杨梅素对肝癌SMMC-7721细胞迁移及侵袭影响的研究
2017-06-22沈维干许正新
朱 磊,沈维干,许正新
(扬州大学医学院,江苏 扬州 225001)
杨梅素对肝癌SMMC-7721细胞迁移及侵袭影响的研究
朱 磊,沈维干,许正新
(扬州大学医学院,江苏 扬州 225001)
目的 探讨杨梅素对肝癌SMMC-7721细胞迁移和侵袭的影响。方法 以不同浓度杨梅素处理的肝癌SMMC-7721细胞为研究对象,分别采用Wound healing实验、Transwell细胞迁移和侵袭实验检测细胞的迁移和侵袭过程,用RT-qPCR和Western blot检测E-cadherin和N-cadherin的表达。结果 杨梅素呈浓度依赖性(10~40 μmol·L-1)抑制肝癌SMMC-7721细胞活力;杨梅素还可以抑制SMMC-7721细胞迁移和侵袭过程,并且随着杨梅素浓度的增加,细胞中丝状、片状伪足逐渐减少,细胞排列越来越紧密;RT-qPCR和Western blot结果均显示,杨梅素可以上调SMMC-7721细胞的E-cadherin表达,下调N-cadherin表达。结论 杨梅素能抑制肝癌SMMC-7721细胞的迁移和侵袭过程,此作用的发生可能是通过上调E-cadherin、下调N-cadherin等EMT相关的信号通路以及肌动蛋白细胞骨架的重排实现的。
上皮间质样转化;杨梅素;SMMC-7721细胞;细胞迁移;细胞侵袭;细胞骨架
肝癌是世界范围内最常见的恶性肿瘤之一,在我国恶性肿瘤中死亡率位列第2位[1]。近年来随着诊疗水平的改善,肝癌患者的生存率虽有较大提高,但是术后肝癌细胞的转移已成为影响肝癌预后的重要因素[2]。而肝癌转移是多基因参与的、复杂的过程,它涉及大量基因异常表达以及相关信号通路的异常[3]。其中, 上皮间质样转化(epithelial-to-mesenchymal transition,EMT)被认为是肿瘤细胞获得迁移和侵袭能力的重要过程;在EMT过程中,肿瘤细胞会丧失本身的上皮样特性而具有某些间质细胞的特性,从而促使细胞的迁移和侵袭[4],而抑制EMT过程则可抑制癌细胞的生长和转移[5]。
杨梅素(myricetin)是一种天然存在的黄酮醇类化合物,广泛存在于双子叶植物中,化学名为3,3′,4′,5,5′,7-六羟基黄酮(Fig 1),又名杨梅树皮素[6],文献报道,该化合物具有抗炎、镇痛[7]、降血糖[8]和保肝[9]等药理作用。此外,Labbé、郑作文等还发现,杨梅素也具有抗肿瘤活性[10-11],然而,此方面的报道目前还不是很多,相关的研究还不够深入,这就给我们对本领域的深入研究提供了空间。

Fig 1 Chemical structural formula of myricetin
李蓉等[12]研究结果表明,漆树黄酮可以通过逆转肝癌HepG2细胞的EMT来抑制细胞的迁移和侵袭能力。庞慧芳等[13]也发现,姜黄素可以通过抑制胰腺癌PANC-1细胞的EMT过程来影响细胞的转移和侵袭。上述结果表明,某些药物可以通过调控EMT过程等环节来影响细胞的运动能力,那么杨梅素的抗肿瘤作用是否也有EMT等因素的参与呢?基于此考虑,本研究将以人肝癌SMMC-7721细胞为研究对象,观察杨梅素对细胞迁移和侵袭的影响并进一步探索杨梅素可能的作用机制,为今后杨梅素在疾病的预防治疗上的进一步开发利用提供实验基础和理论依据。
1 材料与方法
