黄萎病不同发生程度棉田中土壤微生物多样性
2018-04-19刘海洋张仁福
刘海洋,姚 举,张仁福,王 伟,余 璇,王 琦
1 中国农业大学植物保护学院, 北京 100193 2 新疆农业科学院植物保护研究所, 乌鲁木齐 830091 3 新疆农业大学农学院, 乌鲁木齐 830091
新疆是我国最大的棉花种植区,棉花年产量占全国六成以上,棉花区位优势明显。近年来随着棉区重心向新疆转移,棉种跨区调运、病区棉秸秆还田加之长年连作等问题突出,致使棉花黄萎病在新疆发生加重、复杂。调查表明,新疆棉花黄萎病发病田占比超过50.0%,病情指数达5.0以上的严重发病田占比25.0%以上[1],部分棉区该病危害十分较重,已成为制约新疆棉花生产的关键因素。
棉花黄萎病是系统性侵染的土传病害,其病原菌为大丽轮枝菌(Verticilliumdahliae),防治十分困难。生物防治被认为是治理棉花黄萎病最具潜力的方法[2],但是,实践应用中发现,在利用生防菌剂或生物有机肥等防治棉花黄萎病时,存在防治效果不稳定的问题。棉花根际土壤微生物、大丽轮枝菌、土壤之间通过复杂的相互作用形成的微生态系统,是影响棉花黄萎病发生、制约生防菌剂防效的重要因素。
作物根部土壤中微生物群落结构及其组成变化能反映土壤生态现状及变化趋势,对作物健康十分重要。研究认为,不同棉花品种[3-9]、长年连作[10-13]、秸秆还田[14]、作物套种[15]、水稻轮作[16]等都会造成棉田土壤中微生物多样性水平、土壤酶活性、土壤微生物量等发生改变。但是,目前缺乏对不同黄萎病发生程度棉田中的土壤微生物多样性差异的研究。本文以棉花黄萎病不同发生程度棉田土壤为研究对象,深入分析不同发病程度棉田土壤中微生物群落的多样性以及土壤理化性质差异,探究土壤中制约棉花黄萎病发生的关键因素,揭示棉花黄萎病发生与土壤微生物种群之间的相关性,为利用微生态调控手段进行棉花黄萎病的综合防治提供理论依据。
1 材料与方法
1.1 供试材料
1.1.1 供试菌株
大丽轮枝菌强致病力菌株V991及弱致病力菌株V250,由中国农业科学院植物保护研究所惠赠;本实验室保存的细菌菌株AL7。
1.1.2 土壤样品采集与处理
在阿克苏市、库尔勒市、石河子市三地分别选择棉花黄萎病重病田和无病田(轻病田)各1块。于4月20日、5月20日、6月20日、8月10日分别取棉花根围10cm深耕层土样,每块棉田对角线3点取样,每点间隔50m,每点取5份小样,每份小样500g土壤,混合成1份土样。共采集72份土样。
土样经2mm孔径过筛后分3份保存:一份保存于-80℃,供提取土壤DNA进行高通量测序分析;一份保存于4℃冰箱供土壤微生物数量分析;一份土样风干后保存,进行土壤理化性质分析。
1.1.3 取样棉田背景
阿克苏无病田为水稻轮作改良棉田(前茬为重病田);重病田为相邻长年连作棉田。
库尔勒取样重病田为无病田内隔离出,面积约3500m2,通过连续3a人工接种大丽轮枝菌使之成为重病田;轻病田为紧邻的自然棉田,管理措施一致。
石河子轻病田和重病田均为自然棉田,未进行人工干预。
1.1.4 培养基
马丁氏培养基(链霉素200mg/L),高氏一号培养基(重铬酸钾50mg/L),改良微菌核选择分离培养基(青霉素200mg/L)[17],肉汤培养基(Lysogeny broth medium,LB),胰蛋白胨大豆琼脂培养基(Tryptone soy agar medium,TSA),马铃薯葡萄糖琼脂培养基(Potato dextrose agar medium,PDA)。
1.2 大丽轮枝菌发酵液对细菌生长量影响测定
利用PDA培养液培养菌株V991和菌株V250,27℃下在120r/min摇床上振荡培养14d,之后8000r/min离心15min,收集上清液,利用0.22μm细菌过滤器过滤除菌,-4℃保存备用。空白PDA培养液作为对照。
