黄淮稻区优质抗病粳稻“六位一体”育种技术体系的构建与实践
2018-01-27李俊周王书玉尹海庆姬生栋王生轩赵全志
李俊周,王书玉,尹海庆,姬生栋,王生轩,张 栩,陈 楠,赵全志*
(1.河南农业大学 农学院,河南 郑州 450002; 2.新乡市农业科学院,河南 新乡 453002; 3.河南省农业科学院,河南 郑州 450002; 4.河南师范大学,河南 新乡 453007; 5.合肥丰乐种业股份有限公司,安徽 合肥 230031)
水稻是我国最重要的粮食作物,播种面积约占我国粮食作物总播种面积的29%,总产量约占40%。水稻还是我国的优势农作物,是单产水平最高的粮食作物,我国也是世界第一大水稻生产国,稻谷总产量居世界第一位。同时,我国又是世界第一大稻米消费国,约60%以上的人口以大米为主食,消费量占总粮食消费量的40%。从稻米消费总量来看,我国已由稻米净出口国转变为稻米净进口国;从稻米消费市场需求的质量结构上看,随着人民生活水平的不断提高,市场对大米品质的要求越来越高,劣质米相对过剩,中高档优质米缺乏,高档优质米市场需求量增长较快,高产高效无污染地生产出足够的优质稻米是国家粮食安全的保障。
1 稻米外观米质和稻瘟病是影响黄淮稻区粳稻生产的主要问题
稻米品质的主要衡量指标包括碾磨品质的整精米率、外观品质的长宽比、垩白度和垩白粒率和蒸煮食味品质的直链淀粉含量与胶稠度[1]。从稻米的外观品质和商品价值看,稻米粒型和垩白是最重要的决定因素[2]。对于黄淮稻区粳稻来说,垩白是制约稻米品质的首要因素。垩白是指稻米胚乳中白色不透明的部分,包括腹白、心白和背白,主要是由于籽粒灌浆过程淀粉粒排列疏松,存在空腔。垩白不仅会降低稻米碾磨品质和外观品质,也会造成稻米的口感变劣,少或无垩白的稻米商品价值高[3-4]。目前,已有大量的垩白相关QTL/基因被发现,但只有G1F1[5]、cyPPDK[6]、Chalk5[7]等少数几个被克隆。遗传学研究证明,垩白的形成是受许多QTL/基因控制的复杂调控系统的影响[7-8]。
稻瘟病是由子囊菌Magnaporthegrisea(He-bert) Barr引起的世界毁灭性水稻病害之一。据统计,稻瘟病每年给全球带来11%~30%的产量损失,相当于6.00 × 107人 1 a的口粮。中国的稻瘟病危害也相当严重,自上世纪90年代以来,年发生面积均在3.8×106hm2以上,年损失稻谷达数亿公斤[9]。根据发病时期可以分为:苗瘟、叶瘟、穗颈瘟。依据发病部位可以分为:节瘟、叶枕瘟、茎秆瘟和谷粒瘟。其中危害最大的是叶瘟和穗颈瘟。近年,穗颈瘟已经成为黄淮稻区水稻生产的主要病害,年发病面积达66.7 万hm2以上,并且有逐年加重的危险。遗传研究学者已经从不同抗病品种中鉴定出80多个稻瘟病抗性基因,pib、pita、pi9、pi21、pikm、pb1等20多个抗稻瘟病基因已经被克隆,但是稻瘟病生理小种致病性变异频繁,单一抗性品种的抗性会在种植3~5 a后逐渐丧失,需要挖掘、聚合利用更多的广谱抗性基因才能选育更持久抗病的水稻品种[10]。
