拉枝角度对苹果枝条顶梢多胺类物质含量的影响
2018-05-08吕珍珍张满让
吕珍珍,马 玲,张满让
(西北农林科技大学 园艺学院,陕西 杨凌 712100)
多胺(PAs)是一类具有生物活性的低分子脂肪族含氮碱,是近年来研究较多的一类新型植物生长发育调节物质,以多种形式广泛存在于植物体内,通常分为游离态、共轭态和束缚态3种,主要分布在分生组织中,其中最常见研究也较多的属游离态多胺中的腐胺(Put)、亚精胺(Spd)和精胺(Spm)[1-2],它们与植物成花、坐果、生长、休眠、衰老以及抗逆性等生理活动密切相关。近年来,许多研究表明,多胺在园艺作物花芽分化与花粉形成过程中发挥着重要作用[3-4],参与花诱导与花器官形成[5-8]。Sood和Nagar[9]发现,玫瑰开花过程中多胺类物质含量会发生变化,而且变化趋势与其存在形态有关。在果树方面,一些研究者认为多胺可以促进果树花芽分化,提高成花率,且其含量随花芽分化进程的推进而改变[10-11]。
研究发现,拉枝可抑制苹果树冠扩张生长,提高果实产量,改善果实品质,同时可以打破顶端优势,改变树体激素分配格局,对苹果成花可发挥良好的促进效果[12]。近年来,欧洲苹果栽培普遍推行高纺锤形果树,即将枝条角度扩展到95°~120°,其优点是易成花,树体容易控制,因此国内外学者将研究重点集中放在加大拉枝角度、促进苹果成花结果上。而关于拉枝,前人研究较多的则是其对树形构建、果树生理特性、果实品质以及内源激素等物质的影响[13-16],有关拉枝对多胺影响方面的甚少涉及,研究也不够深入。因此,本研究将拉枝与多胺类物质相联系,通过对富士和嘎拉苹果树枝条进行不同角度的拉枝处理,研究在花芽分化期内,短枝和中长枝顶芽内多胺类物质含量的变化规律,旨在为拉枝技术在促进苹果成花方面的广泛应用提供理论依据。
1 材料与方法
1.1 材 料
试验在西北农林科技大学千阳苹果试验站进行。此园建于2012年,以富士为主栽品种,嘎拉为授粉树,基砧为八棱海棠,M26为矮化中间砧,树形采用高效纺锤形,株行距1.3 m×4.0 m, 树势健壮均一,有灌溉条件,管理精细。
多胺标准品腐胺(Put)、亚精胺(Spd)和精胺(Spm)均购自Sigma公司,高氯酸、苯甲酰氯、乙醚、氢氧化钠、氯化钠为国产分析纯,甲醇为Tedia色谱级甲醇,试验用水为Mili-Q超纯水。
1.2 试验设计
于2015年4月上旬对富士、嘎拉进行拉枝处理,分别选取生长健壮、树势一致的富士和嘎拉苹果树各120棵,各分成3个小区,每小区40株树,小区间重复随机排列;每小区进行4种拉枝处理,角度分别是0°(即CK,直立生长枝条),70°,90°和110°。枝条选取方法:在每株树距离地面 1.0~2.0 m位置,选择相同方位、粗度和长度相近且呈自然生长角度的1~2年生主枝全部进行拉枝处理,其余枝条不作处理。分别于2015年5月20日、5月31日、6月10日、6月20日、7月20日、8月20日、9月20日进行采样,每次采样分别在每区每处理的1棵树上进行,3个重复,分别采取短枝(春梢且长度小于5 cm)和中长枝(长度5 cm 以上)的饱满顶芽,用锡箔纸包好放入冰盒或液氮中带回实验室,保存在超低温冰箱中备用。
1.3 成花率统计
2015年4月上旬,分别在每区每处理未采样树体上标记不同方位、长势基本一致的当年生枝条30个,于2016年春季盛花期分别统计短枝顶芽与中长枝顶芽的成花率(成花率=成花枝数/试验总枝数×100%),其中每处理随机统计15个主枝作为1个重复,共3个重复。
1.4 多胺类物质含量的测定