试验设5个处理,分别接种培养24h的AL7菌液5μL:①20mL LB培养液;②16mL LB培养液+4mL大丽轮枝菌发酵上清液;③10mL LB培养液+10mL大丽轮枝菌发酵上清液;④4mL LB培养液+16mL大丽轮枝菌发酵上清液;⑤20mL大丽轮枝菌发酵上清液。每处理以相同比例的LB培养液+PDA空白培养液作为对照。30℃,200r/min培养10h,每隔2h利用分光光度计测量每处理OD600值,3次重复。培养10h后,取5次测量值的平均值作为衡量每处理中细菌AL7生长量的值。
1.3 土壤理化性质分析
分析4月20日与8月10日采集的2批土样的理化性质。每块棉田3点土样混合成1份土样。将处理好的土壤样品送交新疆土肥水农业科技工程中心进行统一化验。分析指标包括pH值、总盐、有机质、全氮、全磷、全钾、微生物氮。
1.4 土壤中可培养微生物数量检测
对阿克苏市、石河子市、库尔勒市采样田4个时期采集的72份土样,利用马丁式培养基、高氏一号培养基、TSA培养基通过平板稀释分离法检测土壤中可培养真菌、放线菌、细菌的数量。
1.5 土壤中大丽轮枝菌微菌核分离
取1.1.2中8月份采集的棉田土样,水筛法处理后在改良微菌核选择分离培养基上利用平板稀释法分离土壤中大丽轮枝菌微菌核;取石河子重病田4个时期采集的土样,按照上述方法进行大丽轮枝菌微菌核分离。
1.6 高通量测序
1.6.1 土壤样品总DNA提取及检测
选择4月和8月份采集的36份土壤样品,每份土样称取0.5g,利用BioTeke DP4001试剂盒提取土壤总DNA。送交北京百迈客生物科技有限公司构建DNA文库,采用Illumina Hiseq 2500 PE250模式进行测序。扩增用引物及预扩增程序:细菌 16S rRNA (V3+V4)区域引物:5′-CTCCTACGGGAGGCAGCA- 3′,5′-GGACTACHVGGGTWTCTAAT- 3′;样品检测PCR 预实验程序:95℃预变性5 min;95℃变性30s,50℃退火30s、72℃延伸40s,25cycles;72℃延伸7min。
1.6.2 测序数据质量评估
对原始数据进行拼接(FLASH[18],version),将拼接得到的序列进行质量过滤(Trimmomatic[19],version),并去除嵌合体(UCHIME[20],version),得到高质量的Tags序列。
1.6.3 OTU 分析
在相似性 97%的水平上对序列进行聚类(UCLUST[21],version 1.2.22),以测序所有序列数的0.005%作为阈值过滤OTU[22]。
1.6.4 物种注释及分类学分析
细菌16S数据库:Silva[23];物种注释RDP Classifier[24]:置信度阈值为0.8;多重比对:PyNAST[25];建树方法:邻接法(Neighbor-Joining);分类学树状图分析:MEGAN5[26];alpha多样性指数分析:Mothur[27]version v.1.30;Beta 多样性分析(n≥3):基于binary jaccard、bray curtis、(un)weighted unifrac(限细菌)多种算法呈现物种多样性矩阵。
1.7 数据分析
常规数据利用Excel、SPSS软件进行数据汇总、计算和作图。
2 结果与分析
2.1 大丽轮枝菌发酵液对细菌AL7生长量的影响

图1 细菌AL7在大丽轮枝菌发酵液中的生长趋势 Fig.1 The growth tendency of bacteria strain AL7 in supernatant of V. dahliaeA:肉汤培养液,Lysogeny broth;B:8比2的肉汤培养液与大丽轮枝菌发酵液混合液,8 compares 2 mixture of lysogeny broth with potato dextrose agar fluid medium;C:1比1的肉汤培养液与大丽轮枝菌发酵液混合液,1 compares 1 mixture of lysogeny broth with potato dextrose agar fluid medium;D:2比8的肉汤培养液与大丽轮枝菌发酵液混合液,2 compares 8 mixture of lysogeny broth with potato dextrose agar fluid medium;E:大丽轮枝菌发酵液,Potato dextrose agar fluid medium
