℃过夜;PBST (0.2% Triton X-100) 洗涤3次,5 min/次,再加入Dylight 488标记羊抗鼠IgG荧光二抗以及Dylight 594标记羊抗兔 IgG 荧光二抗的混合液 (1∶10),室温孵育1 h,需避光;PBST (0.2% Triton X-100) 洗涤3次,5 min/次,DAPI染细胞核,避光静置5 min;封片并荧光显微镜下观察。
1.2.5 GST Pull-down
PCDH-hyg-FLAG-GFPT1和mGST-HIS-PPP2R2A重组质粒依据产品说明书进行脂质体Lipofectamine@2000瞬时转染,24 h后收集细胞抽提蛋白;蛋白上清分成2份,一份取50 μL用作input,剩余的细胞裂解液全部用于GST Pull-down,按每个蛋白样品需要20 μL GST磁珠,取适量的磁珠用细胞裂解液清洗3次,将GST磁珠加入待测样品中,4 ℃旋转孵育过夜;次日,小于500×g离心2 min,弃上清,细胞裂解液清洗磁珠 3次,磁珠沉淀加入50 μL 的 SDS 上样缓冲液,95–100 ℃ 变性 5 min,12 000 r/min离心5 min;Western blotting检测目的蛋白。
1.2.6 免疫共沉淀 (Co-IP)
PCDH-hyg-FLAG-PPP2R2A和mGST-GFPT1/2重组质粒依据产品说明书进行脂质体 Lipofectamine@2000瞬时转染;24 h后收集细胞抽提蛋白,一份取50 μL用作input,剩余的全部用于免疫共沉淀;按每个样品30 μL×2磁珠,取适量的蛋白A/G,用细胞裂解液清洗3次;蛋白样品与30 μL蛋白A/G混匀,4 ℃旋转孵育 1 h;小于 500×g离心2 min,弃沉淀,上清加入鼠源FLAG (2 μg) 抗体,4 ℃ 旋转孵育 1 h,再加 30 μL Protein A/G,4 ℃旋转孵育过夜。次日,小于500×g离心2 min,弃上清。用细胞裂解液洗磁珠3次,磁珠沉淀加入50 μL 的SDS 上样缓冲液中,95–100 ℃变性 5 min,12 000 r/min 离心5 min;Western blotting检测目的蛋白。
2 结果与分析
2.1 串联亲和法联合HPLC-Chip-ESI/MS/MS分析筛选出PPP2R2A结合蛋白GFPT1/2
通过慢病毒感染建立SBP-HIS-FLAG-PPP2R2A稳定表达的 MDA-MB-231乳腺癌细胞,Western blotting检测PPP2R2A表达结果显示,除了内源性的PPP2R2A条带外,在稳转细胞内还有因加入纯化标签而变大的 SBP-HIS-FLAG-PPP2R2A条带,与内源性PPP2R2A表达水平相当 (图1)。先通过SBP-HIS8串联纯化分离PPP2R2A蛋白复合物,进而通过 HPLC-Chip-ESI/MS/MS鉴定与PPP2R2A相互作用的蛋白。蛋白质质谱鉴定结果显示,除了PPP2R2A蛋白外,还有PP2A复合物蛋白PPP2R1A和PPP2CA,这表明了该方法的可靠性。在被鉴定的蛋白中,GFPT1和GFPT2同属于 L-谷氨酰胺-D-果糖-6-磷酸转氨酶,并且均有多个肽段被鉴定 (表 2),因此我们将进一步分析PPP2R2A与GFPT1/2间的相互作用。
2.2 PPP2R2A蛋白与GFPT1/2蛋白相互作用的确认
在 HEK293T细胞中,重组质粒共瞬时转染PCDH-hyg-FLAG-GFPT1/2和mGST-HIS-PPP2R2A,设置瞬转PCDH-hyg-FLAG空载和mGST-HIS空载作为阴性对照。GST Pull-down结果显示PPP2R2A可沉淀细胞中的 FLAG-GFPT1和 FLAG-GFPT-2(图 2A、B);反向,使用 FLAG抗体通过 Co-IP分别沉淀FLAG-GFPT1和 FLAG-GFPT2融合蛋白,结果同样表明二者均可沉淀细胞中的PPP2R2A蛋白。进一步说明PPP2R2A与GFPT1/2存在相互作用。
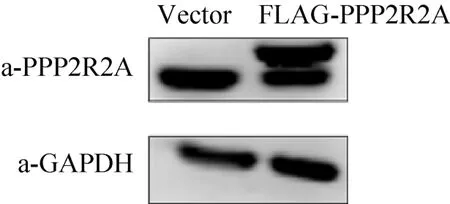
图1 Western blotting检测SBP-HIS-FLAG-PPP2R2A稳定细胞株的PPP2R2A表达水平Fig. 1 Detection of the expression of PPP2R2A in the stable MDA-MB-231 cells of SBP-HIS-FLAG-PPP2R2A by using Western blotting.