采用高效液相色谱法(HPLC)测定Put、Spd、Spm、多胺(PAs)含量,参照刘俊等[17]的方法并加以改进。具体方法为:取植物材料0.5 g,加入5 mL预冷的体积分数5%高氯酸冰浴研磨,冰浴浸提1 h后离心(12 000×g30 min,4 ℃)。取500 μL上清液至10 mL带盖塑料离心管中,加入7 μL苯甲酰氯,再加入1 mL 2 mol/L NaOH溶液,涡漩20 s后于37 ℃水浴反应20 min。然后在反应液中加入2 mL饱和NaCl溶液,混匀后用2 mL乙醚萃取,1 500×g离心5 min,取1 mL醚相真空干燥,用100 μL甲醇涡旋溶解后过0.45 μm滤膜,取10 μL进样。色谱条件:检测器Waters2489,柱子Novapak C18反相柱(Waters,150 mm×3.9 mm,4 μm),柱温30 ℃,流速1 mL/min,检测波长254 nm,流动相为V(甲醇)∶V(水)=60∶40,数据处理采用Breeze色谱工作站软件完成。
1.5 数据分析
数据经Excel 2007整理后,利用SPSS 17.0软件对其进行方差分析和显著性检验。
2 结果与分析
2.1 不同拉枝角度对富士和嘎拉苹果枝条顶芽成花率的影响
从图1可以看出,拉枝可明显提高苹果成花率,富士苹果随着拉枝角度的增大,短枝顶芽的成花率逐步增加,110°时最高,且各处理间差异显著;而中长枝顶芽成花率则较低,差异不明显。嘎拉苹果在 90°与 110°拉枝角度下,短枝顶芽成花率差异不显著,但与拉枝70°相比显著提高,这说明嘎拉苹果在拉枝 90°时便可满足成花结果的要求。此外,对照处理所有中长枝成花率均为0,短枝成花率也极低。整体而言,嘎拉苹果平均成花率略高于富士,这说明拉枝对嘎拉的作用效果更明显;与嘎拉相比,富士对拉枝要求更高。
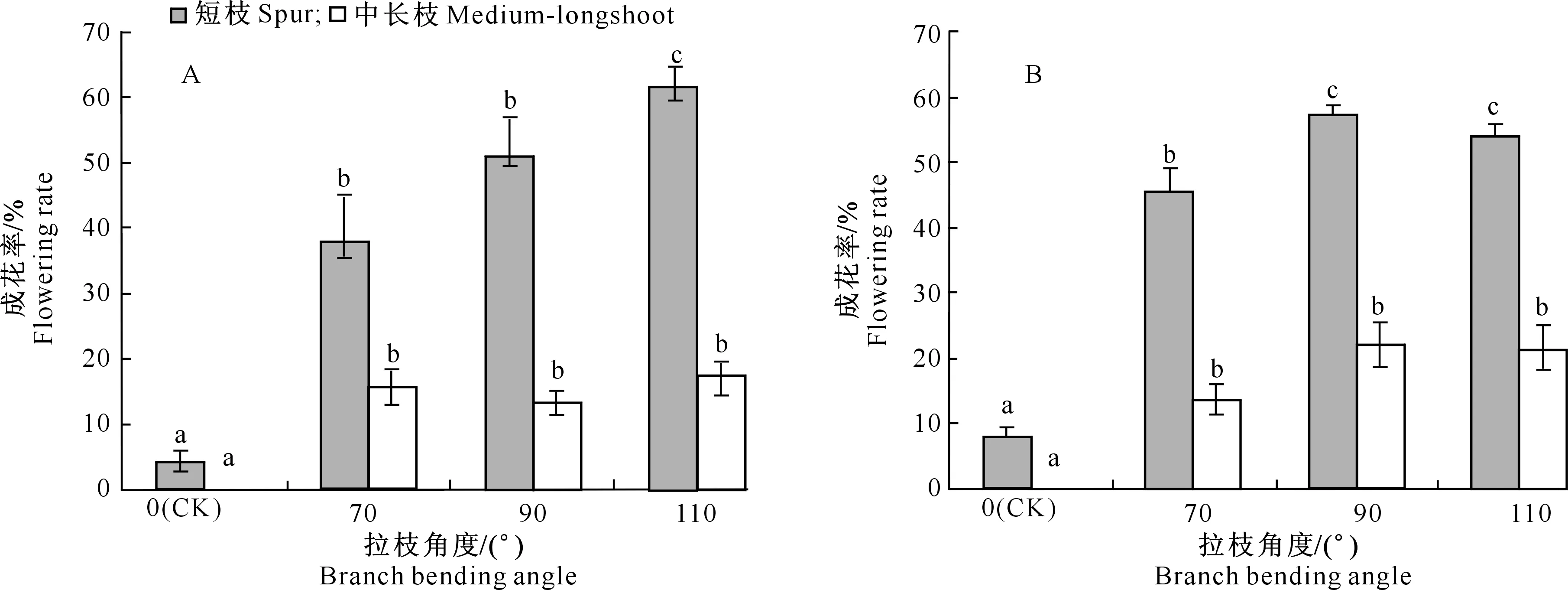
图柱上标不同字母表示同类枝条不同处理差异达到显著水平(P<0.05) Different letters indicate significant difference between treatments at P<0.05图1 拉枝角度对富士(A)和嘎拉(B)枝条顶芽成花率的影响Fig.1 Effect of branch bending angle on flowering rate of terminal buds in Fuji(A) and Gala(B)
2.2 不同拉枝角度下富士苹果枝条顶芽多胺类物质含量的动态变化
在富士苹果整个花芽分化期内,不同枝条顶芽内多胺类物质含量呈现一定的变化规律,3种物质整体均表现为上升趋势,且6月20日至7月20日总体急剧升高,7、8月份含量较高,后期又开始下降。在Put、Spd、Spm 3种物质中,Spm含量远高于前两者;从3种物质总含量看,短枝与中长枝差别不大,但是不同处理间存在差异,拉枝110°处理3类物质含量高于对照与另外2种角度拉枝处理(图2~5)。

同一时期所标字母从上至下依次对应110°,90°,70°和0°(CK)处理,不同字母表示处理间差异达到显著水平(P<0.05)。下图同 The letters at same period from top to bottom correspond to110°,90°,70°and 0°(CK),respectively,different letters indicate significant difference between treatments at P<0.05.The same below图2 不同拉枝角度下富士枝条顶端Put含量的变化Fig.2 Changes of Put content in shoot terminal of Fuji under different branch bending angles