随着大丽轮枝菌发酵上清液比例增加,培养10h后,不同处理中细菌AL7的生长量呈逐渐降低的趋势,而对照处理中细菌AL7的生长没有受到PDA培养液的抑制(图1)。结果表明,大丽轮枝菌菌株V991、V250的发酵上清液均能对细菌AL7的生长起到明显的抑制作用。
强致病力菌株V991发酵上清液对细菌AL7生长的抑制作用超过弱致病力菌株V250,说明致病力强菌株产生的次生代谢产物或营养竞争能力相比弱致病力菌株对细菌AL7的生长影响更大。
2.2 不同发病程度棉田土壤的理化性质分析
经化验分析(表1),阿克苏市、石河子市、库尔勒市3个地区6块取样棉田土壤样品的pH值在7.97—8.46之间,均为偏碱性土壤。库尔勒采样棉田盐份含量显著高于阿克苏市和石河子市采样棉田,是阿克苏市和石河子市采用棉田的6至10倍左右,阿克苏市经水稻轮作改良的无病棉田盐份含量显著低于相邻的常年连作重病田,体现了水旱轮作防病、降盐的特点。
从土壤肥力水平来看,石河子市采样棉田有机质、全氮、全磷含量均为3个采样地区中的最高值,其中石河子市采样重病田中有机质、全氮含量分别达到了38.66g/kg土和2.025g/kg土,约为石河子对照轻病田的2倍,土壤肥力水平高;库尔勒采样重病田土壤中有机质、全氮、全磷、全钾含量均与轻病田无明显差异;阿克苏市水稻改良后的无病棉田土壤中有机质、全氮、全磷、生物氮量均略高于相邻重病田。

表1 不同地区棉田土壤理化性质/(g/kg)
2.3 不同发病程度棉田土壤中可培养微生物数量分析
2.3.1 不同发病程度棉田土壤中的细菌数量
分析结果表明(表2),阿克苏市采样重病田中细菌数量除5月份低于对照田外,4月、6月、8月份3个时期土壤中细菌数量均显著高于轻病田;库尔勒市重病田土壤中4月份与8月份细菌数量高于对照田,5月份和6月份则低于对照田,重病田和对照轻病田中细菌数量趋势呈交替变化,均无显著差异;石河子市重病田4个时期土壤中细菌数量均高于轻病田,除8月份外,重病田土壤中的细菌数量均显著高于轻病对照田。
2.3.2 不同发病程度棉田土壤中的真菌数量
阿克苏市采样重病田中真菌数量除4月份低于无病对照田以外,5月份、6月份与8月份重病田土壤中真菌数量均高于无病对照田,其中6月份达到显著性差异;库尔勒市采样重病田土壤中真菌数量除6月份以外,均略低于轻病对照田,但4个时期均未达显著性差异;石河子市重病田其全生育期土壤中真菌数量均高于轻病田,且在4月份、5月份、6月份均达到显著差异(表3)。

表2 不同时期土壤中细菌数量/(×5×106)
a,b 代表显著性差异分析,相同字母代表无显著性差异,不同字母代表有显著性差异
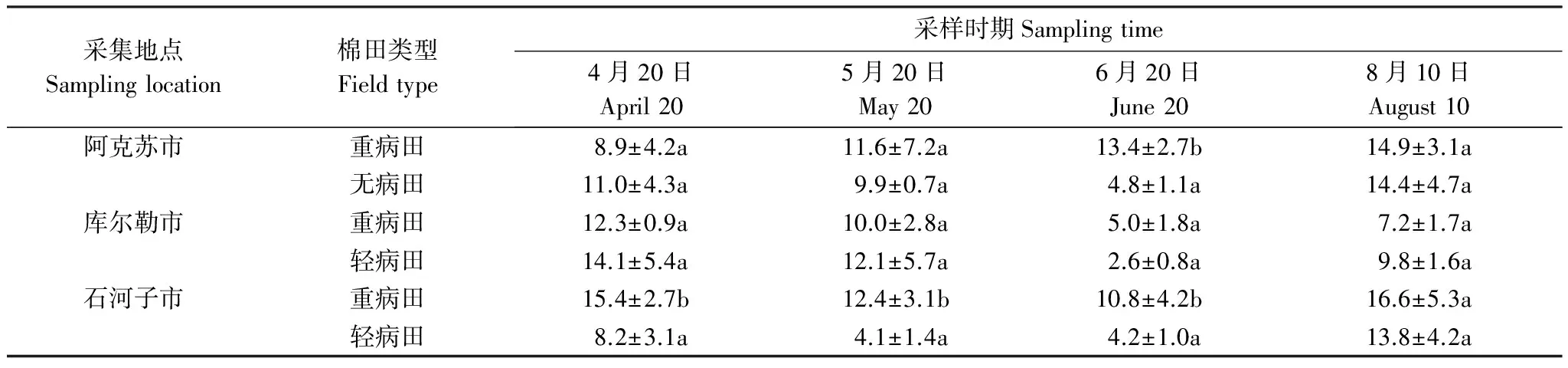
表3 不同时期土壤中真菌数量/(×5×105)
a,b 代表显著性差异分析,相同字母代表无显著性差异,不同字母代表有显著性差异
2.3.3 不同发病程度棉田土壤中放线菌数量
石河子市采样棉田中放线菌数量基数很大,每克土中最高可达1×108个。阿克苏市采样无病田土壤肥力指标均高于重病田(表1),无病对照田4个时期土壤中放线菌含量均高于重病田,且在5月份和8月份达到显著差异;库尔勒市重病田由人工接菌造成,轻病对照田为紧邻棉田,两地土壤肥力差异很小(表1),放线菌数量表现出呈交替变化的趋势,仅在8月份出现了显著差异;石河子市重病田土壤中有机质、全氮含量均显著高于轻病对照田,重病田4个时期土壤中放线菌的含量同样显著高于轻病田(表4)。

表4 不同时期土壤中放线菌数量/(×5×105)
a,b 代表显著性差异分析,相同字母代表无显著性差异,不同字母代表有显著性差异
2.3.4 不同发病程度棉田土壤中大丽轮枝菌微菌核数量
经分析(表5),阿克苏市、石河子市、库尔勒市每个地区重病田和轻病田土壤中大丽轮枝菌微菌核数量均达到显著性差异。