黄淮稻区常年粳稻种植面积约1.730×106hm2,约占我国粳稻种植面积的18.6%,是我国优质粳稻主产区之一。由于黄淮稻区地处我国南北过渡地带,气候生态条件年际间极不稳定,自然灾害发生频率较高,尤其是粳稻抽穗灌浆期经常遭遇低温阴雨寡照不良天气。低温阴雨一方面影响水稻灌浆结实,造成稻穗下部弱势粒灌浆不充分,稻米垩白粒率高、垩白度高[11-12];另一方面,低温阴雨又会导致稻瘟病高发频发,特别是穗颈瘟爆发,严重影响水稻产量和品质[13]。稻瘟病和稻米外观米质一直是影响黄淮粳稻发展的两大突出问题。稻瘟病属于致病生理小种多、变异频繁的病害,稻米外观米质的垩白性状属于多基因调控的复杂数量性状,所以稻米品质改良和抗病性同步提高的难度非常大,优质兼抗稻瘟病的粳稻品种一直十分缺乏。
2 黄淮稻区优质抗病粳稻“六位一体”复合育种技术体系的构建
针对黄淮稻区粳稻生产中长期存在抗稻瘟病和优质稻米品种缺乏的情况,自上个世纪90年代年始,河南农业大学、河南省农业科学院、新乡市农业科学院等河南省水稻育种优势单位,围绕优质抗病粳稻新品种选育开展了系统研究,在不断的实践和研究过程中总结出“六位一体”的复合育种技术体系。该育种技术体系是针对黄淮稻区粳稻生产和生态条件,集“早代大群体选择、多生态点联鉴、生理指标监测、分子标记辅助选择、稻米品质测定和抗病田间鉴定”六位一体(图1)。该育种技术体系的核心是早代大群体选择、稻瘟病抗病基因分子标记选择和田间多点鉴定相结合的抗稻瘟病育种技术和“籽粒相对充实度”为核心指标的灌浆速率快速监测稻米品质生理育种技术,该技术创新了稻米品质和抗病性的协同改良技术途径。
2.1 早代大群体选择
杂交育种后代选择过程中首要问题是如何确保亲本杂交重组出目标基因组合和性状,并且具有较高的出现几率,因此,确定育种群体大小与选择的概率至关重要。后代选留多少个体,一般育种家的计算方法为:n≥ log(1-a)/log(1-P),n是必须种植的个体数,P是群体中育种目标的基因组合或性状出现的概率,a是概率平推[14]。如果某目标性状是分别来自2个亲本的2对隐性基因控制,那么2个亲本杂交的F2群体出现目标性状的概率P=1/16。如果按照a=99%可靠概率平推,在F2至少种植71.35株,才能有99%的概率出现1株携带2对纯合隐性基因的目标植株。类推,如果是3对、4对或5对隐性基因来自不同的2个亲本,F2至少分别种植292.42株、1 176.62株或4 713.39株,才能有99%概率出现1株纯合目标植株。当然,这只是理想状态下保证了目标植株的出现,没有考虑其他因素的影响,如育种家能否把目标单株选择出来的概率。常规粳稻育种工作中,粳粳稻杂交后的F2群体,水稻育种家种植的群体个数一般为1 000株左右。对于1 000株的F2群体,在优化亲本组合、需要改良重组的目标基因和性状较少的情况下,能够出现目标植株。