1.1 细胞株与实验试剂 人肝癌SMMC-7721细胞株(本实验室保存)用含10%胎牛血清高糖DMEM培养。杨梅素购于成都普思生物公司,纯度为98%,以DMSO配成500 g·L-1的贮存液/母液(-20 ℃冰箱中保存),临用前以DMEM培养基配成各种不同浓度的工作液;DMEM高糖培养基购于美国Gibco公司;胎牛血清购于上海洛神生物技术公司;RIPA细胞裂解液、胰蛋白酶、四氮唑蓝(MTT)和SDS-PAGE凝胶配置试剂盒均购于碧云天生物技术公司;小鼠抗人GAPDH 单抗、兔抗人E-cadherin单抗和兔抗人N-cadherin单抗以及辣根过氧化物酶(HRP)标记的山羊抗兔和山羊抗小鼠的二抗均购于美国Cell Signaling Technology公司。
1.2 细胞活力检测(MTT实验) 取SMMC-7721细胞制成单细胞悬液,计数后接种于96孔培养板中(1×104个细胞/孔);置于培养箱中培养过夜;向96孔板中加入不同浓度的杨梅素,共分为7组(分别为0、10、20、30、40、50、60 μmol·L-1),每组6复孔,培养48 h后;取出96孔板,每孔中加入5×10-3mg·L-1的MTT溶液10 μL,继续培养4 h后,吸弃上清,向每孔中加入DMSO 150 μL,在摇床上以100 r·min-1避光孵育15 min后,用酶标仪测定490 nm处的OD值并绘制细胞活力曲线图。实验重复3次。
1.3 细胞形态观察 取SMMC-7721细胞,将密度为3×104个/孔的细胞接种于24孔板中,待细胞贴壁后分别加入含杨梅素的完全培养基,置培养箱中培养48 h;在显微镜下观察细胞的形态。
1.4 细胞划痕实验 取SMMC-7721细胞接种于6孔板中(1×106个细胞/孔),待细胞汇合度约为0.70时,换无血清DMEM培养过夜,次日用枪头划出“伤痕”线,并在光学显微镜下观察、拍摄细胞开始时划痕状态(即0 h);随后向6孔板中分别加入含0、10、20、40 μmol·L-1杨梅素的无血清DMEM,继续培养24 h和48 h后分别在倒置相差显微镜下拍照。每个实验组取5个视野,实验重复3次。
1.5 Transwell细胞迁移和细胞侵袭实验 细胞迁移实验是将SMMC-7721细胞用无血清DMEM培养过夜后制备单细胞悬液;取Transwell小室置于24孔板中,先在Transwell下室中分别加入600 μL含0、10、20、40 μmol·L-1杨梅素的完全培养基;随后向Transwell 上室中分别加入200 μL含0、10、20、40 μmol·L-1杨梅素的无血清细胞悬液(细胞终浓度为2.5×108·L-1);置于培养箱中培养24 h;取出Transwell小室,用PBS漂洗后将小室浸泡在吉姆萨染液中染色,PBS冲洗后用棉签轻柔擦去上室内未迁移的细胞,在倒置相差显微镜下,观察膜底细胞数目、拍照、统计。每孔随机计数6个视野(100×)。每个实验组设2个复孔,实验重复3次。
细胞侵袭实验的实验操作与Transwell细胞迁移实验基本相同,仅稍做修改:① 将Matrigel胶放入冰箱中隔夜使其解冻,在膜上表面铺上100 μL Matrigel(用DMEM稀释成1 mg·L-1),置于培养箱中孵育8 h,使Transwell小室中形成一个基质屏障膜;② 将含有Transwell小室的孔板置培养箱中培养48 h。每个实验组设2个复孔,实验重复3次。
1.6 免疫荧光实验 将SMMC-7721细胞接种于铺有盖玻片的6孔板中,置培养箱中培养;待细胞贴壁后分别在孔中加入含0、10、20、40 μmol·L-1杨梅素的完全培养基;继续培养48 h后(确保细胞汇合度约为0.70),取出盖玻片,PBST漂洗后,用固定液室温固定细胞30 min,漂洗后加入体积分数为0.002 Triton X-100的PBS透化10 min,再漂洗;用罗丹明标记的鬼笔环肽室温避光孵育1h,漂洗后再用5 mg·L-1DAPI避光孵育15 min,充分漂洗后加抗荧光淬灭剂封片;于荧光显微镜下观察,随机拍照。