阿克苏市采样重病田8月10日棉花黄萎病病情指数为58.7,每克土中分离出的大丽轮枝菌微菌核数量为40个,而前期为重病田,经水稻轮作改良后的对照棉田黄萎病病情指数为0,其耕层土中未分离到大丽轮枝菌微菌核;石河子市采样重病田同时期棉花黄萎病病情指数为94.5,每克土中平均分离出的大丽轮枝菌微菌核数量为193个,对照轻病田每克耕层土中大丽轮枝菌微菌核数量为6个,病情指数为24.5;库尔勒市人工接菌重病田同期棉花黄萎病病情指数为80.8,每克土中大丽轮枝菌微菌核数量为73个,对照轻病田每克土中棉花黄萎病微菌核数量为17个,病情指数为2.5。
为探明新疆棉田土壤中大丽轮枝菌微菌核含量的消长动态,对石河子重病田4个时期采集的土壤中微菌核的含量进行了分析。结果显示,4月份石河子重病田每克土中大丽轮枝菌微菌核数量为119个;5月份采集土壤中大丽轮枝菌微菌核数量最高,每克土中微菌核数量达196个,至6月份略有降低,为155个/g土,8月份土壤中微菌核的数量为132个/g土,低于6月份,略高于4月份(图2)。土壤中大丽轮枝菌微菌核的含量全年出现1个高峰,呈现先增高后逐渐降低的趋势。
表5棉田黄萎病发生程度及10cm耕层土壤中大丽轮枝菌微菌核数量
Table5Microsclerotiumquantityofcottonfieldin10centimetersdeepanddiseaseindexofVerticilliumwilt

取样地点Samplinglocation菌核数量Microsclerotiumquantity病情指数Diseaseindex阿克苏40±1.21b58.7b0a0a石河子193±7.99b94.5b6±1.10a24.5a库尔勒73±2.64b80.8b17±0.98a2.5a
a,b 代表显著性差异分析,相同字母代表无显著性差异,不同字母代表有显著性差异
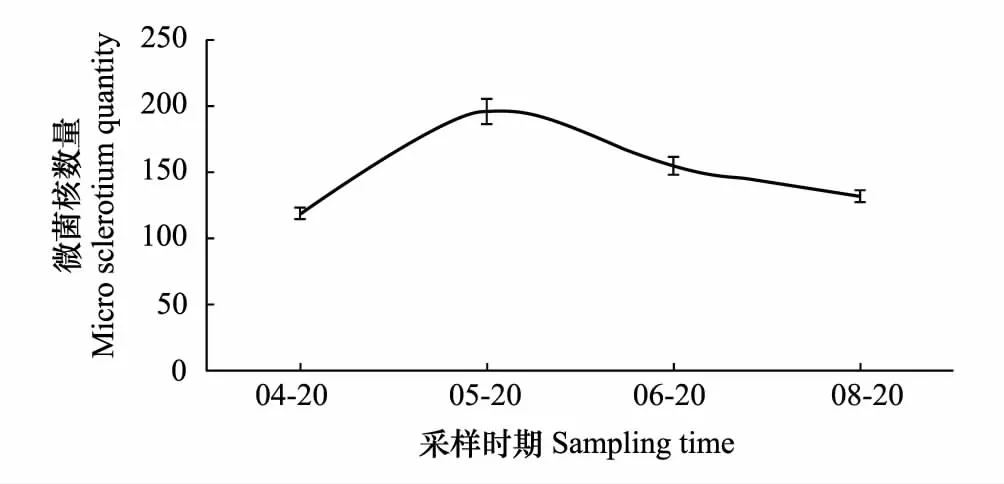
图2 不同时期棉田10cm耕层土壤中大丽轮枝菌微菌核含量的动态变化Fig.2 Dynamics of micro sclerotium quantity in 10 centimeters deep of cotton field in different times
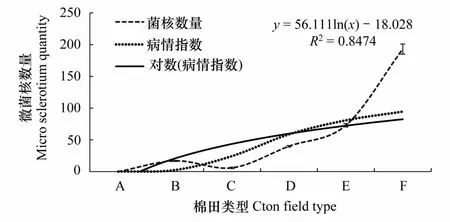
图3 大丽轮枝菌微菌核含量与棉花黄萎病发病程度的相关性 Fig.3 The correlationship of Micro sclerotium quantity and disease degree of Verticillium wilt A:阿克苏无病田, Disease-free field of Aksu;B:库尔勒轻病田, Mild disease field of Korla;C:石河子轻病田,Mild disease field of Shihezi;D:阿克苏重病田,Serious disease field of Aksu;E:库尔勒重病田,Serious disease field of Korla;F:石河子重病田,Serious disease field of Shihezi
2.4 土壤中可培养微生物数量的相关性与棉花黄萎病发生程度分析
2.4.1土壤中大丽轮枝菌微菌核含量与棉花黄萎病发生程度的相关性
阿克苏市无病棉田未检测到大丽轮枝菌微菌核,石河子市、库尔勒市发病严重棉田土壤中微菌核数量均显著高于发病轻棉田。相关性分析表明(图3),大丽轮枝菌微菌核含量与棉花黄萎病发生程度的R2= 0.8474(Sig.=0.036),为显著正相关,在一定范围内,大丽轮枝菌微菌核数量越多,棉花黄萎病发生越重。
2.4.2土壤中大丽轮枝菌微菌核含量与细菌、真菌、放线菌数量的相关性
试验分析证明棉花黄萎病的发生程度与土壤中的大丽轮枝菌微菌核含量呈显著正相关,而大丽轮枝菌发酵液能够对细菌生长产生抑制作用。但是,对不同棉田土壤中大丽轮枝菌微菌核含量与放线菌、真菌、细菌数量的相关性分析表明,土壤中各微生物种群之间没有显著相关性(表6)。

表6 微生物数量与棉花黄萎病发生程度相关性