但是,稻米垩白性状是多基因调控的复杂数量性状,稻瘟病生理小种多变,稻瘟病抗性改良需要聚合多个主效广谱抗病基因。所以,稻米品质垩白性状改良和抗稻瘟病同步提高需要聚合多个品质和抗性相关的基因,F2后代群体必须种植足够的规模。在长期的优质抗病育种过程,应考虑育种试验面积、人力、物力等各方面的因素。具体做法是:首先利用200~300株小群体筛选优异的杂交组合,然后对优异组合F2群体种植5 000~6 000株的大群体,F2大群体确保4~5个目标基因的优异重组植株能够出现。此外,对优异组合的F3~F4世代保留100~200株,以提高优质和抗病基因优异重组出现的频率。F2世代材料多需逐株观察筛选。对于大规模的F2群体必需要有简便、迅速、经济、可靠的鉴定筛选方法。除了常规的株型、产量相关性状的肉眼观察筛选外,可在F2世代重点对稻瘟病抗性基因进行分子标记鉴定和筛选,以提高F2大群体筛选鉴定的准确性。
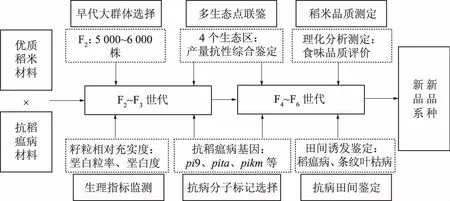
图1 “六位一体”复合育种技术体系
2.2 抗稻瘟病育种技术
由于稻瘟病病原菌小种遗传复杂易变,稻瘟病一直是我国各稻作区的主要病害。但是长期以来稻瘟病育种存在很多问题,比如个别主效广谱抗病基因过度使用,抗性下降甚至丧失;稻瘟病病原菌与水稻寄主间的关系及优势稻瘟病生理小种不清楚;常规稻瘟病抗病育种效率低、周期长,稻瘟病抗性与优质高产存在一定的矛盾[15]。因此,为了加快稻瘟病抗性育种的进度,使抗病与优质、高产更好的结合,在做好稻瘟病抗源引进和稻瘟病生理小种鉴定的基础上,应进一步加强完善育种过程的稻瘟病鉴定筛选体系,把稻瘟病抗病基因分子标记选择和田间多点鉴定相结合,从而提高抗病基因的选择效率,培育聚合多个主效广谱抗病基因的新品种。大量的稻瘟病菌种群分布及优势生理小种研究表明,黄淮稻区流行的稻瘟病菌代表菌株为ZG1、ZF1、ZD5、ZB17、ZC3、ZC5、ZB15等[16-19]。pi9、pikm、pita、pizt等基因为已经克隆或定位的主效稻瘟病抗性基因。pi9源自于小粒野生稻(Oryzaminuta),具有广谱抗性,对来自13个国家的43个稻瘟病菌株均表现出很高的抗性[20]。pikm是稻瘟病抗性基因pik上的一个主效抗病等位位点,编码产物是NBS-LRR类抗病蛋白[21]。pita起源于籼稻,在粳稻中表现出持久稳定的稻瘟病抗性[22]。pizt与水稻叶瘟抗性相关,与pi9、piz、pigm、pi2等互为等位的关系,对日本多个稻瘟病小种表现出较强的叶瘟抗性[23],因而这些基因被广泛用于稻瘟病抗性育种。
