锰胁迫对野大豆种子萌发及幼苗生理生化特征的影响
2018-03-31刘胜
, , , , 刘胜,2
(1.中南林业科技大学生命科学与技术学院, 长沙 410018;2.湖南省环境资源植物开发与利用研究中心, 长沙 410018)
植物修复具有高效、环保和廉价等优点,具有广阔应用前景的重金属污染治理方法[1]。筛选出合适的修复植物是推广应用该方法的前提。矿山自然生长的豆科植物是优良的修复植物资源,因为与其它植物相比,它们不但能够适应矿山重金属污染环境,而且能够有效固氮、提高土壤肥力,从而更好地治理矿山污染。
野大豆(Glycinesoja) 是栽培大豆 (G.max)的近缘种,具有抗旱、耐旱、耐碱盐等优点,是大豆品种改良的野生资源库;该植物还具有良好的固氮能力,并被列为我国国家Ⅱ级保护植物;因而该植物具有重要的生态、经济,甚至战略价值[2]。尤其值得一提的是,该植物能够在锰矿矿区正常生长、开花结实[3-4],是一种有开发价值的锰污染修复植物。目前,该植物的研究主要集中于品种改良[5-6]、形态解剖[7-8]、生物学与生态学特性[9-10]及群体的遗传分化[11-12]等方面,锰胁迫对种子萌发的影响方面的研究较少。
种子萌发及幼苗生长是植物对环境胁迫最敏感的时期,其特征反映了植物对胁迫的耐性[13]。种子的萌发及幼苗生长特征是植物对胁迫的直接反应,是适应胁迫的重要特征。萌发过程中幼苗渗透调节物质、抗氧化酶活性等的改变在一定程度上反映种子萌发过程中代谢、生长情况。这些特征是揭示植物适应环境的重要依据。
本实验研究锰胁迫对野大豆种子萌发特性以及幼苗生理生化特征的影响,揭示该植物适应锰胁迫的生理生化机制。研究结果可为栽培大豆抗性品种的选育以及野大豆应用于锰污染治理的实践提供第一手资料。
1 材料与方法
1.1 材料采集
种子采集时间为2016年11月下旬野大豆种子成熟季节,地点为湖南省湘潭锰矿区,该矿开采始于100多年前,是我国最早的锰矿基地,20世纪80年代之前锰矿开采的主要方式为露天开采。锰矿开采完毕后,部分地段将矿渣与客土混合,然后任其自然恢复。现场调查显示,该恢复地有大面积野大豆分布。选择矿区100个植株采集成熟种子,为使种子覆盖面更广,各植株间株距超过2 m。种子采集后放于纸质采集袋,带回实验室于室温下储藏备用。
1.2 萌发实验
挑选颗粒整齐饱满、大小均一、无伤害的种子开展发芽实验。种子在0.2%的KMnO4中消毒10 min后,用蒸馏水冲洗后晾干。萌发实验在铺有2层滤纸的培养皿中进行,在其中设置6个锰浓度模拟土壤锰胁迫梯度 (0.005,1,5,8,12 mmol/L和15 mmol/L)。各处理浓度设5个重复,共30个处理(培养皿)。每个培养皿内放置30粒种子,共计900粒种子。由于预实验结果显示野大豆在25 ℃下萌发较好,萌发实验选择在25 ℃的人工气候箱中进行,光周期为12 h白天/12 h黑夜。每天向培养皿中添加去离子水以维持培养液原有水平。种子萌发期间,每天观察并记录发芽情况。萌发实验开展至无新种子萌发,共持续12 d。第8天统计发芽势,12 d测定发芽率。萌发完毕后,测定根长、芽长、芽鲜重、根鲜重、根干重、芽干重等指标。
1.3 测定方法
1) 萌发指标。
发芽能力指标参照《国际种子检验规程》[14]来计算发芽率、发芽势、发芽指数和活力指数,具体计算方法如下:
发芽率(%)=第8天内正常发芽的种子数/供试种子总数×100%;
发芽势(%)=第4天内正常发芽的种子数/供试种子总数×100%;
发芽指数(GI)=∑(Gt/Dt) (式中,Dt为发芽日数,Gt为在t日内的发芽数);
活力指数(VI)=GI×S(式中,GI为发芽指数,S为幼苗高度)。
2) 生物量相关指标。
幼苗芽长、根长、鲜重、干重的测定:萌发第12天时,每处理随机选取10株幼苗,采用精度为0.1 mm的游标卡尺分别测量幼苗芽长、根长;并用滤纸吸去表面水分后称量鲜重,然后再于95 ℃下杀青10 min,65 ℃下烘至恒重。
根冠比=根 (地下部分)干重/芽 (地上部分)干重[15]。
3) 幼苗生理生化特性测定。
于处理第8天测定种子萌发过程中的可溶性糖、可溶性蛋白和脯氨酸含量。第10天测定幼苗的超氧化物歧化酶 (SOD)、过氧化物酶 (POD) 活性和丙二醛 (MDA) 含量。
可溶性糖含量的测定采用蒽酮比色法[16],可溶性蛋白质含量的测定采用考马斯亮蓝G-250 法[16],游离脯氨酸含量的测定采用茚三酮显色法[16],超氧化歧物酶活性的测定采用氮蓝四唑 (NBT) 光化还原法[16],过氧化物酶活性的测定采用愈创木酚法[16]。
表2锰胁迫对野大豆幼苗生长的影响
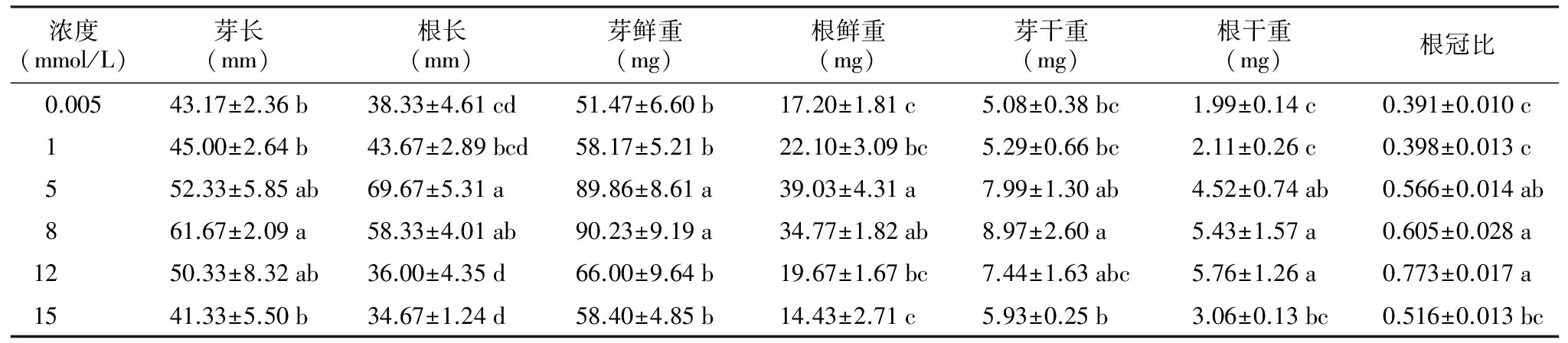
浓度(mmol/L)芽长(mm)根长(mm)芽鲜重(mg)根鲜重(mg)芽干重(mg)根干重(mg)根冠比0.00543.17±2.36b38.33±4.61cd51.47±6.60b17.20±1.81c5.08±0.38bc1.99±0.14c0.391±0.010c145.00±2.64b43.67±2.89bcd58.17±5.21b22.10±3.09bc5.29±0.66bc2.11±0.26c0.398±0.013c552.33±5.85ab69.67±5.31a89.86±8.61a39.03±4.31a7.99±1.30ab4.52±0.74ab0.566±0.014ab861.67±2.09a58.33±4.01ab90.23±9.19a34.77±1.82ab8.97±2.60a5.43±1.57a0.605±0.028a1250.33±8.32ab36.00±4.35d66.00±9.64b19.67±1.67bc7.44±1.63abc5.76±1.26a0.773±0.017a1541.33±5.50b34.67±1.24d58.40±4.85b14.43±2.71c5.93±0.25b3.06±0.13bc0.516±0.013bc
1.4 数据处理
种子萌发及幼苗生长各指标均以“平均值±标准差”表示,并采用SPSS 19.0软件进行单因素方差分析(One-way ANOVA),利用最小显著差数法(LSD法)对不同处理间的差异显著性进行多重比较,并用Sigma Plot 12.5软件作图。
2 结果与分析
2.1 种子萌发与幼苗生长