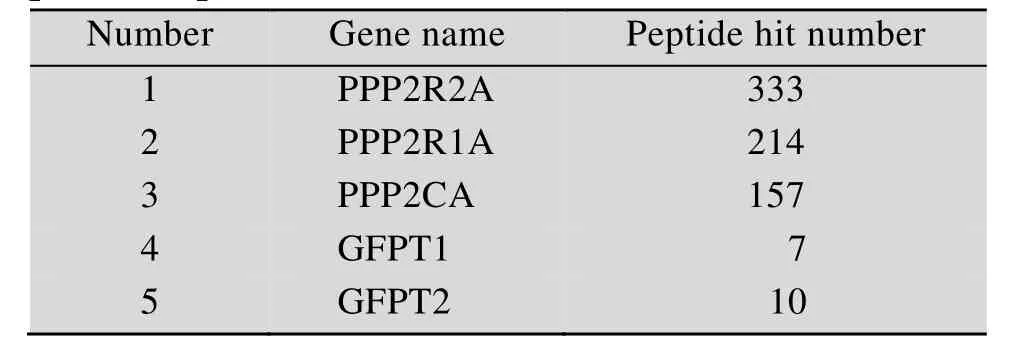
表2 串联亲和法联合 HPLC-Chip-ESI/MS/MS结果显示GFPT1/2是PPP2R2A潜在的结合蛋白Table 2 The results of tandem affinity and HPLCChip-ESI/MS/MS identified the GFPT1/2 as the potential partner of PPP2R2A
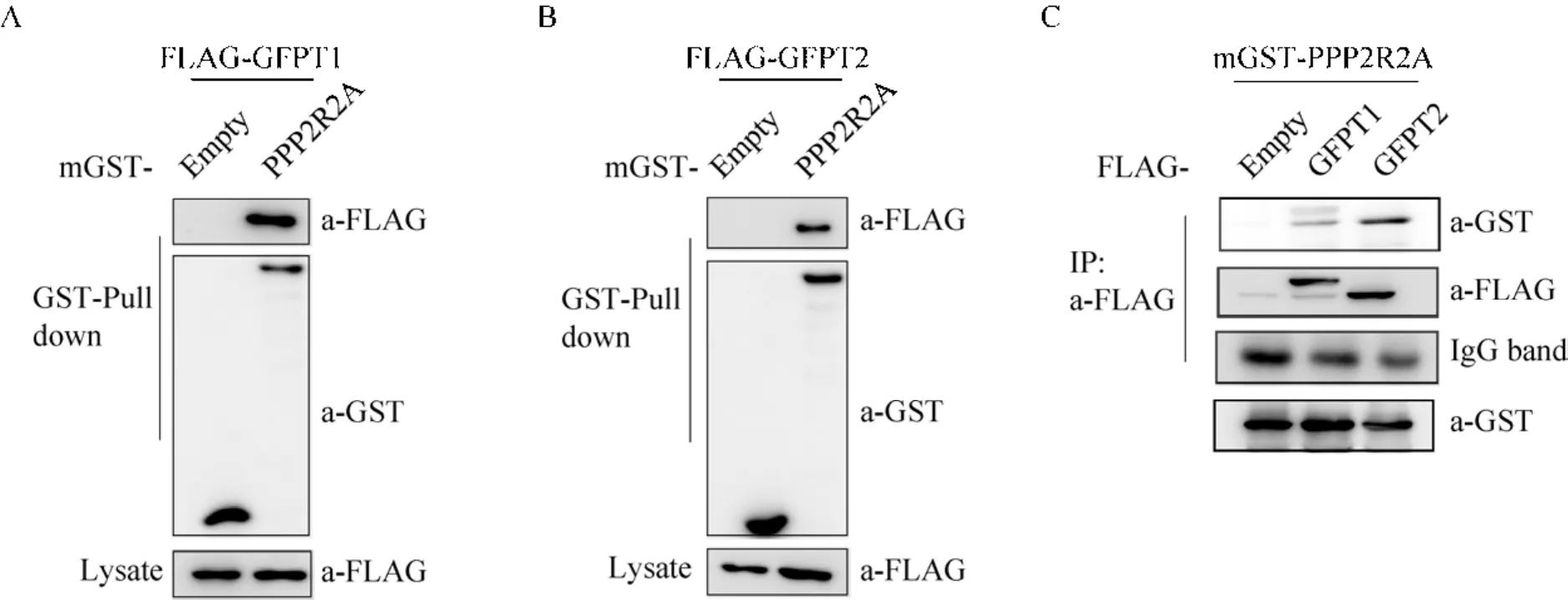
图2 PPP2R2A蛋白与GFPT1/2蛋白相互作用Fig. 2 The interaction between PPP2R2A and GFPT1/2 was further validated. (A, B) GST Pull-down of PPP2R2A with Flag-GFPT1/2. HEK-293T cells were co-transfected with plasmids expressing GST-tagged PPP2R2A (or empty vector) and Flag-tagged GFPT-1/-2. PPP2R2A was precipitated with GST Pull-down, and the pull-down products were analyzed by anti-Flag immunoblotting. (C) Co-immunoprecipitation (IP) of PPP2R2A with Flag-tagged