本科研团队利用pi9、pita、pikm、pizt、pib、pigm等稻瘟病抗性基因,设计筛选简单可靠的分子标记,在F2和F32个世代连续进行分子标记辅助筛选,在早代筛选出携带较多稻瘟病抗性基因的纯合植株。特别强调,稻瘟病分子标记辅助选择优先尽早进行,尽量在苗期移栽插秧前完成,这样减少插秧、田间种植和后期其他性状选择的工作量。另一方面,在F4~F6世代对相对稳定株系进行稻瘟病人工接种诱发鉴定,接种的稻瘟病生理小种为ZG1、ZF1、ZD5、ZB15等优势菌株,同时在豫南丘陵山区和江苏连云港赣榆稻瘟病重发区进行自然发病鉴定。这样通过低世代的稻瘟病抗性基因分子标记辅助选择和高世代田间自然及诱发多点鉴定,抗性基因的选择效率大大提高。培育的多个水稻品种都携带2~5个稻瘟病抗性基因,稻瘟病抗性都达到中抗以上水平(表1),表明该抗稻瘟病育种技术手段是切实可行的。
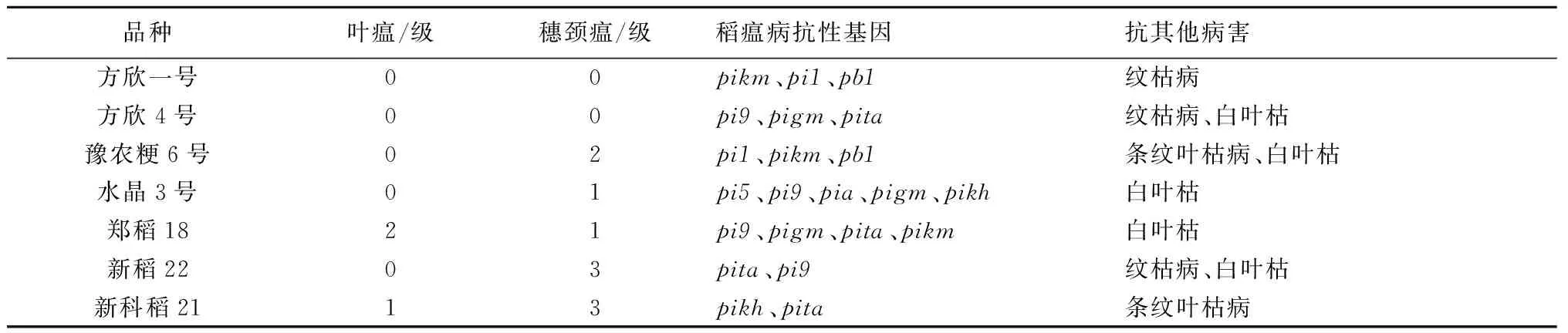
表1 部分育成水稻品种的抗病性及抗病基因
2.3 稻米品质生理育种技术
稻米品质是具有复杂遗传组成的综合性状,稻米品质的选择相比产量和株型等性状比较困难,且在早世代育种家很难做到准确鉴定。现代育种主要是基于对理想株型和产量指标的选择,而基于植株生理机能选择的生理育种可能是在获得理想株型基础上进一步提升产量和品质的一种途径[24]。水稻存在“两段灌浆”的生理现象,水稻生长中后期“二次灌浆”好的品种籽粒充实度往往较优。现代大穗型高产品种均普遍存在下部弱势粒籽粒充实度差的问题,水稻弱势粒籽粒充实度差是限制现代水稻品种品质和产量潜力进一步提升的瓶颈。水稻籽粒充实度差的品种基本表现为强势粒和弱势粒的异步灌浆[25]。大量研究也表明,花后5~12 d,籽粒相对充实度迅速降低;花后12~19 d迅速上升,花后9~32 d 缓慢上升,籽粒灌浆呈先降后升的偏V型曲线。灌浆中后期(花后19~32 d),籽粒相对充实度与千粒质量和产量呈显著正相关,但是与强势粒和弱势粒的垩白粒率、垩白度、垩白面积呈显著负相关,与弱势粒的糙米率呈显著正相关[26-27]。所以,增强灌浆中后期弱势粒的灌浆速率,降低灌浆中后期弱势粒与强势粒之间的灌浆差异,可以提高弱势籽粒的相对充实度,从而增加水稻产量、改善稻米品质,尤其是可以达到改善稻米垩白粒率和垩白度等外观品质的目的。
基于以上研究基础,提出了以“籽粒相对充实度”为核心指标的灌浆速率快速监测生理育种技术,用籽粒相对充实度对目标F2~F3世代植株进行筛选,选择生长中后期下部弱势籽粒相对充实度高的株系,减少品种的垩白粒率和垩白度。实践中,河南农业大学、新乡市农业科学院、河南省农业科学院等单位以“籽粒相对充实度”为筛选指标选择了方欣一号、水晶3号、新稻22等品种,这些品种生长中后期的籽粒灌浆比较充分,下部弱势籽粒相对充实度高,稻米的垩白粒率、垩白面积较少,米质都达到国标3级以上(表2)。说明籽粒相对充实度可以反映水稻生长中后期2次灌浆的灌浆速率,可用来鉴别稻米外观品质,作为育种家筛选稻米品质的一个生理参数。需要注意的是,籽粒相对充实度的筛选与稻瘟病分子标记选择是紧密结合的,一定要在分子标记筛选确定抗病目标候选植株/株系的基础上,再利用相对充实度筛选获得稻米品质优良的材料,以减少工作量,达到事半功倍的效果。