2.5 棉田土壤细菌群落结构的组成与多样性分析
对36个土壤样品进行高通量测序,共获得4062258对Reads,双端Reads拼接、过滤后产生3052997条Clean tags,平均每个样品产生84805条Clean tags。
样本文库的覆盖率指数均在99.6%以上,反映本次测序结果代表了样本中细菌种群的真实情况。各样品的菌群丰度指数(Chao1、Ace)、生物多样性指数(Shannon、Simpson)表明采集的土壤样品中的细菌种群多样性较高(表7)。分析表明,在97%相似度水平下,阿克苏市、库尔勒市重病田和轻病田分别采集的样品之间的操作分类单元(OUT)数目存在显著性差异,而石河子市重病田和轻病田采集的样品生物OUT数目不存在显著性差异。

表7 不同样品中细菌群落多样性指数
OTUs:Operational taxonomic units,操作分类单元;a4w2、a4w4、a4w6: 4月份阿克苏无病田土壤样品;a8w1、a8w2、a8w3:8月份阿克苏无病田土壤样品;an4b1、an4b3、an4b6:4月份阿克苏重病田土壤样品;an8b1、an8b2、an8b3:8月份阿克苏重病田土壤样品;bn4w2、bn4w4、bn4w6:4月份石河子轻病田土壤样品;bn8w1、bn8w2、bn8w3:8月份石河子轻病田土壤样品;sn4b2、sn4b4、sn4b6:4月份石河子重病田土壤样品;sn8b1、sn8b2、sn8b3:8月份石河子重病田土壤样品;k4c1、k4c2、k4c3:4月份库尔勒轻病田土壤样品;k8c1、k8c2、k8c3:8月份库尔勒轻病田土壤样品;k4v1、k4v2、k4v3:4月份库尔勒重病田土壤样品;k8v1、k8v2、k8v3:8月份库尔勒重病田土壤样品
2.5.1 不同土壤中细菌群落结构的多样性分析

图4 不同样品中细菌群落多样性的NMDS分析Fig.4 NMDS analysis of bacterial community diversity NMDS:非度量多维尺度法,non-metric multidimensional scaling
根据36个土壤样品中细菌OTU类型的丰度值利用非度量多维尺度法(NMDS)进行多样性分析,比较不同样品在细菌OTUs组成上的差异。在应力值=0.055水平上,不同土壤中的细菌菌群受采集地点差异的影响最大(图4),同一棉田采集的土壤样品中细菌菌群相似,距离最近;继而是同一地区土壤样品中的菌群相似,距离次之。阿克苏市和石河子市土壤样品采集的棉田是相互独立的,因此在不同的棉田水平上,阿克苏、石河子采样重病田和轻病田土壤中的细菌菌群距离较近,均能够独立区分开来。而库尔勒市土壤样品中细菌菌群受采集棉田的影响较小,不同棉田土壤样品中细菌菌群距离较远;反而受采样时间的影响较大,同一时期采集的土壤中细菌菌群距离更近,表明棉田接种大丽轮枝菌并未对土壤中细菌菌群造成明显影响。
2.5.2 不同土壤中细菌群落结构的聚类分析

图5 不同样品中细菌群落组成的层次聚类分析 Fig.5 Hierarchical cluster analysis of bacterial community composition in different samples
对36个样品基于Beta多样性分析得到距离矩阵,通过非加权组平均法(UPGMA)进行层次聚类分析(图5)。同一地区采集的土壤样品均单独归为一支,其中阿克苏市采集的土壤样品与石河子市采集的土壤样品归为一个大的分支,库尔勒市采集的样品单独归为一支,在物种组成上阿克苏市土壤样品与石河子市土壤样品较库尔勒地区更近。
具体到采样棉田,阿克苏市同一棉田不同时期采集的土壤样品能够聚为一支;更进一步,同一时期的采集土壤样品归为一支。石河子市采集土壤样品的物种组成聚类情况与阿克苏相同,同一时期、同一棉田均能够单独归为一支。
库尔勒市采样棉田的物种组成聚类分析结果与阿克苏和石河子不同,由于库尔勒采样的重病田与轻病田为同一块棉田隔离而来,且管理措施一致,因此在棉田水平上没有把重病田与轻病田单独归为一支,反而是相同采样时期的土壤样品中物种组成更为接近,4月份采集的样品和8月份采集的样品各自归为一支,该结果与NMDS分析结果一致,充分说明人工接入大丽轮枝菌短期内对土壤细菌种群未产生明显影响。