GFPT1/2.PPP2R2A was immunoprecipitated with an anti-Flag antibody, and co-immunoprecipitated PPP2R2A was analyzed by anti-GST immunoblotting.
2.3 PPP2R2A蛋白与GFPT1/2蛋白在细胞内共定位
在HEK 293细胞中,共同转染HA-PPP2R2A和FLAG-GFPT1/2表达质粒,用鼠源抗FLAG和兔源抗HA抗体做免疫荧光检测,结果如图3A、B显示,PPP2R2A与GFPT1/2在细胞中共定位于细胞质中。
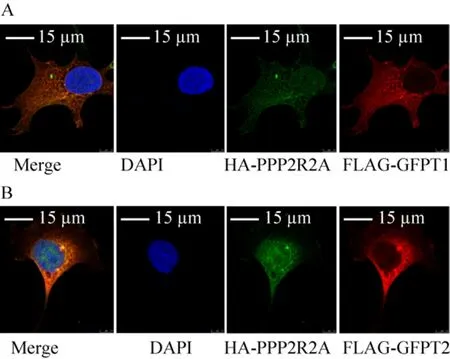
图3 PPP2R2A与GFPT1/2在细胞质中共定位的免疫荧光结果Fig. 3 Colocalization of PPP2R2A and GFPT1/2 in cytoplasm. (A, B) GFPT1/2 was co-localized with PPP2R2A in the cytoplasm. The HEK293 cells were co-transfected with plasmids expressing FLAG-GFPT1/2 and HA-PPP2R2A, and after 24 h, cells were fixed and stained with anti-HA (green) and anti-FLAG (red)antibody. And the DAPI were used for nuclear staining.