种子萌发与幼苗生长受锰胁迫的直接影响。表1显示,不同锰浓度下野大豆种子发芽率在35.0%~72.5%之间,随着锰浓度的增高,野大豆种子发芽率呈先增加后下降的趋势。与对照相比,在锰浓度为5,8,12,15 mmol/L时,发芽率分别下降15.79%、36.84%、47.37%、50.87%,说明一定浓度的锰抑制野大豆种子发育,发芽势、发芽指数表现出与发芽率一致的趋势(表1)。活力指数综合了种子发芽速率和生长量,能较好地反映种子萌发质量。表1显示,野大豆种子活力指数随着锰浓度的增加呈先上升后下降趋势,在锰浓度为8 mmol/L时达到最大值,然后下降;与对照相比,较低锰浓度 (1,5,8,12 mmol/L)下,其值均显著高于对照,分别比对照增加8.00%、52.81%、59.20%、7.72%;锰浓度为15 mmol/L时,其值则较对照降低36.87%。
表1锰胁迫对野大豆种子萌发的影响

浓度(mmol/L)发芽势(%)发芽率(%)发芽指数活力指数0.00531.25±2.88c71.25±2.92a8.24±0.07a565.60±7.18d136.25±3.53a72.50±1.89a7.61±0.21ab610.86±6.02c532.50±3.53b60.00±1.30b6.71±0.11ab864.29±5.16b821.25±2.89d45.00±4.18c7.20±0.18ab900.46±5.77a1221.25±2.50d37.50±1.89d7.11±0.41ab609.28±2.36c1518.75±2.73e35.00±1.32d4.90±0.46b357.05±3.81e
注:同列不同小写字母表示处理间差异显著(p<0.05),下同。
幼苗生长是植物对锰胁迫响应的综合体现。表2显示,随着锰浓度的增加,野大豆幼苗根长、芽长均呈先增加后降低的趋势,均在5 mmol/L与8 mmol/L时达到最大值,然后逐渐下降。与对照相比,较低锰浓度(1,5,8,12 mmol/L)时其值均较高,而较高浓度(15 mmol/L)时则较低。幼苗的鲜重、干重以及根冠比均表现出相同的趋势,说明锰胁迫下野大豆幼苗生长表现出低浓度促进生长、高浓度抑制生长的规律。野大豆在高浓度锰胁迫下根与芽生长的抑制效果较为明显,尤其是对根的胁迫。
2.2 可溶性糖、可溶性蛋白和游离脯氨酸含量的变化
可溶性糖、可溶性蛋白和游离脯氨酸不仅是组成细胞等结构的主要成分,也是增加渗透性物质的重要组成成分,是植物抗性的重要指标。图1显示,野大豆幼苗的可溶性糖含量随着锰浓度的升高呈上升趋势。其中,锰离子浓度为1 mmol/L时最低,其值为0.57 mg/g(FW),在锰离子浓度为12 mmol/L时最高,其值为0.95 mg/g(FW);与对照相比,除1 mmol/L时较低外,其余各浓度均有一定程度增加。
图1还显示,野大豆幼苗可溶性蛋白含量随着锰浓度的升高呈先上升后下降的趋势。当锰浓度为1 mmol/L时最高,锰浓度为15 mmol/L时最低;与对照相比,最高可溶性蛋白含量较对照增加10.36%,在锰浓度为15 mmol/L时比对照减少了13.59%。
野大豆幼苗脯氨酸含量也呈先增加后降低的趋势,但整体均高于对照。在锰浓度为5 mmol/L时达到最大值9.72μg/kg,比对照高出90.21% (图1),随后在锰浓度为8,12,15 mmol/L时有所下降,但仍分别比对照增加了38.58%、29.63%、12.83%。
2.3 抗氧化酶活性和膜脂过氧化
SOD、POD为植物体内重要的抗氧化酶。图2显示,随着锰浓度增加,野大豆SOD活性呈上升趋势;与对照相比,其含量分别增加4.27%、6.45%、52.99%、79.48%和146.44%(图2)。