2.2.1 Put含量 由图2可知,富士苹果在整个花芽分化期内,无论是何种拉枝角度处理,不同枝条顶芽Put含量均处于上升趋势,而且短枝中Put含量高于中长枝;前期Put含量上升幅度不大,相对比较稳定,6月20日之后急剧升高,7月20日之后开始下降,且后期波动较大。整体而言,富士苹果在6月20日之后,拉枝110°处理Put含量相对较高,而对照中长枝即直立枝条Put含量变化不大;同一角度处理下,不同生长时期短枝与中长枝顶端Put含量均在7月20日出现峰值,其中拉枝110°下Put含量分别为91.996 8和70.865 2 nmol/g。
2.2.2 Spd含量 图3显示,富士苹果在整个花芽分化期内,无论是何种拉枝角度处理,Spd含量均远高于Put含量。在整个生长时期,Spd含量变化规律与Put较为相似,即在前期阶段,Spd含量相对比较稳定,6月20日至7月20日急剧升高,7月20日之后变化较为缓慢;7月20日以后,短枝中除了拉枝110°处理表现为下降外,其余3种处理仍呈上升趋势,最终趋于稳定,而中长枝所有处理Spd含量均开始下降。其中110°的短枝以及所有中长枝Spd含量均在7月20日出现最大值。短枝与中长枝在拉枝110°下的Spd含量峰值分别为689.958 0和621.773 3 nmol/g。

图3 不同拉枝角度下富士枝条顶端Spd含量的变化Fig.3 Changes of Spd content in shoot terminal of Fuji under different branch bending angles
2.2.3 Spm含量 图4表明,富士苹果在整个花芽分化期内,无论是何种拉枝角度处理,前期Spm含量均低于后期;与Put、Spd相比,Spm含量最高,且变化趋势较为平缓。总体而言,短枝与中长枝中Spm含量差别不大,而且均在8月20日出现峰值,峰值出现时间滞后于Put和Spd;其中在拉枝110°下,短枝与中长枝的Spm含量峰值分别为1 477.771 1和1 492.464 8 nmol/g。
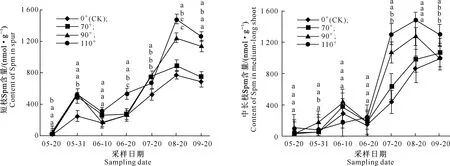
图4 不同拉枝角度下富士枝条顶端Spm含量的变化Fig.4 Changes of Spm content in shoot terminal of Fuji under different branch bending angles
2.2.4 PAs含量 由图5可知,富士不同枝条顶端PAs含量在整个生长阶段总体上升,且均在8月20日出现峰值,随后开始下降。后期PAs含量远大于前期,短枝与中长枝PAs含量差别不大。在拉枝初期,不同处理PAs含量差异不大;随着拉枝时间的延长,不同角度拉枝处理间的差异也越来越大,并且在后期阶段PAs含量随着拉枝角度的增大依次升高。4种拉枝角度中,以拉枝110°处理PAs含量最高,9月20日时其与对照和拉枝70°处理有显著差异。
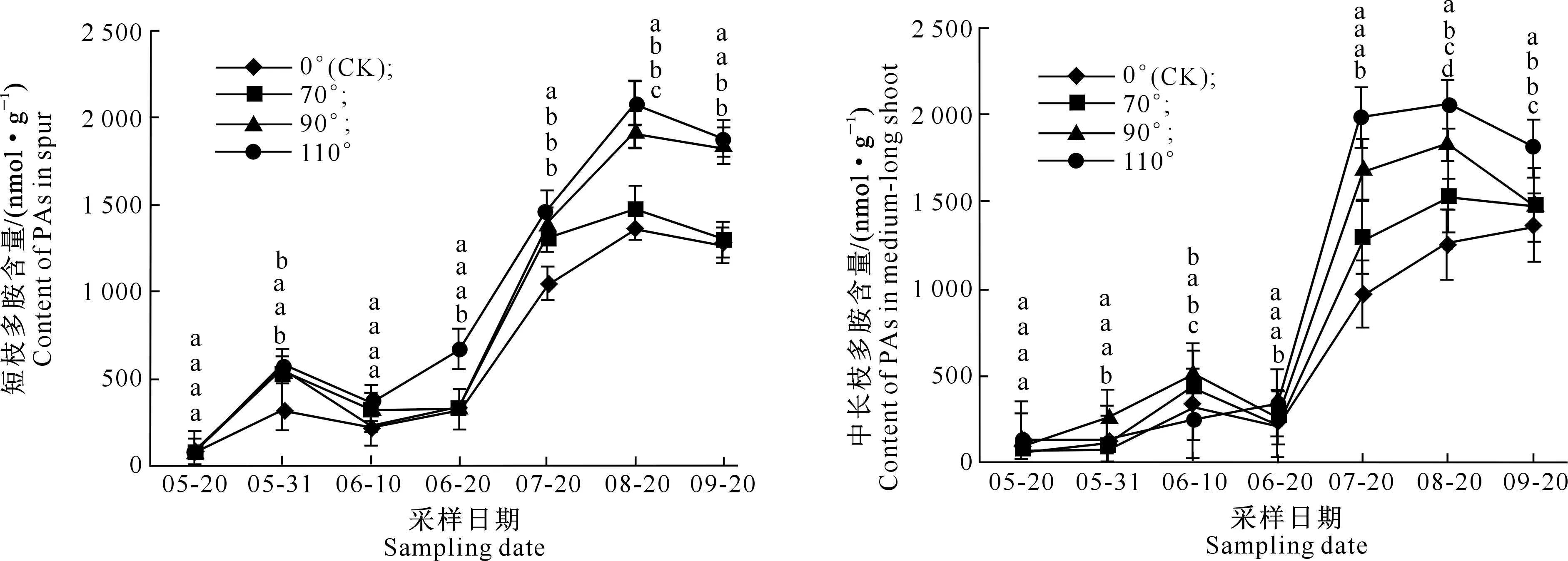
图5 不同拉枝角度下富士枝条顶端PAs含量的变化Fig.5 Changes of PAs content in shoot terminal of Fuji under different branch bending angles
2.3 不同拉枝角度下嘎拉苹果枝条顶芽多胺类物质含量的动态变化
在嘎拉苹果整个花芽分化期内,多胺类物质含量均高于富士品种,变化规律与其相类似,即整个阶段表现为上升趋势,6月20日至7月20日变化剧烈,7、8月份含量较高,后期末开始下降;与富士不同的是嘎拉短枝与中长枝PAs含量之间差异比较明显。另外,总体上看拉枝70°,90°,110°枝条顶芽多胺类物质含量总体均高于对照,且以拉枝90°处理最高。在Put、Spd、Spm 3种物质中,Spm含量远高于前两者,且Put和Spm均有2个相对较为明显的峰值(图6~9)。