2.4 PPP2R2A影响GFPT2的磷酸化水平
已有的报道显示PKA可导致GFPT1/2的磷酸化,为了检测PPP2R2A去磷酸酶对GFPT1/2磷酸化的影响,我们通过慢病毒感染并表达shRNA,建立PPP2R2A稳定下调的 HEK 293T细胞,Western blotting结果显示该shRNA有效下调PPP2R2A表达。再在该稳定细胞株中转染 mGST-GFPT1/2表达质粒,用GST-pull down将外源性表达的GST-GFPT1/2进行纯化,Anti-GST-WB检测结果显示在PPP2R2A稳定下调细胞和对照细胞中,GST-GFPT1和GST-GFPT2的表达量相同,同时用 PKA底物抗体检测其磷酸化水平,结果显示 PPP2R2A下调后,GFPT1磷酸化水平改变不明显,GFPT2磷酸化水平增加 (图4 A、B)。另外,在FLAG-GFPT2稳定表达的MDA-MB-231细胞株中使用shRNA下调PPP2R2A表达,通过 Anti-FLAG-IP沉淀 FLAGGFPT2蛋白,Anti-PKA substrate WB结果同样显示,PPP2R2A的下调可促进 GFPT2的磷酸化,与HEK293T中的结果一致 (图4 C)。
2.5 在 MDA-MB-231中下调 PPP2R2A促进O-连接的蛋白质糖基化修饰
己糖胺途径是一系列的酶促生化反应,最终生成UDP-GlcNAC,它是蛋白质的O-连接的丝氨酸-苏氨酸的糖基化修饰以及糖原、糖脂修饰的底物,GFPT2是己糖胺途径中的关键限速酶,于是我们又进一步检测 PPP2R2A下调对细胞内总蛋白S/T-O-GlcNAC糖基化的影响,使用O-GlcNAC抗体 (CTD110.6) 进行Western blotting检测,结果 (图4) 表明下调PPP2R2A后,乳腺癌细胞中蛋白质的 S/T-O-GlcNAC糖基化修饰的水平显著增加。
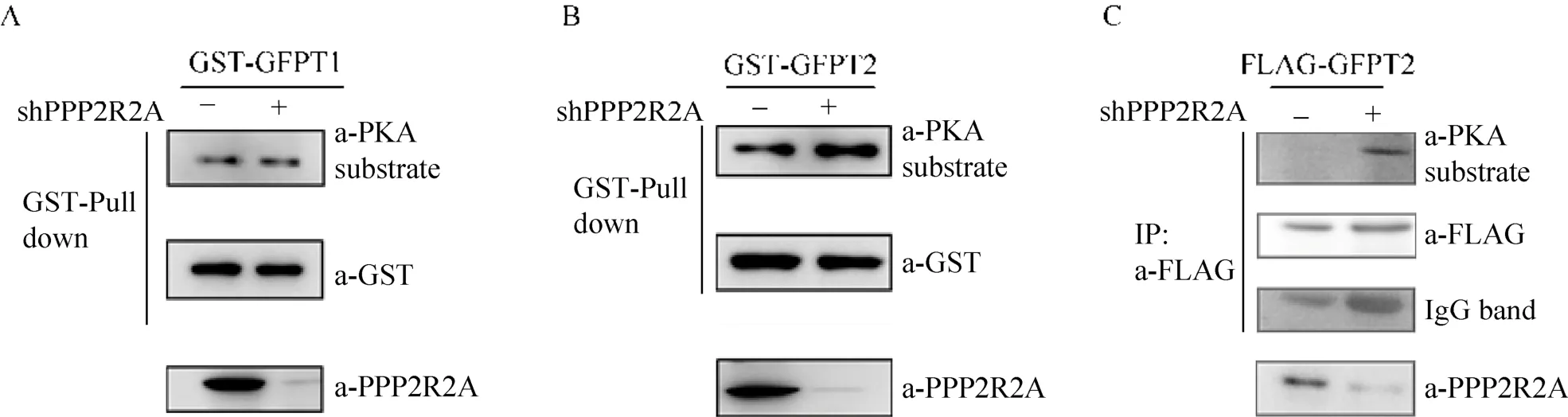
图4 PPP2R2A影响GFPT2的磷酸化水平Fig. 4 Knockdown of PPP2R2A enhanced phosphorylation of GFPT2. (A, B) Knockdown of PPP2R2A by lentivirus-mediated shRNA enhanced the phosphorylation of GFPT2, while the phosphorylation of GFPT1 had no significant change In HEK 293T. The plasmid of GST-GFPT1 or GFPT2 was further transfected into the normal and PPP2R2A-knockdown cells. GST Pull-down was used to precipitated the GFPT1 or GFPT2, and the phosphorylation was detected by the anti-PKA substrate immunoblotting. (C) In MDA-MB-231, knockdown of PPP2R2A enhanced the phosphorylation of GFPT2. MDA-MB-231 cells stably expressing FLAG-GFPT2 were established by infection with lentiviral particles. FLAG-GFPT2 was immunoprecipitated with an anti-FLAG antibody, and phosphorylation of GFPT2 was analyzed by the anti-PKA substrate immunoblotting.
