同一养殖环境下不同规格三疣梭子蟹竞食能力的比较研究
2017-12-18史会来许文军王庚申谢建军
何 杰,史会来,许文军,王庚申,汪 玮,施 慧,谢建军
( 1.浙江省海洋水产研究所,浙江省海水增养殖重点实验室,浙江 舟山 316021;2.浙江海洋大学,浙江 舟山 316021 )
同一养殖环境下不同规格三疣梭子蟹竞食能力的比较研究
何 杰1,2,史会来1,许文军2,王庚申1,汪 玮1,2,施 慧1,谢建军1
( 1.浙江省海洋水产研究所,浙江省海水增养殖重点实验室,浙江 舟山 316021;2.浙江海洋大学,浙江 舟山 316021 )
将3种不同规格的三疣梭子蟹[平均体质量分别为(101.26±5.36) g、(62.37±4.10) g和(24.37±2.10) g]各10只饲养于2.6 m×1.8 m×1 m的室内水泥池单元格内,水深40 cm,重复3组。每日17:00投喂鲜活的带鱼块,通过摄像机观察不同规格三疣梭子蟹的竞食行为,每组重复3次。结果表明:(1)不同规格三疣梭子蟹发现、抱持食物所需的时间相近,无显著差异(P>0.05);(2)个体间竞食激烈,(101.26±5.36) g组蟹抢食的比率最高,抢夺(62.37±4.10) g组或(24.37±2.10) g组蟹所持有的食物;(101.26±5.36) g组蟹抢食的次数显著多于(62.37±4.10) g组或(24.37±2.10) g组蟹的抢食次数(P<0.05),(24.37±2.10) g组蟹抢食的次数最少;(24.37±2.10) g组蟹被(101.26±5.36) g、(62.37±4.10) g、(24.37±2.10) g组蟹抢食的次数及总次数最高,显著多于(101.26±5.36) g组蟹(P<0.05),(62.37±4.10) g组蟹次之,(101.26±5.36) g组蟹最少,后两组蟹无显著差异(P>0.05),表现出明显的“大欺小”现象;(3)(101.26±5.36) g组蟹的食物占有率(45%)显著高于(24.37±2.10) g组蟹(近20%)(P<0.05),而(62.37±4.10) g组蟹占有的食物数量介于(101.26±5.36) g组和(24.37±2.10) g组之间,(101.26±5.36) g组蟹啃食完食物所用的时间仅约为(24.37±2.10)g组蟹的1/5倍。
三疣梭子蟹;规格;竞争;食物