2.3.1 Put含量 如图6所示,嘎拉不同枝条顶芽的Put含量高于富士,变化趋势基本相似,即前期低于后期。与富士相比,嘎拉前期阶段Put含量波动较大,总体先升后降,5月31日出现第1个峰值,之后先降后升,在7月20日出现第2个峰值,7月20日之后所有短枝Put含量开始下降,而拉枝70°,90°与110°处理中长枝则表现为先降后升。此外,在所有处理中,拉枝90°处理枝条顶芽Put含量高于其他处理,其短枝和中长枝顶芽内的Put含量在7月20日峰值分别为102.906 0和49.848 4 nmol/g。
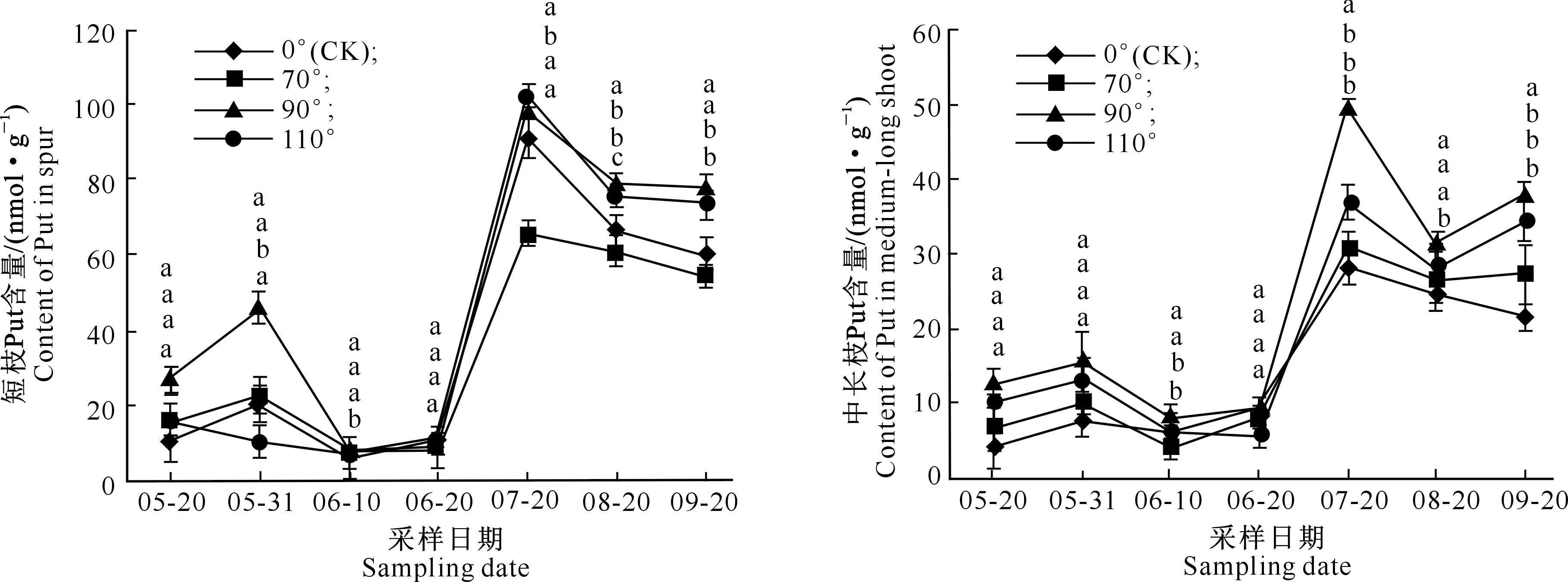
图6 不同拉枝角度下嘎拉枝条顶端Put含量的变化Fig.6 Changes of Put content in shoot terminal of Gala under different branch bending angles
2.3.2 Spd含量 由图7可知,嘎拉不同枝条顶芽的Spd含量高于富士,整体变化趋势基本相似,即前期低于后期。短枝中除了对照,其他3种处理枝条顶芽内Spd含量在整个采样期一直处于上升趋势。中长枝中,拉枝70°与110°处理的Spd含量在7月20日出现峰值,之后Spd含量总体都呈下降趋势;对照Spd含量在6月20日之前较低,之后则表现为升高;而拉枝90°处理Spd含量在8月20日达到峰值,之后降低。在采样末期(8月20日-9月20日),所有处理中Spd含量最高的均是拉枝90°处理,该处理下短枝最大值出现在9月20日,为843.556 7 nmol/g;而该处理下中长枝最大值出现在8月20日,为779.266 0 nmol/g。

图7 不同拉枝角度下嘎拉枝条顶端Spd含量的变化Fig.7 Changes of Spd content in shoot terminal of Gala under different branch bending angles
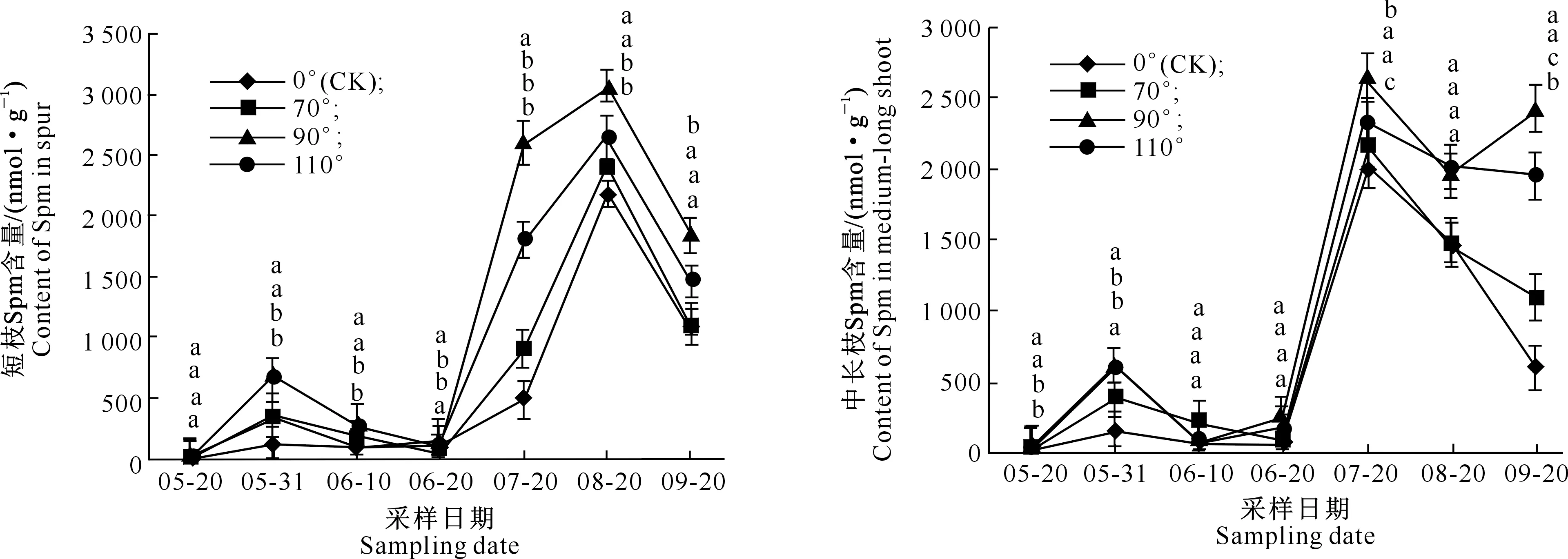
图8 不同拉枝角度下嘎拉枝条顶端Spm含量的变化Fig.8 Changes of Spm content in shoot terminal of Gala under different branch bending angles

图9 不同拉枝角度下嘎拉枝条顶端PAs含量的变化Fig.9 Changes of PAs content in shoot terminal of Gala under different branch bending angles
2.3.3 Spm含量 由图8可知,嘎拉不同枝条顶芽的Spm含量高于富士,且前期含量低于后期。
前期即6月20日之前,短枝与中长枝均在5月31 日出现第1个峰值;后期短枝与中长枝分别在8月20日、7月20日出现第2个峰值,其中拉枝90°处理短枝与中长枝Spm含量峰值明显高于其他处理,分别为3 081.924 1和2 657.238 4 nmol/g。短枝不同处理Spm含量在6月20日至8月20日间差异较大,而中长枝各处理间差异在7月20日至9月20日较大(图8)。
2.3.4 PAs含量 图9显示,嘎拉不同枝条顶芽PAs含量明显高于富士,而且在整个生长阶段总体表现为上升趋势,后期远大于前期,且短枝高于中长枝,均出现2个峰值,其中短枝和中长枝第1个峰值均出现在5月31日,而第2个峰值分别出现在8月20日和7月20日。4种拉枝角度中以90°处理PAs含量最高,与70°和110°处理差异显著。相比于中长枝,中后期短枝不同处理间的差异更为明显,而且拉枝时间越久,差异越大。
3 讨 论