3 讨论
PP2A的活性和特异性是通过调节亚基调控的,PPP2R2A作为PP2A复合物的调节亚基之一,通过底物特异性结合从而影响其磷酸化水平[12-13]。已有的研究表明,肿瘤细胞中谷氨酰胺的缺乏会促进PPP2R2A的表达,进而通过抑制P53通路提高肿瘤细胞在此环境中的生存[9]。为了更加深入地解释PPP2R2A的分子作用机制,我们进一步寻找新的与PPP2R2A结合的其他蛋白。
本研究通过慢病毒感染建立 FLAG-SBP-HISPPP2R2A稳定表达 MDA-MB-231细胞株,并首次使用 SBP-HIS串联亲和纯化联合质谱的方式对PPP2R2A的结合蛋白进行了分析。与文献中常使用的其他串联标签纯化 (Tandem Tag Purification,TAP) 方法相比,该方式具有以下3个优点[14-16]:1) 通过慢病毒感染建立稳定表达细胞,PPP2R2A表达量与内源性PPP2R2A表达量大致相当,更好地模拟 PPP2R2A内源性表达量;2) SBP-tag与Streptavidin-beads具有极高的亲和力,再经过HIS8-tag纯化的方法后,进一步增加了PPP2R2A复合物的纯度[17];3) 第二步HIS-tag的纯化不会引入其他过量的外源蛋白 (例如使用抗体时的免疫球蛋白或Streptavidin-beads上的Streptavidin),从而消除了过量纯化基质蛋白引入对质谱分析的影响。
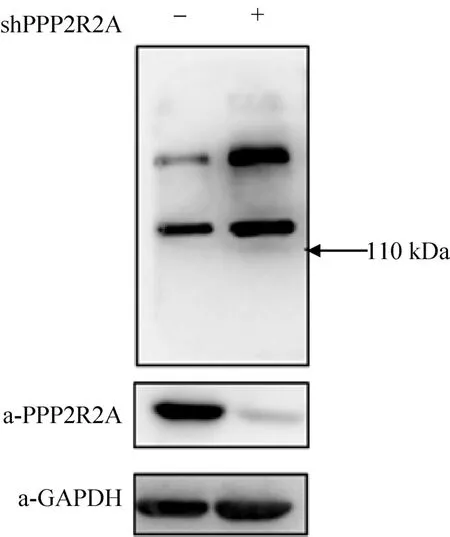
图5 PPP2R2A下调促进S/T-O-GlcNAC糖基化修饰Fig. 5 Cellular O-GlcNAcylation was elevated when PPP2R2A was knocked down in MDA-MB-231 cells.The PPP2R2A were knocked down by using the lentivirus-mediated shRNA, and the cellular O-GlcNAcylation was analyzed by anti-CTD110.6 immunoblotting.