1.7 RT-qPCR实验 取对数生长期的SMMC-7721细胞种于6孔板中,待密度为0.70左右时加入相应浓度(0、10、20、40 μmol·L-1)的杨梅素作用48 h。用Trizol提取细胞总RNA;取2 μL RNA样品用于浓度和纯度的测定;对总RNA样品按照TaKaRa宝生物工程(大连)有限公司的PrimeScriptTMRT Master Mix试剂盒的说明书进行反转录合成cDNA。对合成的cDNA样品按照南京Vazyme公司的AceQTMqPCR SYBR Green Master Mix试剂盒的说明书进行Real time-PCR反应,每种浓度的cDNA样品设3个复孔,实验重复3次。实验所用引物见Tab 1。qPCR反应的条件是95 ℃,5 min预变性,95 ℃,10 s和60 ℃,34 s共40个循环。实验结果可采用2-△△Ct来分析。
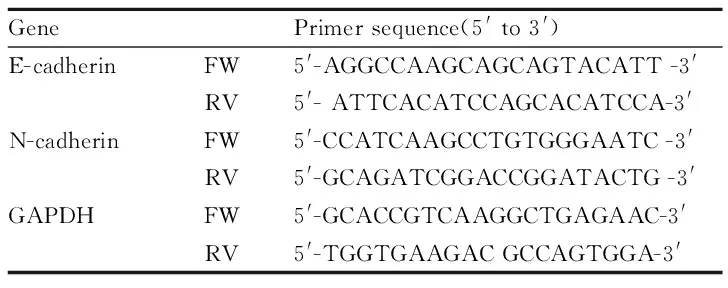
Tab 1 Primers used for real-time PCR
1.8 Western blot实验 将细胞种于6孔板中,待生长至50%左右密度分别加入0、10、20和40 μmol·L-1的杨梅素作用48 h;收集细胞,加入RIPA buffer裂解细胞,裂解后收集上清,加入上样缓冲液,在沸水中煮5~10 min可制得蛋白样品备用;取细胞裂解后收集的上清进行BCA蛋白定量,确定蛋白样品的浓度;根据定量结果将蛋白样品置于SDS-PAGE上进行电泳分离,并在PVDF膜上进行转膜反应;用质量浓度为50 g·L-1的封闭用奶粉进行封闭;加入稀释的E-cadherin、N-cadherin、Vimentin、GAPDH等单克隆抗体于4 ℃下孵育过夜;加入相应HRP标记的二抗;在凝胶成像系统中进行化学发光。实验结果以GAPDH作为内参,利用软件Image J对蛋白条带进行灰度分析。
2 结果
2.1 杨梅素对肝癌细胞活力的影响 为了证明杨梅素对SMMC-7721细胞活力是否具有影响,本研究采用MTT法检测不同浓度的杨梅素处理前后的细胞的活力,结果见Fig 2。由图可见,与对照组(0 μmol·L-1)相比,细胞活力在杨梅素浓度为0和40 μmol·L-1之间均没有明显改变,提示杨梅素浓度在这个范围内对细胞的影响较小;而当浓度增加至50~60 μmol·L-1时,细胞活力则明显下降,说明此浓度范围的杨梅素对细胞具有明显的毒性作用。因此本项研究选择浓度为10、20、40 μmol·L-1的杨梅素作下一步的继续探讨。

Fig 2 Effect of myricetin on viability of SMMC-7721 cells
*P<0.05vscontrol
2.2 杨梅素对SMMC-7721细胞形态的影响 为了说明杨梅素对SMMC-7721细胞的形态是否具有影响,本实验系统观察了不同浓度的杨梅素处理前、后(48 h)的细胞的形态变化,结果见Fig 3。结果显示:与对照组(0 μmol·L-1)比较,SMMC-7721细胞经40 μmol·L-1的杨梅素作用48h后,其细胞形态由长梭形间质样细胞转变为鹅卵石状上皮样细胞;提示杨梅素可能促进SMMC-7721细胞的形态发生EMT(上皮间质样转化)向MET(间质上皮样转化) 的转换。

Fig 3 Effect of myricetin on cell morphous of SMMC-7721 cells