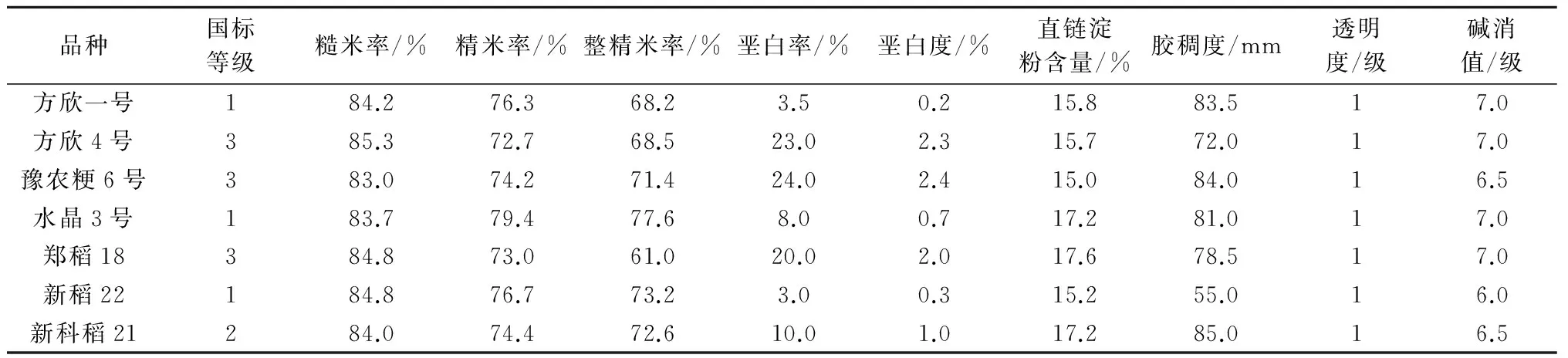
表2 部分育成水稻品种的稻米品质指标
3 “六位一体”复合育种技术体系的实践及成效
自上世纪90年代年始,按照“六位一体”优质抗病粳稻复合育种技术体系的技术思路,河南农业大学、新乡市农业科学院、河南省农业科学院等水稻育种单位先后选育了方欣一号、方欣4号、豫农粳6号、水晶3号、郑稻18、新稻22、新科稻21等一批优质抗病粳稻新品种,其中达国标1级优质米品种3个、2级优质米1个、3级优质米3个,稻瘟病全部达中抗以上,大田产量也都达到7 240 kg/hm2以上(表3)。“六位一体”复合育种技术在粳稻优质和抗稻瘟病两大性状有效聚合上具有很强的优势,选育的品种经济、生态和社会效益显著,有力地推动了黄淮稻区水稻科技进步和优质稻产业的发展,简介如下。
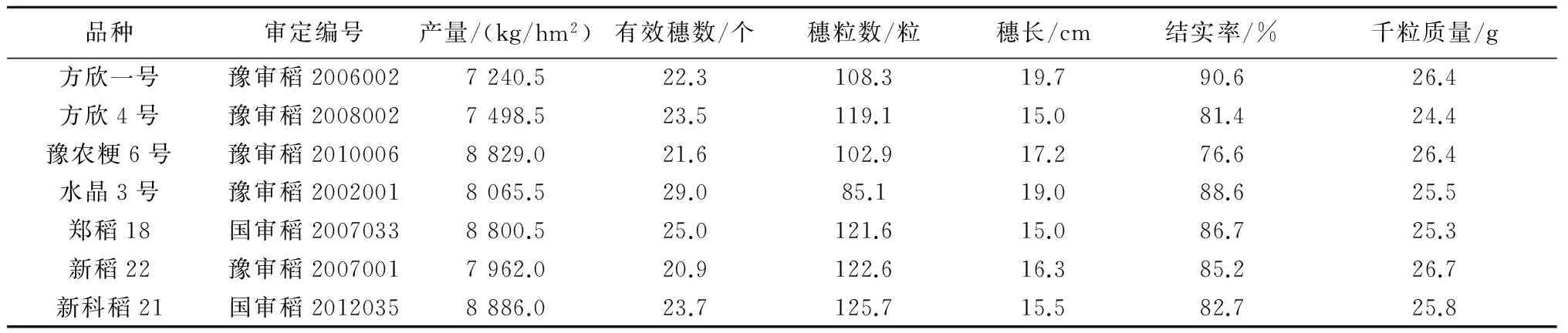
表3 部分育成水稻品种的产量性状指标
3.1 方欣一号
河南省审定。经农业部食品质量监督检验测试中心(武汉)2004—2005年测试分析,该品种品质均达国家优质米标准1级。糙米率84.1%,精米率76.3%,整精米率68.2%,垩白粒率2.0%,垩白度0.1%,直链淀粉含量15.8%,透明度1.0级,碱消值7级,胶稠度83.5 mm。第三届中国优质稻米博览会上,被评为“中国十大金奖大米”第1名。具有稻瘟病广谱抗病基因pikm、pi1和pb1,对稻瘟病菌代表小种菌株均表现为免疫,对水稻穗颈瘟表现为免疫,对白叶枯病2个致病型菌株浙173、PX079表现为抗病(1级),对KS-6-6、JS-49-6表现为感病,对纹枯病表现为抗病。
3.2 方欣4号
河南省审定。经农业部食品质量监督检验测试中心(武汉)分析,该品种品质达国家优质米标准3级。糙米率85.3%,整精米率68.5%,垩白粒率23%,垩白度2.3%,直链淀粉15.7%,胶稠度72 mm,气味香,口感食味佳。具有稻瘟病广谱抗病基因pi-ta、pi9和pigm,对稻瘟病菌种ZB13、2A5、2C5、ZE3、ZF1、ZG1、ZD5表现为免疫,对穗颈瘟高抗;对白叶枯菌株PX079、JS-49-6表现中抗,对浙173表现为中感,对KS-6-6表现为抗病,对纹枯病表现为抗。
3.3 豫农粳6号
河南省审定。经农业部食品质量监督检验测试中心(武汉)分析,该品种品质达国家优质米标准3级。糙米率83.0%,整精米率71.4%,垩白粒率30%,垩白度2.4%,直链淀粉15.0%,胶稠度84 mm。具有稻瘟病广谱抗病基因pi1、pb1和pikm,对多个稻瘟病菌代表菌株ZG1、ZC15、ZD7、ZE3、ZB10、ZF1均表现为免疫或中抗,中抗穗颈瘟;对白叶枯病5个代表菌株表现为抗病。
3.4 水晶3号