3 讨论
本研究旨在分析棉田土壤中大丽轮枝菌与微生物群落变化之间的相关性,通过研究棉花黄萎病不同发病程度棉田土壤中微生物群落的结构差异,探究土壤中制约棉花黄萎病发生的关键因素,为利用微生态调控措施防控棉花黄萎病提供理论依据。
本文研究表明,在纯培养条件下,大丽轮枝菌无菌发酵滤液对细菌AL7的生长起明显的抑制作用,且强致病力菌株的抑制能力更高,可能是大丽轮枝菌的致萎毒素[28]抑制了细菌的生长或大丽轮枝菌生长消耗了PDA培养液中的营养物质所致。据此推测,当土壤中大丽轮枝菌种群达到一定数量时,可能会通过致萎毒素直接抑制或通过侵染棉花引起根系分泌物的改变来影响土壤微生物的种群结构[29-31]。但是,库尔勒市采样的无病棉田通过连续3年人工大量接种大丽轮枝菌使之成为棉花黄萎病重病田后,经土壤微生物数量分析发现,与轻病对照田相比,细菌、真菌、放线菌的数量动态呈交替变化的趋势,各微生物类群数量均没有明显的差异;结合高通量测序,对各样品进行NMDS和层次聚类分析,也得出接种大丽轮枝菌并未对土壤中细菌菌群遗传关系产生较大影响的结论。加之,石河子、阿克苏棉花黄萎病重病田中细菌数量反而显著高于轻病田(阿克苏重病田5月份样品除外),据此认为,短时期内大丽轮枝菌并不能够引起棉田土壤中细菌菌群的明显改变。
研究认为细菌型土壤是土壤肥力提高的一个生物指标[32],李秀英[33]、章家恩[34]等认为土壤养分含量与微生物数量存在较好的正相关性,土壤微生物的数量高低在一定程度上反映了土壤的肥力水平。石河子采样重病田为连作20年以上棉田,该棉田土壤肥力水平较高,土壤中的有机质、全氮含量2—4倍高于其他棉田;同时,该棉田黄萎病发病最重,病情指数接近100,土壤中大丽轮枝菌微菌核含量高达近200个/g土。高通量测序表明该田与石河子轻病对照田内细菌群落存在遗传差异,且重病田其土壤中细菌、真菌、放线菌数量均显著高于对照轻病田,与张海燕等对新疆南疆不同连作年限棉田土壤微生物群落结构研究的结果接近[35]。该结果证明了土壤肥力水平和微生物数量高的棉田并不能有效降低发病,而土壤中的大丽轮枝菌数量才是决定棉花黄萎病发生的主导因素。棉田土壤中有机质主要来源于棉籽饼以及棉秸秆还田等,当常年棉花连作加之带菌秸秆还田会造成大量的病原菌积累,就会导致棉花黄萎病的严重发生。控制该病关键在于利用生物防治、轮作、深翻等生态调控手段增加土壤中有益菌群数量,降低耕层土壤中菌源数量,同时种植抗病品种结合改善土壤生态、水肥条件加之健康的农事管理促进棉花健康生长,降低该病的危害。
土壤中是否存在大丽轮枝菌微菌核是棉花黄萎病能否发生流行的先决条件。研究认为,每克土壤中0.03个微菌核就可造成发病,每克土壤中微菌核3.5个以上该病的发病率可达100%[36];另有研究认为导致棉花黄萎病发病的临界量为0.5个/g土[37-38]。本文研究发现,石河子、库尔勒轻病田8月份每克土壤中微菌核数量分别为6个和17个,对应病情指数则分别为24.5和2.5,发病率未达100%,说明发病程度不是单纯的由菌源数量决定,而是由棉花品种的抗病性以及大丽轮枝菌的致病力等因素在黄萎病发生程度与菌核数量的相互关系上起到主导作用。阿克苏棉花黄萎病重病田经过水稻轮作后未发生黄萎病,土壤中也未分离到大丽轮枝菌微菌核,充分证明了菌源是该病发生的先决条件,同样也证明了水旱轮作对土传病害极佳的控制效果[15]。
土壤中大丽轮枝菌微菌核含量与细菌、真菌、放线菌数量的相关性分析同样证明,棉花黄萎病的发生与大丽轮枝菌微菌核含量呈显著正相关,与放线菌、真菌、细菌三大微生物种群之间没有显著相关性。制约棉田土壤中微生物群落的因素很多。前人研究认为,棉花抗枯萎病品种连作田枯萎病菌数量明显少于重病田[4-5],连作是新疆棉花种植中的突出问题,长期连作造成棉花根际微生物多样性水平降低,土壤中三大类微生物总数下降[10],病原拮抗菌减少,土壤酶活性下降[11]。秸秆还田虽然能提高连作棉田土壤微生物量的含量,缓解棉花连作的不利影响,提高土壤质量[14],但是从病原菌传播、积累的角度来看,当棉田黄萎病发生时该模式不利于控制棉花黄萎病的蔓延。而轮作可提高土壤酶活性和微生物群落结构多样性,水稻棉花轮作[16]、蒜棉套作、麦棉套作[15]均提高棉田土壤微生物中细菌和放线菌的数量,有效抑制棉田连作障碍的发生。