当同种或异种动物对有限的公共资源有共同需要时就会发生种内或种间的竞争。竞争的主要目标包括食物、窝巢或配偶等,而对食物的竞争最为常见[1]。目前,国内外有关动物竞争食物的研究主要见于陆上昆虫、鼠类以及部分大型哺乳动物[2-3],而有关水生动物的竞食效应研究较少,且主要集中于自然水域条件下鱼、蟹、虾类的种间食物竞争关系研究等[4-5],有关人工养殖条件下的种内或种间食物竞争研究极为鲜见。
虾蟹类甲壳动物依靠蜕壳(或蜕皮)实现体型的增大和体质量的增加,呈现出阶梯式的生长模式。由于自身遗传和社群控制的作用,即使在同一环境下,同一群体的虾蟹类蜕壳也难以同步,致使其在养殖中后期规格分化现象极其严重[6-8]。然而,迄今为止,尚未见有关虾蟹类规格分化(个体大小差异)对其种内竞争食物的行为能力研究。鉴于此,本文以生性好斗、领域性极强的海水养殖的三疣梭子蟹(Portunustrituberculatus)为对象,初步探讨了在同一养殖环境下规格分化对竞食能力的影响,旨在为丰富甲壳动物的行为生态学、种群生态学和养殖生态学资料,为三疣梭子蟹的科学养殖提供参考。
1 材料与方法
1.1 材料
本试验于2016年8月在浙江省海洋水产研究所西闪岛试验场进行。在池塘养殖三疣梭子蟹去雄期间,从池塘内诱钓、挑选十足健全的雄梭子蟹200余只运回实验室,暂养于8 m×1.8 m×1 m的室内水泥池,水深40 cm,连续充氧,每日17:00投喂鲜活带鱼(Trichiuruslepturus)块,次日8:00清除残饵,换水1/3。暂养3 d后,从中挑选相对较大规格(101.26±5.36) g、中等规格(62.37±4.10) g和较小规格(24.37±2.10) g的蟹各30只用于正式试验。
1.2 方法
选1口8 m×1.8 m×1 m的室内水泥池,用渔网、砖块等材料等分成3个2.6 m×1.8 m×1 m的单元格,单元格间的水流相通,水深40 cm,连续充氧。在各单元格上空中心位置安装一不锈钢支架,用于固定摄像机(索尼HDR-PJ410)。之后向各单元格内放养(101.26±5.36) g组、(62.37±4.10) g组、(24.37±2.10) g组三疣梭子蟹各10只,每日17:00投喂鲜活的带鱼块,次日8:00清除残饵,换水1/2,试验期间的水温(28±2) ℃,pH 8.8±0.1,溶解氧>5 mg/L。4 d后,待蟹基本适应试验环境后开始正式试验。试验期间,调准好摄像机焦距后,向单元格中间区域投喂10块(份)大小相近(1.52±0.13) g的带鱼块,待所有蟹啃食完毕后关闭摄像机(15~30 min),按同样方法每日依次记录3个单元格内蟹的摄食和竞食行为,持续3 d,即整个试验重复9次。拍摄结束后,使用联想G425笔记本电脑读取摄像机SD卡内的影像资料,统计每次摄食期间蟹索食所用时间、鱼块被不同蟹的占有率、啃完食物所用时间,并从每个重复中随机挑选6~8只蟹,分别记录每只蟹在整个啃食过程中被同类抢食的次数、是否被成功抢走以及其携带食物更换空间位置(以水平位移大于20 cm为标准)的次数等。