苹果花芽分化是一个十分复杂的过程,传统意义上将该过程分为2个时期,即生理分化期与形态分化期,而实际上两者之间并不存在清楚的界限,而是在时间上有重叠交叉现象。研究表明,多胺对苹果的花芽分化起着重要作用[10,18-20]。在生产实践中人们发现,富士与嘎拉2个苹果品种的成花能力差异较大,在本研究采样过程中笔者也观察到,嘎拉顶芽明显大于富士,且更为饱满。同时,本试验结果显示,供试芽中内源多胺含量表现为嘎拉远高于富士,可见内源多胺含量高低与苹果的成花能力密切相关,因此在生产中可采取喷施外源多胺的方法来提高一些成花较难的苹果品种的成花率,从而达到提高苹果产量的目的。此外,本试验还发现,在2个苹果品种的花芽分化过程中,短枝顶芽内多胺类物质含量高于中长枝,说明苹果树不同组织部位的多胺类物质含量有差异。
曾骧等[21]发现,在金冠苹果花原基大量分化时,芽内精氨酸含量及其在总氨基酸中的比例明显上升,而精氨酸是多胺合成的重要前体物质,说明多胺类物质与苹果花芽分化过程之间存在重要关联。本研究结果显示,在5月20日至6月20日即生理分化期内,多胺类物质含量相对较低,这可能是因为它们在成花诱导过程中参与了蛋白质与核酸的形成。但3种多胺类物质含量变化具有一定规律,其中Put与Spm含量在该时期整体表现为先升后降,5月31日前后出现高峰,而Spd含量变化平缓,说明在苹果花芽分化中,Put和Spm协同诱导花芽分化,起到关键作用。而到采样后期,3种多胺类物质含量均开始升高,则说明苹果花芽在形态分化时期需要大量多胺参与花器官的形成,进而完成花芽分化过程,尤其是Spm。以上结果表明,3种多胺类物质在苹果花芽分化过程中发挥作用的具体时期有所不同。而徐继忠等[20]曾报道,在核桃雌花芽生理分化期,混合芽中的游离态Put、Spd和Spm含量升高,当形态分化开始时,这3种多胺类物质含量开始下降,可见在不同植物花芽分化中,多胺类物质含量变化有所差异,这可能与花芽形成过程以及分化时期不同有关。
近年来,随着人们对多胺研究的日益深入,关于高等植物的花芽分化与内源多胺的关系已有不少报道,而在不同植物花芽分化中,起调节作用的内源多胺类物质种类也有所不同。郑家基等[22]研究表明,内源Put、Spd增加有利于龙眼营养生长,而Spm增多则有利于花芽分化;陈鸿等[23]认为,内源多胺可促进新铁炮百合花芽成花,Put和Spd含量的增加有利于雌蕊原基形成,而 Spm增多则有利于雄蕊原基的形成;钟晓红等[24]发现,在奈李花芽孕育期间,芽内Put含量降低,而Spd和Spm含量出现高峰;Guo等[25]则认为,在菊花花芽分化中,Spd含量升高可能对花原基的发生与形成有利。本研究中,富士和嘎拉苹果枝条顶芽在整个采样时期,Put、Spd和Spm 3种多胺类物质的含量变化有差异,Spm含量最高,Spd次之,Put最低,这首先说明Spm在苹果花芽分化中的作用更直接、效果更显著,占主导地位;其次,由于Spm主要分布于细胞核中,高含量Spm可能是大量合成DNA所需要的,与成花基因DNA的启动和合成有关。根据多胺在植物中的合成途径,已知多胺的合成始于Put,Put是Spd、Spm合成过程中的直接底物,其含量高低直接影响后续的Spd和Spm含量[26],而在苹果花芽分化过程中,形态分化的进行有可能进一步加剧了3种物质间的转化,使得Spm含量远高于Put和Spd,并且随着花器官原始体的逐渐形成,其含量逐渐下降,可见Spm对苹果花器官的形成发挥着至关重要的作用。
目前,国内外学者关于拉枝等修剪方式对内源多胺含量影响的研究甚少。王世平等[27]发现,在对苹果树进行环切处理后,内源多胺类物质含量与成花作用显著相关,认为Put有助于分生组织的活化,而Spd和Spm与花诱导的启动及促进有关。徐东花等[28]研究证明,外源Spd可促进花芽分化阶段木质素的合成,说明在植物体内多胺类物质与木质素之间存在某种联系。这一结论证实了“拉枝通过影响枝条内木质素的合成与积累,进而间接致多胺类物质含量发生变化”的观点,用此观点可解释本试验中拉枝使多胺含量升高这一现象。