河南省审定。经农业部食品质量监督检验测试中心(武汉)分析,该品种品质达国家优质米标准1级。糙米率83.7%,整精米率77.6%,垩白粒率8%,垩白度0.7%,直链淀粉17.2%,胶稠度81 mm。具有稻瘟病广谱抗病基因pi5、pi9、pia、pikh、pigm,抗稻瘟病和白叶枯病。
3.5 郑稻18
国家审定。经农业部食品质量监督检验测试中心(武汉)分析,该品种品质达国家优质米标准3级。糙米率84.8%,整精米率61%,垩白粒率20.0%,垩白度2.0%,直链淀粉含量17.6%,胶稠度78.5 mm。具有稻瘟病广谱抗病基因pi9、pigm、pita、pikm,对稻瘟病菌代表小种ZC13、ZF1免疫(0级);抗水稻穗颈瘟病(1级),综合抗性指数3.3;对白叶枯病2个致病型菌株PX079、JS-49-6表现为抗,对菌株浙173、KS-6-6表现为中抗。
3.6 新稻22
河南省审定。经农业部食品质量监督检验测试中心(武汉)分析,该品种品质达国家优质米标准1级。糙米率84.8%,精米率76.7%,整精米率73.2%,垩白粒率3%,垩白度0.3%,直链淀粉15.2%,胶稠度55 mm,粒型长宽比1.8,透明度1级,碱消值6.0级。具有稻瘟病广谱抗病基因pita和pi9,对叶瘟6个小种的代表菌株ZB15、ZC15、ZD1、ZE1、ZF1、ZG1表现为抗病(0级),对穗颈瘟表现为中抗(3级);对白叶枯病代表菌株浙173、JS-49-6、PX079均表现较好的抗性,仅对白叶枯病病菌KS-6-6 表现为中感(5级);对纹枯病表现为抗病。
3.7 新科稻21
国审。经农业部食品质量监督检验测试中心(武汉)分析,该品种品质达国家优质米标准2级。糙米率84.0%,整精米率72.6%,垩白粒率10.0%,垩白度1.0%,直链淀粉含量17.2%,胶稠度85 mm。具有稻瘟病广谱抗病基因pikh和pita,稻瘟病综合抗性指数3.4,穗颈瘟损失率最高级3级,条纹叶枯病最高发病率3.96%,中抗稻瘟,高抗条纹叶枯病。
[1] 莫惠栋.我国稻米品质的改良[J].中国农业科学,1993,26(4):8-14.
[2] 张昌泉,赵冬生,李钱峰,等.稻米品质性状基因的克隆与功能研究进展[J].中国农业科学,2016,49(22):4267-4283.
[3] Che R,Tong H,Shi B,etal.Control of grain size and rice yield by GL2-mediated brassinosteroid responses[J].Nature Plants,2015,2:15195.
[4] Bridgemohan P,Bridgemohan R S H.Crop nutrition studies on grain filling and chalkiness in rice[J].Journal of Plant Breeding and Crop Science,2014,6(10):144-152.