1.3 数据处理
试验数据用平均值±标准差表示,不同试验组数据间采用单因素方差分析,对检测达到显著水平的均值用Duncan检验,用SPSS 11.5进行方差分析和多重比较,在Sigmaplot 10.0下绘制相关图表。
2 结 果
2.1 不同规格三疣梭子蟹索食所需的时间
投喂食物后,蟹反应积极,索食行为明显,一旦发现食物,用螯足迅速抱持食物,送入口中,第一大颚不断切碎和磨损食物,第一和第二对颚足护住食物,以防流失。由图1可见,不同规格三疣梭子蟹发现并抱持食物所需的时间均约为0.28 min,差异不显著(P>0.05)。
2.2 不同规格三疣梭子蟹抢食的行为特征
试验期间投喂的饵料不足,同类之间发生了激烈的争夺食物行为。其行为可分为5个阶段:(1)相遇阶段,双方身体姿态不变,没有额外使用附肢进攻或防御(图2a);(2)示威阶段,抢食者向食物持有者靠近,抬高身体、展开大螯向对方示威,食物持有者用其中一个螯足携持食物,高举另一螯足防御,并试图侧身而逃(图2b,c);(3)抢夺阶段,抢食者直接用螯足抢夺对方所持有的食物,双方均用螯足夹住食物,互不相让(图2d);(4)拳(螯)击阶段,双方不断使用大螯推挤,并努力翻转或扭曲对方的螯足,企图损伤或卸去对方的螯足(图2e,f),双方打斗极为激烈,水波的干扰致拍摄的照片较为模糊;(5)撤退阶段,食物完全被对方抢走后,失败方撤离,抢夺结束(图2g,h)。

图1 不同规格三疣梭子蟹索食所需的时间
40%~80%持有食物的试验蟹曾被(101.26±5.36) g组蟹抢夺,35%~60%的个体曾被(62.37±4.10) g组蟹抢夺,20%~50%的个体曾被(24.37±2.10) g组蟹抢夺,被(101.26±5.36) g组蟹抢食的比率最高,被(62.37±4.10) g组蟹抢食的比率次之,被(24.37±2.10) g组蟹抢食的比率最低,说明大规格蟹表现出最明显的主动抢食行为;另外,(101.26±5.36) g组蟹更倾向于抢夺(62.37±4.10) g或(24.37±2.10) g组蟹所持有的食物,而(24.37±2.10) g组蟹更多地选择向与其规格相近的(24.37±2.10) g组蟹发起抢食攻击,但其中也有部分个体向(101.26±5.36) g和(62.37±4.10) g组蟹抢夺食物,其比率显著小于向(24.37±2.10) g组蟹抢食的比率(P<0.05)(图3)。