图2显示,野大豆幼苗POD活性随锰浓度增加呈先增加后降低的趋势。在8 mmol/L时达到最大值,比对照增加8.13%,随后逐渐下降;与对照相比,在12 mmol/L时显著低于对照,当锰浓度达到15 mmol/L时,POD活性比对照减少了80.30%。
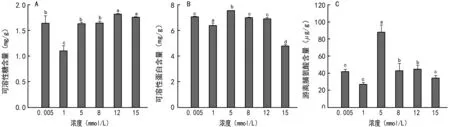
图1 锰胁迫对野大豆种子萌发过程中可溶性糖(A)、可溶性蛋白(B)和脯氨酸(C)含量的影响

图2 锰胁迫对野大豆幼苗超氧化物歧化酶(A)、过氧化物酶(B)的影响
丙二醛 (MDA) 是细胞膜脂过氧化反应的产物,其含量的变化可作为检测逆境条件下膜系统受损程度的指标。图3显示,随着锰浓度的增加,野大豆幼苗丙二醛含量呈先降低后增加的趋势;与对照相比,在12 mmol/L时显著高于对照,比对照增加44.44%(图3)。

图3 锰胁迫对野大豆幼苗丙二醛含量的影响
3 讨 论
3.1 种子萌发与幼苗生长
种子萌发与幼苗生长是植物生活史中对外界环境变化最敏感的阶段,能够较好地反映植物的适应能力[17];发芽率、发芽势和活力指数均为衡量种子活力的基础指标。本实验野大豆种子的发芽率、发芽势、发芽指数随着锰浓度的升高均呈下降趋势,这与张秀玲等[18]对盐胁迫下野大豆的研究、赵红玉等[19]对Cu、Cd、Pb、Zn等4种重金属胁迫下红豆草(Onobrychisviciaefolia)、白三叶(Trifoliumrepens)、柠条(Caraganakorshinskii)、胡枝子(Lespedezabicolor)4种豆科植物的研究结果一致。这说明重金属胁迫下植物发芽受到抑制是一种普遍现象。
本研究显示,锰胁迫对野大豆的生长表现出低浓度促进和高浓度抑制的效应。这与张芬琴等[20]、梁文斌等[21]、张秀玲等[18]的研究结果一致,说明适量的重金属可促进植物幼苗生长。本研究还显示,根受到的抑制程度明显高于芽,这与张芬琴等[20]、梁文斌等[21]的研究结果一致,而张秀玲等[18]则发现,盐胁迫下野大豆芽受到的抑制程度明显高于根,这说明不同重金属对幼苗生长的影响不一致。例如,过量的锰引起幼苗中分生细胞减少,而根尖分生区是直接接触锰且易受锰毒危害的部位[21],因此锰毒对幼苗胚根的抑制程度大于胚芽,而野大豆胚根和胚芽对盐胁迫的敏感程度有所不同,胚芽大于胚根,因而胚芽受到的影响较大。关于不同重金属对植物幼苗生长的相关机制尚需进一步研究。
3.2 可溶性糖、可溶性蛋白和游离脯氨酸含量
可溶性糖、可溶性蛋白和游离脯氨酸均为渗透调节物质。受到重金属胁迫时,植物通过生成渗透调节物质来维持细胞正常代谢环境,以减轻重金属毒害[22]。可溶性糖作为植物碳代谢的物质之一,与植物重金属抗性密切相关。本研究显示,野大豆幼苗可溶性糖含量呈现出随着锰浓度的升高而上升的趋势。这与张小艾等[23]、蒋和平等[24]的研究结果一致。其原因可能是低浓度锰尚不能刺激野大豆发生防御性反应,而是直接破坏了可溶性糖分子结构或抑制可溶性糖合成,导致其含量降低;随着锰浓度的升高,植物的自我保护机制促进糖的合成,以满足幼苗生长所需的营养物质。