本文研究表明,水稻轮作能够有效降盐、减病、改善土壤肥力;接种大丽轮枝菌短期内不能造成土壤微生物群落结构的明显改变;常年连作的重病田在土壤肥力高的情况下,土壤中微生物种群数量会高于健康棉田。不同类型棉田土壤中微生物群落主要受土壤肥力、栽培措施、种植品种等因素影响,土壤中大丽轮枝菌微菌核数量与棉花黄萎病的发生程度之间呈显著正相关,但并未显著造成微生物数量的变化。改善棉田土壤微生物群落结构可能会一定程度促进棉田土壤生态健康,但是,通过生物防治水旱、轮作、机械深翻等调控措施降低棉田耕层土壤中病原菌数量,才是减轻棉花黄萎病危害的基础。
致谢: 感谢中国农业科学院植物保护研究所简桂良研究员惠赠2株大丽轮枝菌标准菌株,感谢新疆农垦科学院植物保护研究所刘政和新疆兵团第一师农科所武刚在土壤样品采集过程中给予的帮助。
参考文献(References):
[1] 刘海洋, 王伟, 张仁福, 武刚, 姚举. 新疆主要棉区棉花黄萎病发生概况. 植物保护, 2015, 41(3): 138- 142.
[2] 马平. 棉花黄萎病生物防治研究进展. 河北农业科学, 2003, 7(3): 38- 44.
[3] Neal J L Jr, Atkinson T G, Larson R I. Changes in the rhizosphere microflora of spring wheat induced by disomic substitution of a chromosome. Canadian Journal of Microbiology, 1970, 16(3): 153- 158.
[4] 吴传德, 延欣芳. 棉花抗枯萎病品种连作防病效果的研究. 植物保护学报, 1985, 12(3): 195- 200.
[5] 王守正, 王海燕, 李洪连, 于思勤, 袁红霞, 岳红宾. 植物微生物区系和植物抗病性研究. 河南农业科学, 2001, (5): 20- 23.
[6] 张美俊, 杨武德, 李燕娥. 不同生育期转Bt基因棉种植对根际土壤微生物的影响. 植物生态学报, 2008, 32(1): 197- 203.
[7] 李永山, 范巧兰, 陈耕, 柴永峰, 张冬梅, 李燕娥. 利用PLFA方法研究转Bt基因棉花对土壤微生物群落结构变化的影响. 棉花学报, 2009, 21(6): 503- 507.
[8] 范巧兰, 陈耕, 李永山, 柴永峰, 张冬梅, 李燕娥. 转Bt基因棉花不同生育期对土壤微生物的影响. 山西农业科学, 2010, 38(12): 34- 36.
[9] 娜布其, 红雨, 杨殿林, 赵建宁, 李刚, 娜日苏, 刘玲. 利用根箱法解析转双价(Bt+CpTI)基因棉花对土壤微生物数量及细菌多样性的影响. 棉花学报, 2011, 23(2): 160- 166.
[10] 施宠, 梁智, 徐万里, 单鸿宾, 贾宏涛, 王丽. 不同连作年限棉田土壤微生物和酶的特征. 新疆农业科学, 2010, 47(1): 163- 167.
[11] 李锐, 刘瑜, 褚贵新. 不同种植方式对绿洲农田土壤酶活性与微生物多样性的影响. 应用生态学报, 2015, 26(2): 490- 496.
[12] 高旭梅, 刘娟, 张前兵, 罗宏海, 谷天佐, 张旺锋. 耕作措施对新疆绿洲长期连作棉田土壤微生物、酶活性的影响. 石河子大学学报: 自然科学版, 2011, 29(2): 145- 152.
[13] 顾美英, 徐万里, 茆军, 梁智, 张志东, 房世杰. 连作对新疆绿洲棉田土壤微生物数量及酶活性的影响. 干旱地区农业研究, 2009, 27(1): 1- 5, 11- 11.