通过该方法,我们鉴定出多个PPP2R2A已知结合蛋白及许多潜在的相互作用蛋白,其中包括GFPT1和GFPT2。通过GST Pull-down、Co-IP以及免疫荧光,我们进一步确定了PPP2R2A的确能够与GFPT1/2结合。人源GFPT1和GFPT2有76%的同源性[18],自身酶的活性受磷酸化水平的调控[19]。例如,PKA可通过磷酸化GFPT1的205S (丝氨酸) 而抑制其活性,PKA也可磷酸化GFPT2的202S (丝氨酸) 促进其活性[20-21]。我们的结果显示PPP2R2A下调可增加GFPT2的PKA磷酸化水平。
GFPT1和GFPT2是己糖胺途径的限速酶,催化谷氨酰胺水解为谷氨酸,使 6-磷酸-果糖转化为 6-磷酸-葡萄糖胺。然后经一系列反应产生UDP-GlcNAC,它是蛋白质O-连接的丝氨酸-苏氨酸的糖基化修饰以及糖原、糖脂修饰的底物。参与细胞内的多条信号通路的调控[22-24]。我们的结果表明PPP2R2A下调后促进GFPT2磷酸化水平的同时,也会促进细胞内总O-GlcNAC糖基化修饰的增加。
在本研究中,我们还通过慢病毒感染在MDA-MB-231细胞中上调了PPP2R2A的表达,但未检测到GFPT2和O-GlcNAC糖基化修饰的改变,推测这是由于细胞本身就有很高的内源性PPP2R2A蛋白表达。例如,在图 1中内源性PPP2R2A蛋白与慢病毒感染表达的 FLAG-HISPPP2R2A蛋白量很近似;在图4C中PPP2R2A未下调的 MDA-MB-231细胞中基本检测不到GFPT2的磷酸化状态,这说明在细胞中具有高活性的PPP2R2A;同时我们也注意到,最近发表的一篇关于PPP2R2A去磷酸化EDD并调控P53的报道中[9],作者同样仅开展了 PPP2R2A的下调研究,且在对照组中 EDD磷酸化水平也是几乎检测不到。这些均说明在细胞中具有很高的PPP2R2A活性,也提示PPP2R2A具有作为治疗靶标的潜力。
另外,在细胞中大约有 80多种蛋白发生O-GlcNAC糖基化修饰[25],在接下来的研究中我们将进一步鉴定,哪种关键的下游蛋白 O-GlcNAC糖基化修饰的改变介导了 PPP2R2A下调引起的生物学效应。
REFERENCES
[1]Janssens V, Goris J. Protein phosphatase 2A: a highly regulated family of serine/threonine phosphatases implicated in cell growth and signalling. Biochem J,2001, 353(3): 417–439.
[2]Eichhorn PJA, Creyghton MP, BERNARDS R. Protein phosphatase 2A regulatory subunits and cancer. Biochim Biophys Acta (BBA)-Rev Cancer, 2009, 1795(1): 1–15.
[3]Seshacharyulu P, Pandey P, Datta K, et al. Phosphatase:PP2A structural importance, regulation and its aberrant expression in cancer. Cancer Lett, 2013, 335(1): 9–18.
[4]Gilan O, Diesch J, Amalia M, et al. PR55α-containing protein phosphatase 2A complexes promote cancer cell migration and invasion through regulation of AP-1 transcriptional activity. Oncogene, 2015, 34(10):1333–1339.
[5]Mori N, Ishikawa C, Uchihara JN, et al. Protein phosphatase 2A as a potential target for treatment of adult T cell leukemia. Curr Cancer Drug Targ, 2013,13(8): 829–842.
[6]Beca F, Pereira M, Cameselle-Teijeiro JF, et al. Altered PPP2R2A and Cyclin D1 expression defines a subgroup of aggressive luminal-like breast cancer. BMC Cancer,2015, 15(1): 285.
[7]Suganuma M, Fujiki H. Tumor promotion by inhibitors of protein phosphatase 1 and 2A. Tanpakushitsu Kakusan Koso, 1998, 43(S8): 1102–1110.