各组蟹被同类抢食的次数相似(图4),即(101.26±5.36) g、(62.37±4.10) g、(24.37±2.10) g组蟹均被(101.26±5.36) g组蟹抢食的次数最多,被(24.37±2.10) g组蟹抢食的次数最少,其中(101.26±5.36) g和(24.37±2.10) g组蟹被(101.26±5.36) g组蟹抢食的次数显著多于被(62.37±4.10) g或(24.37±2.10) g组蟹抢食的次数(P<0.05);而各组蟹之间相比,(24.37±2.10) g组蟹分别被(101.26±5.36) g、(62.37±4.10) g、(24.37±2.10) g组蟹抢食的次数及总次数均为最多,(62.37±4.10) g组蟹次之,(101.26±5.36) g组蟹最少,且(24.37±2.10) g组蟹被同类抢食的次数显著多于(101.26±5.36) g组蟹(P<0.05),(62.37±4.10) g组蟹与(101.26±5.36) g组和(24.37±2.10) g组蟹均无显著差异(P>0.05)。
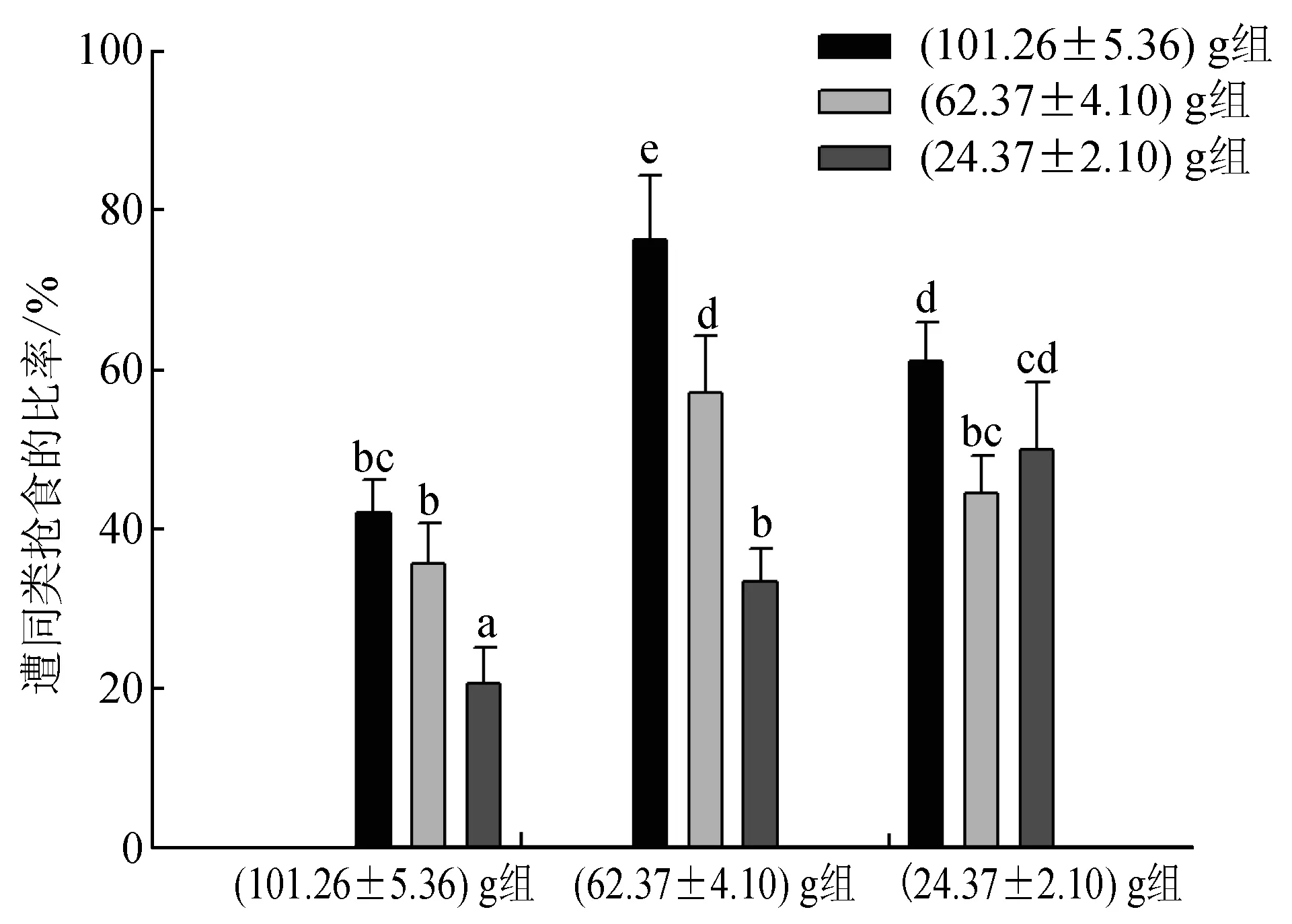
图3 不同规格三疣梭子蟹遭同类抢食的比率同一系列上标有相同字母表示差异不显著(P>0.05),标有相异字母表示差异显著(P<0.05),下同.
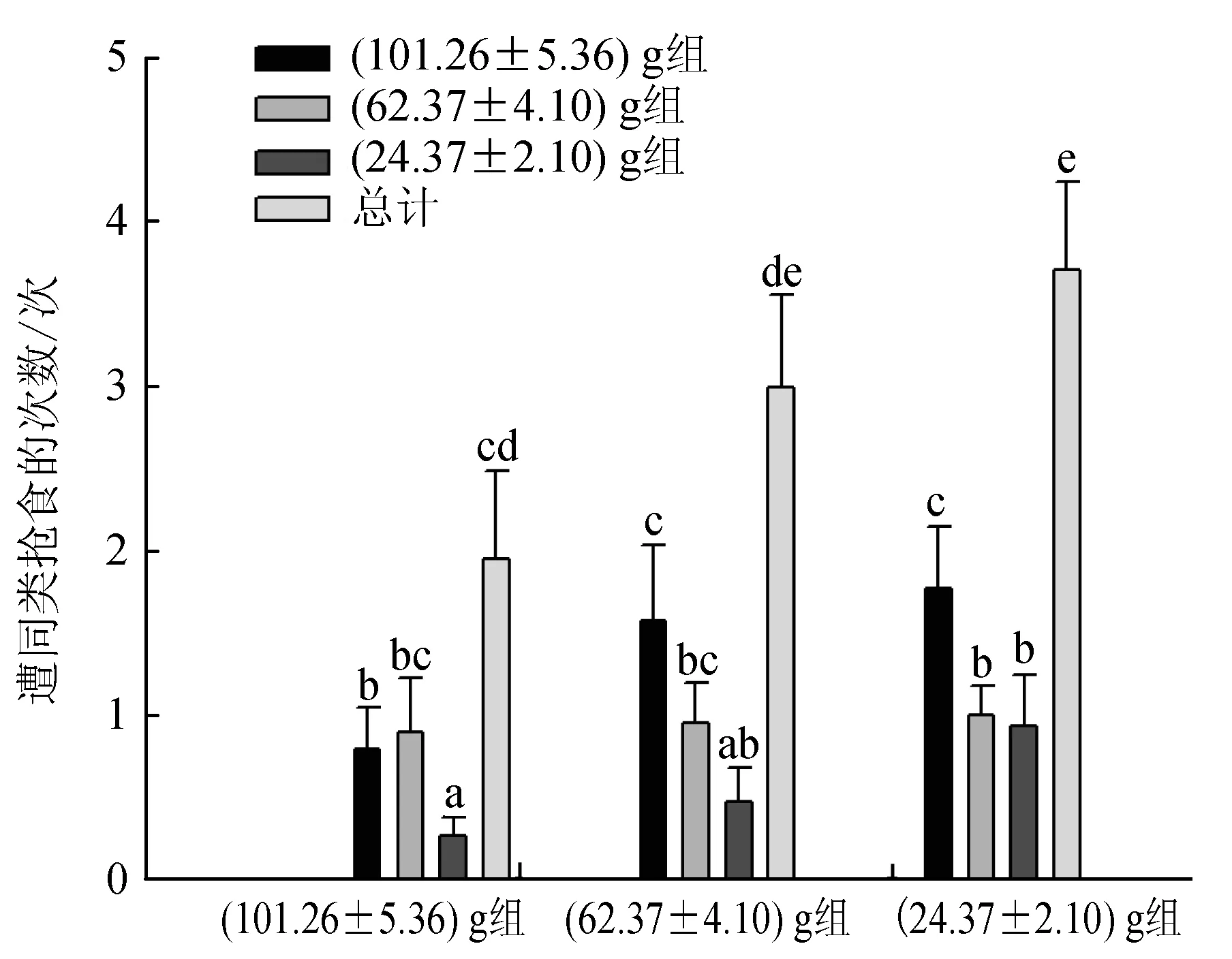
图4 不同规格三疣梭子蟹遭同类抢食的次数
三疣梭子蟹在啃食期间不停更换空间位置来降低食物被同类发现或被抢夺的概率,其中(24.37±2.10) g组蟹最为明显,在整个啃食过程中更换了约10次空间位置,显著高于(62.37±4.10) g组(约6次)和(101.26±5.36) g组(约4次)蟹(P<0.05)(图5)。无论是(101.26±5.36) g、(62.37±4.10) g还是(24.37±2.10) g组,三疣梭子蟹所持有的食物均可能被同类抢走。(24.37±2.10) g组蟹中50%的个体所持有的食物被同类抢走,显著高于(62.37±4.10) g组(约30%)和(101.26±5.36) g组(约10%)的蟹(P<0.05)(图6)。