[5] Wang E,Wang J,Zhu X,etal.Control of rice grain-filling and yield by a gene with a potential signature of domestication[J].Nature Genetics,2008,40(11):1370-1374.
[6] Kang H G,Park S,Matsuoka M,etal.White-core endosperm floury endosperm-4 in rice is generated by knockout mutations in the C4-type pyruvate orthophosphate dikinase gene(OsPPDKB)[J].The Plant Journal,2005,42(6):901-911.
[7] Li Y,Fan C,Xing Y,etal.Chalk5 encodes a vacuolar H+-translocating pyrophosphatase influencing grain chalkiness in rice[J].Nature Genetics,2014,46(4):398-404.
[8] Wang Z,Li H,Liu X,etal.Reduction of pyruvate orthophosphate dikinase activity is associated with high temperature-induced chalkiness in rice grains[J].Plant Physiology and Biochemistry,2015,89:76-84.
[9] 孙国昌,杜新法.水稻稻瘟病防治策略和21世纪研究展望[J].植物病理学报,1998,28(4):289-292.
[10] 王生轩,李俊周,谢瑛,等.河南粳稻抗稻瘟病基因pi9、pita和piz-t的分子检测[J].分子植物育种,2017,15(3):951-955.
[11] 杜彦修,季新,张静,等.弱光对水稻生长发育影响研究进展[J].中国生态农业学报,2013,21(11):1307-1317.
[12] 马爱婷.低温阴雨对早稻结实率的影响及防御对策[J].作物研究,1995,9(1):17-19.
[13] 司兆胜,宫香余,李鹏.水稻稻瘟病流行因素及绿色防控措施[J].北方水稻,2011,41(5):41-43.
[14] 俞世蓉,沈克全.作物繁殖方式和育种方法[M].北京:中国农业出版社,1996
[15] 鄂志国,张丽靖,焦桂爱,等.稻瘟病抗性基因的鉴定及利用进展[J].中国水稻科学,2008,22(5):533-540.
[16] 刘永锋,陈志谊,胡明,等.江苏省稻瘟病菌群体分布及优势小种的毒力研究[J].中国水稻科学,2004,18(4):351-356.
[17] 肖丹凤,张佩胜,王玲,等.中国稻瘟病菌种群分布及优势生理小种的研究进展[J].中国水稻科学,2013,27(3):312-320.
[18] 徐建第,张全芳,姜明松,等.2013 年山东省部分稻区稻瘟病菌生理小种的鉴定及分析[J].山东农业科学,2015,47(4):110-111.
[19] 陈莉,苏贤岩,丁克坚.安徽稻区稻瘟病菌生理小种的鉴定[J].中国农学通报,2007,23(4):302-306.
[20] Liu G,Lu G,Zeng L,etal.Two broad-spectrum blast resistance genes,pi9(t) andpi2(t),are physically linked on rice chromosome 6[J].Molecular Genetics and Genomics,2002,267(4):472-480.
[21] Costanzo S,Jia Y.Sequence variation at the rice blast resistance genepi-kmlocus:implications for the development of allele specific markers[J].Plant Science,2010,178(6):523-530.
[22] Huang C L,Hwang S Y,Chiang Y C,etal.Molecular evolution of thepi-tagene resistant to rice blast in wild rice(Oryzarufipogon)[J].Genetics,2008,179(3):1527-1538.
[23] Zhou B,Qu S,Liu G,etal.The eight amino-acid differences within three leucine-rich repeats betweenpi2 andpiz-tresistance proteins determine the resistance specificity toMagnaporthegrisea[J].Molecular Plant-microbe Interactions,2006,19(11):1216-1228.
[24] 永锐.水稻生理育种[M].北京:科学技术文献出版社,1995.
[25] 谢光辉,杨建昌,王志琴,等.水稻籽粒灌浆特性及其与籽粒生理活性的关系[J].作物学报,2001,27(5):557-565.
[26] 赵全志,吕强,殷春渊,等.大穗型粳稻籽粒相对充实度的化学调控及其与产量和品质的关系[J].作物学报,2006,32(10):1485-1490.
[27] 赵全志,殷春渊,宁慧峰,等.氮素调控对水稻子粒相对充实度的影响及子粒相对充实度与产量形成和品质的关系[J].河南农业大学学报,2007,41(2):128-133.