除此之外,多胺含量升高也可能与逆境胁迫响应机制有关,即苹果树在拉枝胁迫下,形成响应机制,枝条内多胺发生积累,但这还有待进一步证实。本研究发现,拉枝可以明显提高苹果成花率,与此同时苹果枝条顶芽多胺含量也升高,且增加幅度与拉枝角度有关。与对照相比,富士苹果在110°拉枝处理下多胺类物质含量明显高于另外2种拉枝处理;而嘎拉苹果在拉枝90°处理下多胺含量更高,70°和110°处理下差异并不显著。随着拉枝时间的延长,2个苹果品种的多胺类物质含量持续增加,这可能是由于拉枝影响了此类物质的运输,使其在枝条顶端不断积累,也可能是花原基分化的需要,后期它们大量参与了花器官的形成;至于末期多胺类物质含量总体出现下降则可能是花芽进入休眠期所致,因为苹果花芽在进入休眠期前后,芽体内相关的营养物质及激素代谢调节变慢,甚至趋于停止。关于拉枝是否可以通过影响内源多胺含量进而调控相关基因表达,并最终影响花芽分化;或是内源多胺含量升高直接调控了基因表达,从而影响了花芽分化,还需要从分子水平方面进行深入地研究。
[参考文献]
[1] Buchanan B B,Gruissem W,Jones R L.Biochemistry and molecular biology in plants [M].Maryland:American Society of Plant Physiologists,2000:1-90.
[2] Martin T J.Metabolism and function of polyamines in plants:recent development (new approaches) [J].Plant Growth Regul,2001,34:135-148.
[3] Aribaud M,Martin T J.Polyamine metabolism,floral initiation and floral development in chrysanthemum (ChrysanthemummorifoliumRamat) [J].Plant Growth Regul,1994,15:23-31.
[4] Wimalasekera R,Tebarta F,Scherer G F E.Polyamines,polyamine oxidases and nitric oxide in development,abiotic and biotic stresses [J].Plant Sci,2011,181:593-603.
[5] Malmberg R L,Mcindod J,Hiatt A C,et al.Genetics of polyamine synthesis in tobacco:developmental switches in the flower [J].Cold Spring Harbor Symp Quant Biol,1985,50:475-482.
[6] Kaur S R,Tiburcio A F,Galston A W.Spermidine and flower-bud differentiation in thin-layer explants of tobacco [J].Planta,1988,173:282-284.
[7] Kakkar R K,Rai V K.Plant polyamines in flowering and fruit ripening [J].Phytochemistry,1993,33(6):1281-1288.
[8] Huang C K,Chang B S,Wang K C,et al.Changes in polyamine pattern are involved in floral initiation and development inPolianthestuberosa[J].J Plant Physiol,2004,161:709-713.