而且(24.37±2.10) g组蟹所持食物均可能被任何蟹成功抢夺,被(101.26±5.36) g组蟹抢走的比率最高,被(24.37±2.10) g组蟹抢走的比率最低,而(101.26±5.36) g组的蟹仅能被与其规格相近的(101.26±5.36) g组蟹抢走,不能被比其规格小的(62.37±4.10) g组和(24.37±2.10) g组蟹抢走,同样,(62.37±4.10) g组蟹所持有的食物仅能被(101.26±5.36) g组和(62.37±4.10) g组的蟹抢走,不可能被(24.37±2.10) g组的蟹抢走。
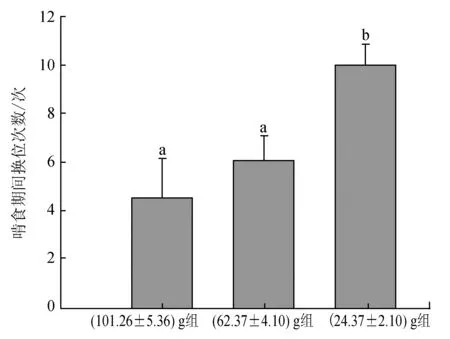
图5 不同规格三疣梭子蟹在啃食期间身体移动次数
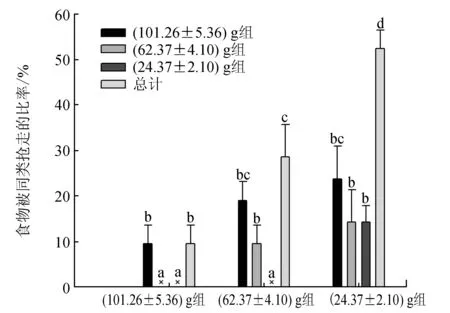
图6 不同规格三疣梭子蟹所持食物被同类抢走的比率
2.3 不同规格三疣梭子蟹对有限食物的占有率及其啃食所需时间
不同规格三疣梭子蟹对有限食物的占有率不同,其中(101.26±5.36) g组三疣梭子蟹占有率最高,侵占了约45%的食物,显著高于(24.37±2.10) g组蟹(近20%)(P<0.05),而(62.37±4.10) g组蟹占有的食物数量介于(101.26±5.36) g和(24.37±2.10) g之间(图7)。啃食完食物所用的时间越短,啃食速度越快,花费时间越短,小规格蟹啃食所需时间约为大规格蟹的5倍(图8)。

图7 不同规格三疣梭子蟹对食物的占有率

图8 不同规格三疣梭子蟹啃食食物所需的时间
3 讨 论
动物种内竞争主要表现在同种对食物、生存空间、配偶等的争夺与保卫[1]。水生动物的种内竞争强度由各种错综复杂的因素综合引起,栖息环境恶劣、个体间大小悬殊、饵料不足以及种群拥挤等约能加剧种内竞争行为[9-10],在高密度、封闭的人工饲养环境下,养殖动物在获取有限的生物资源时更易发生激烈的竞争[11]。养殖动物通过摄取食物来获得能量,在饵料投喂不充足条件下,动物种内的食物竞争直接关系到个体的生存和生长,影响养殖效益,因此,探讨养殖条件下水生经济动物种内食物竞争行为的发生机制以及调节措施,对养殖对象的稳产和高产具有直接的指导意义。
虾蟹类理想的养殖效果可概括为“两高一短”,即“成活率高,蜕壳同步率高,蜕壳周期短”[12]。Segal等[13]研究发现,单个个体隔离养殖的罗氏沼虾(Macrobrachiumrosenbergii)的蜕壳同步率明显高于群体混养模式,认为直接的食物竞争是导致部分个体生长快速,部分个体被抑制生长的主要原因;董鹏生等[8]发现,即使是同一家系的中华绒螯蟹(Eriocheirsinensis)在同一生境下群养后,蜕壳仍然非常不同步,随着养殖时间的延长,规格差异日益显现,收获时的大、小规格个体的体质量差异可达10倍以上,甚至部分个体生长过快出现早熟现象。可见,除甲壳动物自身遗传背景外,社群控制是导致甲壳动物规格严重分化的主要诱因之一,而且种内竞争(主要指食物和生存空间)是社群控制的主要表现形式。