[8]Hein AL, Seshacharyulu P, Rachagani S, et al. PR55α subunit of protein phosphatase 2A supports the tumorigenic and metastatic potential of pancreatic cancer cells by sustaining hyperactive oncogenic signaling. Cancer Res, 2016, 76(8): 2243–2253.
[9]Reid MA, Wang WI, Rosales KR, et al. The B55α subunit of PP2A drives a p53-dependent metabolic adaptation to glutamine deprivation. Mol Cell, 2013,50(2): 200–211.
[10]Wellen KE, Lu C, Mancuso A, et al. The hexosamine biosynthetic pathway couples growth factor-induced glutamine uptake to glucose metabolism. Genes Dev,2010, 24(24): 2784–2799.
[11]Ferrer CM, Sodi VL, Reginato MJ. O-GlcNAcylation in cancer biology: linking metabolism and signaling. J Mol Biol, 2016, 428(16): 3282–3294.
[12]Ory S, Zhou M, Conrads TP, et al. Protein phosphatase 2A positively regulates Ras signaling by dephosphorylating KSR1 and Raf-1 on critical 14-3-3 binding sites. Curr Biol, 2003, 13(16): 1356–1364.
[13]Ruvolo PP. The broken “off” switch in cancer signaling:PP2A as a regulator of tumorigenesis, drug resistance,and immune surveillance. BBA Clin, 2016, 6: 87–99.
[14]Li Y. The tandem affinity purification technology: an overview. Biotechnol Lett, 2011, 33(8): 1487–1499.
[15]Kosobokova EN, Skrypnik KA, Kosorukov VS.Overview of fusion tags for recombinant proteins.Biochemistry (Moscow), 2016, 81(3): 187–200.
[16]Ma Z, Fung V, D’orso I. Tandem affinity purification of protein complexes from eukaryotic cells. J Vis Exp,2017(119), doi: 10.3791/55236.
[17]Loughran ST, Walls D. Tagging recombinant proteins to enhance solubility and aid purification//Walls D,Loughran S, Eds. Protein Chromatography: Methods and Protocols. New York: Humana Press, 2017,1485:131–156.
[18]Oki T, Yamazaki K, Kuromitsu J, et al. cDNA cloning and mapping of a novel subtype of glutamine:fructose-6-phosphate amidotransferase (GFAT2) in human and mouse. Genomics, 1999, 57(2): 227–234.
[19]Yamazaki K. Glutamine–fructose-6-phosphate transaminase 1, 2 (GFPT1, 2)//Taniguchi N, Honke K, Fukuda M, et al. Handbook of Glycosyltransferases and Related Genes.Tokyo: Springer, 2014: 1465–1479.
[20]Hu Y, Riesland L, Paterson AJ, et al. Phosphorylation of mouse glutamine-fructose-6-phosphate amidotransferase 2 (GFAT2) by cAMP-dependent protein kinase increases the enzyme activity. J Biolog Chem, 2004, 279(29):29988–29993.
[21]Chang Q, Su KH, Baker JR, et al. Phosphorylation of human glutamine: fructose-6-phosphate amidotransferase by cAMP-dependent protein kinase at serine 205 blocks the enzyme activity. J Biolog Chem, 2000, 275(29):21981–21987.
[22]Ruan HB, Singh JP, Li MD, et al. Cracking the O-GlcNAc code in metabolism. Trends Endocrinol Metabol, 2013, 24(6): 301–309.
[23]Slawson C, Hart GW. Dynamic interplay between O-GlcNAc and O-phosphate: the sweet side of protein regulation. Curr Opin Struct Biol, 2003, 13(5): 631–636.
[24]Comer FI, Hart GW. O-GlcNAc and the control of gene expression. Biochim Biophys Acta (BBA)-Gen Subj,1999, 1473(1): 161–171.
[25]Wells L, Whelan SA, Hart GW. O-GlcNAc: a regulatory post-translational modification. Biochem Biophys Res Commun, 2003, 302(3): 435–441.