[9] Sood S,Nagar P K.Changes in endogenous polyamines during flower development in two diverse species of rose [J].Plant Growth Regulation,2004,44:117-123.
[10] 徐继忠,陈海江,邵建柱,等.外源多胺促进红富士苹果花芽形成的效应 [J].果树科学,1998,15(1):10-12.
Xu J Z,Chen H J,Shao J Z,et al.Effects of exogenous polyamines on flower bud formation of red Fuji apple variety [J].Journal of Fruit Science,1998,15(1):10-12.
[11] 李 璇.两个砂梨品种花芽分化期芽内内源多胺含量的动态变化 [J].中国南方果树,2013,42(2):26-29.
Li X.Dynamic changes of endogenous polyamines in the buds of ‘Shinsui’ and ‘Housui’ pear (Pyruspyrifolia) during the differentiation of floral bud [J].South China Fruits,2013,42(2):26-29.
[12] Lauri P E,Lespinasse J M.Genotype of apple tree affects growth and fruiting responses to shoot bending at various times of year [J].Journal of the American Society for Horticultural Science,2001,126(2):169-174.
[13] 韩明玉,李永武,范崇辉,等.拉枝角度对富士苹果树生理特性和果实品质的影响 [J].园艺学报,2008,35(9):1345-1350.
Han M Y,Li Y W,Fan C H,et al.Effects of branch bending angle on physiological characteristics and fruit quality of Fuji apple [J].Acta Horticulturae Sinica,2008,35(9):1345-1350.
[14] Zhang M R,Han M Y,Ma F W,et al.Effect of bending on the dynamic changes of endogenous hormones in shoot terminals of ‘Fuji’ and ‘Gala’ apple trees [J].Acta Physiol Plant,2015,37:76.
[15] 艾沙江·买买提,杨 清,王晶晶,等.短截、拉枝、刻芽对苹果枝条不同部位芽激素含量的影响 [J].园艺学报,2013,40(8):1437-1444.
Maimaiti A S J,Yang Q,Wang J J,et al.Effects of cutting back,branch-bending and bud-notching treatments on endogenous hormones in the buds of Fuji apple [J].Acta Horticulturae Sinica,2013,40(8):1437-1444.