本研究中,不同或同一规格间的三疣梭子蟹均表现出激烈的种内竞食行为,其行为大致可分为5个阶段:相遇、示威、抢夺、拳击和撤退阶段,整个竞食过程与先前报道的小龙虾的争胜行为相似[14]。笔者认为,这是甲壳动物个体间格斗、竞争资源时通用的行为模式。不同规格三疣梭子蟹被同类抢食的比率和次数表明,大规格个体的抢食欲望强于小规格个体,各规格蟹均趋向抢食比其自身规格小的个体,表现出明显的“大欺小”的行为特征,这与罗氏沼虾的种内竞争研究结果一致[15]。而小规格蟹只能通过不断的更换空间位置,尽量回避竞争者的抢食行为。这是动物在应对危险环境下做出的一种规避性本能反应,也是本研究中小规格蟹啃食完相同大小食物所需时间远大于大规格蟹的原因之一。本研究还发现,小规格三疣梭子蟹也能向比其自身大的个体进行攻击性的抢夺食物,但比率较低,其更多的选择与其规格相近的小规格个体抢食。从食物被同类抢走的比率以及各规格蟹对食物的最终占有率来看,大规格个体的抢食能力也明显强于小规格个体,致使其获得更多的食物资源,所以,在食物不充足的情况下,小规格三疣梭子蟹面临饥饿的可能性更大。众多研究已表明,饥饿胁迫不仅直接抑制甲壳动物成活和蜕壳成功率,还不同程度地影响蜕壳后的质量增加率和蜕壳周期,加剧了养殖群体的规格分化[16-17],种内食物竞争越趋激烈,以此恶性循环的发展,最终导致养殖动物成活率和养殖产量低下[6,18]。
规格分化将加剧甲壳动物种内竞争食物行为的发生,使部分个体生长摄入营养过多,生长过快而早熟,部分个体摄入营养不足,生长滞后甚至停止生长。因此,要不断改进养殖设施和技术,降低种群内规格分化的程度,减少种内对食物、生存空间的竞争。近年来,三疣梭子蟹单个体养殖技术应运而生。该技术可大大提高三疣梭子蟹的成活率,但由于投饵、去残饵的自动化管理技术配套不够等,没有得到大范围的推广,所以,今后的研究应加强三疣梭子蟹单个体养殖配套技术的开发。针对现有的池塘群养技术,应通过合理的放养密度、均匀投饵等措施,尽可能减少种内对食物的竞争,提高养殖成活率和规格整齐度。
[1] 尚玉昌. 普通生态学[M]. 北京:北京大学出版社, 2010:173-174.