可溶性蛋白含量是植物体内酶系统稳定的标志。本研究发现,野大豆幼苗可溶性蛋白含量呈先上升后下降趋势。这与张静等[25]的研究结果一致。其原因可能是可溶性蛋白含量的变化是植物响应重金属毒害的一种解毒机制,而锰的过量积累则导致细胞结构遭到破坏、蛋白质变性,使高浓度下可溶性蛋白含量下降[1]。
脯氨酸是一种渗透保护剂,具有维持细胞渗透压、清除活性氧、保护蛋白质及细胞膜的作用,植物体内积累的游离脯氨酸含量与重金属耐性紧密相关[26-27]。本研究显示锰胁迫下野大豆脯氨酸含量呈先上升后下降的趋势。这与张虎等[28]的研究结果一致。其原因可能是低浓度锰胁迫下,为抵抗外界伤害,植物体积累脯氨酸;而高浓度锰胁迫下植物受毒害程度增加,渗透调节平衡被打破,细胞无法修复,导致脯氨酸含量有所下降。
3.3 抗氧化酶活性和膜脂过氧化
SOD可催化超氧化物阴离子自由基(O2·-)发生歧化反应,有效阻止氧自由基在植物细胞内的积累,从而减轻毒害作用[29]。锰是超氧化物歧化酶的重要组成元素以及活化剂,可促使超氧化物歧化酶活性增强。POD则利用各种基质作为电子供体将H2O2还原为水,从而减轻超氧化物的毒害作用。本研究结果显示野大豆幼苗的超氧化物歧化酶活性随着锰浓度的升高而上升,过氧化物酶活性随着锰浓度的增加先升高后下降。这与於丙军等[30]以及张虎等[28]的研究结果一致。这表明低锰胁迫刺激植物体内保护酶活性提高,以抵御氧自由基对植物的伤害,提高幼苗在锰胁迫下的自我调节能力和耐性;随着锰胁迫程度的继续加强,细胞抗氧化能力逐渐减弱。
丙二醛是植物在环境胁迫下脂质过氧化而产生的一种具有细胞毒性的物质,是一种受伤害的指标[31],细胞膜脂的过氧化伤害程度可通过分析膜脂过氧化产物丙二醛的含量来评价。本研究中野大豆幼苗丙二醛含量随锰浓度的增加呈下降-上升的趋势。这与於丙军等[30]、王松华等[32]的研究结果一致。其原因可能是在低浓度锰胁迫下,由于保护酶系统的启动,保护酶对丙二醛起到了一定的抑制作用,其含量有所降低;随着重金属浓度继续增大,保护酶逐渐降低,丙二醛抑制作用减弱。
参考文献:
[1]李洋,于丽杰,金晓霞.植物重金属胁迫耐受机制[J].中国生物工程杂志,2015,35(9):94-104.
[2]林雅茹,唐宏亮,申建波.野生大豆根系形态对局部磷供应的响应及其对磷吸收的贡献[J].植物营养与肥料学报,2013,19(1):158-165.
[3]李有志,罗佳,张灿明,等.湘潭锰矿区植物资源调查及超富集植物筛选[J].生态学杂志,2012,31(1):16-22.
[4]李韵诗,石润,吴晓芙.湘潭锰矿区本土先锋植物耐受与积累锰的特性分析[J].广西林业科学,2015,44(1):25-30.
[5]苏伯鸿,李忠峰,邱丽娟.一个大豆理想株型突变体it1的表型和生理鉴定[J].植物遗传资源学报,2016,17(3):523-528.
[6]夏正俊.大豆基因组解析与重要农艺性状基因克隆研究进展[J].植物学报,2017,52(2):148-158.
[7]邹淑华,罗希明.栽培大豆(Glycinemax)与野生大豆(G.soia)解剖[J].大豆科学,1991(4):320-326.
[8]高伟,陆静梅,段肖,等.不同生态环境下野生大豆根解剖结构演化研究[J].河南农业科学,2016,45(1):46-49.
[9]徐豹,路琴华,庄炳昌.中国野生大豆(G.soja)生态类型的研究[J].中国农业科学,1987,20(5):29-35.