[16] 李雪薇,李丙智,刘富庭,等.刻芽、扭枝和去顶梢对富士苹果枝条导水率、激素含量和花芽形成的影响 [J].中国农业科学,2013,46(17):3643-3650.
Li X W,Li B Z,Liu F T,et al.Effects of bud-notching, twistiing shoot and removing terminal shoot on branches hydraulic conductivity,hormone content and flower bud formation of Fuji apple [J].Scientia Agricultura Sinica,2013,46(17):3643-3650 .
[17] 刘 俊,吉晓佳,刘友良.检测植物组织中多胺含量的高效液相色谱法 [J].植物生理学通讯,2002,38(6):596-598.
Liu J,Ji X J,Liu Y L.High performance liquid chromatography method for measuring polyamine content in plant tissue [J].Plant Physiology Communications,2002,38(6):596-598.
[18] 陈海江,徐继忠,袁小乱,等.不同时期喷施外源多胺对新红星苹果坐果的影响 [J].河北农业大学学报,1999,22(2):47-49.
Chen H J,Xu J Z,Yuan X L,et al.Effects of polyamines application at different times on fruit set of ‘Starkrimson’ apple [J].Journal of Agricultural University of Hebei,1999,22(2):47-49.
[19] 徐继忠,陈海江,马宝焜,等.外源多胺对富士苹果花和幼果内源多胺与激素的影响 [J].园艺学报,2001,28(3):206-210.
Xu J Z,Chen H J,Ma B K,et al.Effect of exogenous polyamines on endogenous polyamines and hormones in flowers and young fruits of Fuji apple [J].Acta Horticulturae Sinica,2001,28(3):206-210.
[20] 徐继忠,陈海江,李晓东,等.外源多胺对核桃雌雄花芽分化及叶片内源多胺含量的影响 [J].园艺学报,2004,31(4):437-440.
Xu J Z,Chen H J,Li X D,et al.Effect of exogenous polyamines on female and male flower differentiation and content of endogenous polyamines in leaves of walnut [J].Acta Horticulturae Sinica,2004,31(4):437-440.
[21] 曾 骧,吴显荣,孟昭清,等.金冠苹果短枝芽和叶片在花芽分化期氨基酸变化的初步研究:Ⅰ.花芽大量开始分化期酸、碱氨基酸的含量和动态变化 [J].北京农业大学学报,1985,11(1):59-70.
Zeng X,Wu X R,Meng Z Q,et al.Preliminary studies on amino acids during the flower bud differentiation period in buds and leaves of Golden Delicious apple spurs:Ⅰ.the levels and dynamic patterns of basic and acidic amino acids during the period of mass flower bud initiation [J].Journal of Beijing Agricultural University,1985,11(1):59-70.
[22] 郑家基,谢厚钗.龙眼越冬期叶片多胺含量的变化 [J].园艺学报,1996,23(3):295-296.
Zheng J J,Xie H C.The content changes of polyamines in longan during wintering stage [J].Acta Horticulturae Sinica,1996,23(3):295-296.
[23] 陈 鸿,李智辉,李天来,等.新铁炮百合花芽分化及发育过程中内源多胺及激素含量变化的研究 [J].沈阳农业大学学报,2010,41(3):284-288.
Chen H,Li Z H,Li T L,et al.Changes in the contents of endogenous polyamines,hormones during the initiation and development of flower bud in lilium formologi [J].Journal of Shenyang Agricultural University,2010,41(3):284-288.
[24] 钟晓红,罗先实,陈爱华.李花芽分化与体内主要代谢产物含量的关系 [J].湖南农业大学学报,1999,25(1):31-35.
Zhong X H,Luo X S,Chen A H.A study on nai plum’s flower bud differentiation and its major content of metabolic production [J].Journal of Hunan Agricultural University,1999,25(1):31-35.
[25] Guo J E,Li T,Sun X Z.Relationship between endogenous polyamines and floral bud differentiation inChrysanthemummorifoliumunder short-day conditions [J].Kor J Hort Sci Technol,2015,33(1):31-38.
[26] Jose A,Gil-Amado·Maria C,Gomez-Jimenez.Regulation of polyamine metabolism and biosynthetic gene expression during olive mature-fruit abscission [J].Planta,2012,235:1221-1237.
[27] 王世平,宋长冰,雷建菊,等.环切对苹果短枝顶芽内源多胺水平的影响及其与成花的关系 [J].西北植物学报,1997,17(3):298-304.
Wang S P,Song C B,Lei J J,et al.Effect of girdling on the levels of endogenous polyamine in relation to floral bud [J].Acta Bot Boreal Occident Sin,1997,17(3):298-304.
[28] 徐东花,郭俊娥,徐 璐,等.多胺氧化酶活性及木质素沉积与菊花花芽分化的关系 [J].华北农学报,2014,29(3):164-169.
Xu D H,Guo J E,Xu L,et al.The relationship between polyamine oxidase activity and lignin deposition and chrysanthemum flower bud differentiation [J].Acta Agriculturae Boreali-Sinica,2014,29(3):164-169.