[2] 杨效文. 麦蚜种间竞争研究初报 [J]. 生态学杂志, 1991,10(1):1-5.
[3] Farjana T, Tuno N, Higa Y, et al. Effects of temperature and diet on development and interspecies competition inAedesaegyptiandAedesalbopictus[J]. Medical and Veterinary Entomology, 2012, 26(2):210-217.
[4] 张波, 唐启升, 金显仕, 等. 东海和黄海主要鱼类的食物竞争[J]. 动物学报, 2005, 51(4):616-623.
[5] 郭斌, 张波, 戴芳群, 等. 海州湾小黄鱼幼鱼和黄鲫幼鱼的食物竞争[J]. 渔业科学进展, 2011, 32(1):8-15.
[6] Gebauer P, Paschke K, Anger K. Delayed metamorphosis in decapod crustaceans:evidence and consequences[J]. Revista Chilena de Historia Natural,2003,76(2):169-175.
[7] He J, Gao Y, Xie J J, et al. Limb autotomy patterns in the juvenile swimming crab (Portunustrituberculatus) in earth ponds[J]. Aquaculture,2016,463(3/4):189-192.
[8] 董鹏生, 刘青,吴旭干, 等.中华绒螯蟹一龄性早熟和二龄成熟家系扣蟹阶段生长和早熟的比较研究[J].上海海洋大学学报,2016,25(1):51-60.
[9] Thomas C W, Carter C G, Crea B J. Feed availability and its relationship to survival, growth, dominance and the agonistic behaviour of the southern rock lobster,Jasusedwardsiiin captivity[J]. Aquaculture,2003, 215(1/4):45-65.
[10] 陈学雷, 林琼武, 李少菁, 等.日本对虾仔虾相残的实验研究[J]. 厦门大学学报:自然科学版,2003,42(3):358-361.