[10]吴东丽,张金屯,王春乙,等.野生大豆群落主要种群生态位特征研究[J].草地学报,2009,17(2):166-173.
[11]范虎,赵团结,丁艳来,等.中国野生大豆群体特征和地理分化的遗传分析[J].中国农业科学,2012,45(3):414-425.
[12]向仕华,王吴彬,何庆元,等.多环境下野生大豆染色体片段代换系群体农艺性状相关QTL/片段的鉴定[J].中国农业科学,2015,48(1):10-22.
[13]蔺吉祥,穆春生.松嫩草地羊草种子发育进程、休眠特性及与盐碱耐性关系的研究[J].草地学报,2016,24(2):479-482.
[14]国际种子检验协会(ISTA).国际种子检验规程[M].北京:北京农业大学出版社,1985:54-57.
[15]吴雅薇,李强,豆攀,等.低氮胁迫对不同耐低氮玉米品种苗期伤流液性状及根系活力的影响[J].植物营养与肥料学报,2017,23(2):278-288.
[16]张志良,瞿伟菁,李小方.植物生理学实验指导(第3版)[M].北京:高等教育出版社,2003:127-130.
[17]Nasr S M H,Savadkoohi S K,Ahmadi E.Effect of different seed treatments on dormancy breaking and germination in three species in arid and semi-arid lands[J].Forest Science and Practice,2013,15(2):130-136.
[18]张秀玲,李瑞利,石福臣.盐胁迫对野大豆种子萌发特性的影响[J].种子,2007,26(8):21-24.
[19]赵玉红,拉巴曲吉,罗布,等.铜、镉、铅、锌对4种豆科植物种子萌发的影响[J].种子,2017,36(1):22-28.
[20]张芬琴,金自学.两种豆科作物的种子萌发对Cd2+处理的不同响应[J].农业环境科学学报,2003,22(6):660-663.
[21]梁文斌,薛生国,沈吉红,等.锰胁迫对垂序商陆生长发育的影响[J].生态学杂志,2011,30(8):1 632-1 636.
[22]Batool R,Hameed M,Ashraf M,et al.Structural and functional response to metal toxicity in aquaticCyperusalopecuroides,Rottb.[J].Limnologica-Ecology and Management of Inland Waters,2014,48(7):46-56.
[23]张小艾,李名扬,汪志辉,等.重金属及盐碱对二月兰幼苗生长和生理生化的影响[J].草业学报,2013,22(2):187-194.
[24]蒋和平,郑青松,朱明,等.条浒苔和缘管浒苔对镉胁迫的生理响应比较[J].生态学报,2011,31(16):4 525-4 533.
[25]张静,赵秀侠,汪翔,等.重金属镉(Cd)胁迫对水芹生长及生理特性的影响[J].植物生理学报,2015(11):1 969-1 974.
[26]Fukuoka S,Saka N,Koga H,et al.Loss of function of a proline-containing protein confers durable disease resistance in rice[J].Science,2009,325(5 943):998-1 001.
[27]Singh V,Bhatt I,Aggarwal A,et al.Proline improves copper tolerance in chickpea (Cicerarietinum)[J].Protoplasma,2010,245(1):173.
[28]张虎,寇江涛,师尚礼.紫花苜蓿种子萌发对钴胁迫的生理生化响应[J].草业学报,2015,24(9):146-153.
[29]Bhaduri A M,Fulekar M H.Antioxidant enzyme responses of plants to heavy metal stress[J].Reviews in Environmental Science and Bio/Technology,2012,11(1):55-69.
[30]於丙军,刘友良.盐胁迫对一年生盐生野大豆幼苗活性氧代谢的影响[J].西北植物学报,2003,23(1):18-22.
[31]Fan L,Wang Q,Lv J,et al.Amelionration of post harvest chilling injury in cowpea (Vignasinensis) by methyl jasmonate (MeJA) treatments.Scientia Horticulturae,2016,203:95-101.
[32]王松华,张华,何庆元.铜胁迫对紫花苜蓿幼苗叶片抗氧化系统的影响[J].应用生态学报,2011,22(9):2 285-2 290.
(本栏目责任编辑:周介雄、戴燚)