[14] 郭成藏, 李鲁华, 黄金花, 刘军, 杨志兰, 魏飞, 刘建国. 秸秆还田对长期连作棉田土壤微生物量碳氮磷的影响. 农业资源与环境学报, 2015, 32(3): 296- 304.
[15] 赵庆龙, 宋宪亮, 孙学振, 张美玲, 李宗泰. 蒜棉、麦棉套作对土壤微生物数量及相关酶活性的影响. 植物营养与肥料学报, 2011, 17(6): 1474- 1480.
[16] 徐幼平, 蔡新忠, 祝小祥. 水旱作物轮作田块土壤中微生物群落结构的PLFA法比较分析. 浙江农业学报, 2013, 25(5): 1056- 1061.
[17] 冯争光, 鹿秀云, 彭巧慧, 马平, 董金皋. 一种新的检测土壤棉花黄萎菌微菌核的选择性培养基. 华北农学报, 2004, 19(S): 44- 47.

[19] Bolger A M, Lohse M, Usadel B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics, 2014, 30(15): 2114- 2120.
[20] Edgar R C, Haas B J, Clemente J C, Quince C, Knight R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics, 2011, 27(16): 2194- 2200.
[21] Edgar R C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics, 2010, 26(19): 2460- 2461.
[22] Bokulich N A, Subramanian S, Faith J J, Gevers D, Gordon J I, Knight R, Mills D A, Caporaso J G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nature Methods, 2013, 10(1): 57- 59.
[23] Quast C, Pruesse E, Yilmaz P, Gerken J, Schweer T, Yarza P, Peplies J, Glöckner F O. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Research, 2013, 41(D1): D590-D596.
[24] Wang Q, Garrity G M, Tiedje J M, Cole J R. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Applied and Environmental Microbiology, 2007, 73(16): 5261- 5267.
[25] Caporaso J G, Bittinger K, Bushman F D, DeSantis T Z, Andersen G L, Knight R. PyNAST: a flexible tool for aligning sequences to a template alignment. Bioinformatics, 2010, 26(2): 266- 267.
[26] Huson D H, Auch A F, Qi J, Schuster S C. MEGAN analysis of metagenomic data. Genome Research, 2007, 17(3): 377- 386.
[27] Schloss P D, Westcott S L, Ryabin T, Hall J R, Hartmann M, Hollister E B, Lesniewski R A, Oakley B B, Parks D H, Robinson C J, Sahl J W, Stres B, Thallinger G G, Van Horn D J, Weber C F. Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Applied and Environmental Microbiology, 2009, 75(23): 7537- 7541.
[28] 桑茜, 袁虹霞, 王振跃, 李洪连, 汪敏. 河南省不同地区棉花黄萎病菌分离物致病性及其毒素致萎活性测定. 棉花学报, 2010, 22(4): 333- 338.
[29] 董丽红, 郭庆港, 张晓云, 李社增, 鹿秀云, 马平. 棉花根系分泌物对枯草芽胞杆菌NCD- 2生物膜形成和根际定殖的影响. 植物病理学报, 2015, 45(5): 541- 547.
[30] 孙磊, 陈兵林, 周治国. 麦棉套作Bt棉花根系分泌物对土壤速效养分及微生物的影响. 棉花学报, 2007, 19(1): 18- 22.
[31] 赵小亮, 刘新虎, 贺江舟, 万传星, 龚明福, 张利莉. 棉花根系分泌物对土壤速效养分和酶活性及微生物数量的影响. 西北植物学报, 2009, 29(7): 1426- 1431.
[32] 张丽娟, 曲继松, 郭文忠, 杨冬艳, 冯海萍. 微生物菌肥对黄河上游地区设施土壤微生物及酶活性的影响. 中国土壤与肥料, 2014, (5): 32- 36.
[33] 李秀英, 赵秉强, 李絮花, 李燕婷, 孙瑞莲, 朱鲁生, 徐晶, 王丽霞, 李小平, 张夫道. 不同施肥制度对土壤微生物的影响及其与土壤肥力的关系. 中国农业科学, 2005, 38(8): 1591- 1599.
[34] 章家恩, 刘文高, 胡刚. 不同土地利用方式下土壤微生物数量与土壤肥力的关系. 土壤与环境, 2002, 11(2): 140- 143.
[35] 张海燕, 贺江舟, 徐彪, 龚明福, 张利莉. 新疆南疆不同连作年限棉田土壤微生物群落结构的变化. 微生物学通报, 2010, 37(5): 689- 695.
[36] 董金皋. 农业植物病理学. 北京: 中国农业出版社, 2001: 203- 207.
[37] 石磊岩, 简桂良, 孙文姬, 冯洁. 我国棉花黄萎病研究现状与方向. 植物保护学报, 1998, 25(2): 103- 107.
[38] 简桂良, 邹亚飞, 马存. 棉花黄萎病连年流行的原因及对策. 中国棉花, 2003, 30(3): 13- 14.