[11] 李玉全, 孙霞. 水生动物的争胜行为[J]. 动物学研究, 2013,34(3):214-220.
[12] Figueiredo J, Lopes G P, Narciso L, et al. Effect of starvation during late megalopa stage ofMithraculusforceps(Brachyura:Majidae) on larval duration, synchronism of metamorphosis, survival to juvenile, and newly metamorphosed juvenile size[J]. Aquaculture, 2008,274(1):175-180.
[13] Segal E, Roe A. Growth and behaviour of post-juvenileMacrobrachiumrosenbergii(de Man) in close confinement[J].Proceedings of the World Mariculture Society, 1975, 6(1/4):67-88.
[14] Moore P A. Agonistic behavior in freshwater crayfish——the influence of intrinsic and extrinsic factors on aggressive encounters and dominance[G]//Duffy J E, Thiel M. Evolutionary Ecology of Social and Sexual Systems-Crustaceans as Model Organisms, New York:Harvard University Press, 2007:90-114.
[15] Karplus I. Social control of growth inMacrobrachiumrosenbergii(De Man):a review and prospects for future research[J]. Aquaculture Research, 2005, 36(3):238-254.
[16] 吴立新, 董双林, 姜志强.饥饿对甲壳动物生理生态学影响的研究进展[J].应用生态学报, 2004, 15(4):723-727.
[17] 何杰,王春,邵路畅,等.饥饿胁迫对中华绒螯蟹(Eriocheirsinensis)仔蟹的影响[J].生态学杂志, 2013,32(8):2077-2084.
[18] Polis G A. The evolution and dynamics of intraspecific predation[J].Annual Review of Ecology and Systematics,1981,12(12):225-251.
ComparisonofCompetitionforFoodamongSwimmingCrabPortunustritubercatuswithVariousSizesinSameHabitat
HE Jie1,2, SHI Huilai1, XU Wenjun2, WANG Gengshen1, WANG Wei1,2, SHI Hui1, XIE Jianjun1
( 1. Key Laboratory of Marine Culture and Enhancement of Zhejiang Province, Marine and Fisheries Research Institute of Zhejiang Province, Zhoushan 316021, China; 2. Zhejiang Ocean University, Zhoushan 316021, China )
A total of 30 swimming crabPortunustrituberculatuswith three different sizes [average body weight of (101.26±5.36) g (LC), (62.37±4.10) g (MC) and (24.37±2.10) g (SC)] were raised in the same indoor tanks of each 2.6 m×1.8 m×1 m with water depth of 40 cm with triplication. After crabs were fed fresh ribbon fish at 17:00 daily, camera was used to record their eating and competing behavior for food with triplication. The results showed that swimming crab with different body weight had similar foraging time for food (P>0.05). Intraspecific competition for food among swimming crab individuals was highly fierce and the behaviors were approximately divided into five stages:meet stage, demonstration stage, snatch stage, pugilism stage, and withdraw stage. The proportion and frequency of competition for food exhibited that the larger-sizes individuals had a stronger competition desire and were inclined to compete for food with smaller individuals. As for the proportion of being deprived of food and the ultimate occupancy of food, smaller-size individuals had relatively poorer capability of invasion and protection for food.
Portunustritubercatus; size;competition;food
10.16378/j.cnki.1003-1111.2017.06.008
S968.25
A
1003-1111(2017)06-0741-06
2016-10-10;
2017-01-16.
浙江省科技厅院所专项(2016F50042, 2015F10001, 2015F30003);浙江海洋大学博士启动基金资助项目(22135010815 );舟山市科技计划项目(2017C43032);舟山市普陀区科技计划项目(2015YN209).
何杰(1985-),男,博士;研究方向:虾蟹类健康养殖.E-mail:he_0902@126.com. 通讯作者:许文军(1971-),男,教授;研究方向:海水健康养殖,E-mail:wjxu1971g@hotmail.com.