2.3 杨梅素对SMMC-7721迁移能力的影响 为了研究杨梅素能否影响SMMC-7721细胞的迁移亦或其影响程度,本实验分别采用细胞划痕实验和Transwell细胞迁移实验对此进行研究。在细胞划痕实验中,先对用不同浓度杨梅素处理的SMMC-7721细胞“伤口”愈合情况进行评价。如Fig 4A所示,与对照组细胞相比,无论是24 h还是48 h,随着药物浓度的增加,“伤口”的面积越大,伤口愈合的速度越慢;并且在48 h时,杨梅素对SMMC-7721细胞的伤口愈合速度仍然有抑制作用。结果提示杨梅素能明显抑制SMMC-7721细胞“二维”平面的迁移,且这种抑制作用具有时间和浓度依赖性。
Transwell 细胞迁移实验结果见Fig 4B和4C。由图可知,与对照组相比,随着杨梅素浓度的增加,SMMC-7721细胞的迁移数量逐渐减少。提示杨梅素能浓度依赖性地抑制SMMC-7721细胞“三维”平面的迁移。因此,细胞划痕实验和Transwell细胞迁移实验结果均证明杨梅素确实对SMMC-7721细胞的迁移有明显抑制作用。
2.4 杨梅素对SMMC-7721细胞侵袭的影响 为了探讨杨梅素是否能对SMMC-7721细胞侵袭能力产生影响,本实验分别给予细胞进行0、10、20、40 μmol·L-1各不同浓度的杨梅素处理,采用Transwell细胞侵袭实验对细胞侵袭能力进行检测,结果见Fig 4B和4D。由图可见,与对照组相比,当杨梅素的浓度为20和40 μmol·L-1时可以明显地抑制SMMC-7721细胞侵袭(P<0.05)。
2.5 杨梅素对SMMC-7721细胞肌动蛋白细胞骨架纤维重排的影响 肌动蛋白细胞骨架纤维在装配动力学上的改变在细胞迁移和侵袭过程中发挥了重要作用。前述实验已经证明杨梅素能抑制SMMC-7721细胞的迁移和侵袭,那么,此作用是否有杨梅素影响了SMMC-7721细胞肌动蛋白细胞骨架纤维重塑因素的参与?为了回答这一问题,我们设计了这项实验,结果如Fig 5所示。由图可见,与对照组相比,随着杨梅素浓度的增加,SMMC-7721细胞中张力纤维数目明显减少,细胞边缘的丝状伪足和片状伪足也有所减少;实验结果提示,杨梅素对SMMC-7721细胞迁移和侵袭的影响可能是通过影响细胞中肌动蛋白细胞骨架纤维的重塑来实现的。
2.6 杨梅素对EMT相关标记基因表达的影响 为了进一步阐明杨梅素调控SMMC-7721细胞的迁移和侵袭的分子机制,本实验分别采用RT-qPCR和Western blot检测不同浓度的杨梅素处理后SMMC-7721细胞中的E-cadherin、N-cadherin等相关标记基因的表达情况。RT-qPCR实验结果如Fig 6A所示,与对照组相比, E-cadherin相对表达量在杨梅素浓度为40 μmol·L-1时明显上升(P<0.05);而N-cadherin的相对表达量在杨梅素浓度为20和40 μmol·L-1时明显下降(P<0.05)。Western blot实验结果如Fig 6B和6C所示,与对照组相比,E-cadherin蛋白相对表达量随着杨梅素浓度增加而明显上调(P<0.05);N-cadherin蛋白相对表达量随着杨梅素浓度增加而明显下调(P<0.05)。
以上结果证明,杨梅素可能通过上调E-cadherin,下调N-cadherin等相关标记基因的表达来抑制细胞的迁移和侵袭。
3 讨论
EMT是上皮细胞受外源或内源性因素刺激后,通过细胞骨架重塑转变为具有运动能力的间质细胞从而获得有较强的转移能力以及侵袭、降解细胞外基质能力等间质表型的生物学过程[3]。细胞骨架纤维重塑过程需要肌动蛋白(F-actin)参与,F-actin通过改变细胞片状伪足及丝状伪足的形成,进而影响细胞迁移和侵袭过程[14]。本实验采用罗丹明标记的鬼笔环肽染色SMMC-7721细胞中F-actin,结果发现随着杨梅素浓度的增加,SMMC-7721细胞中张力纤维数目逐渐减少,同时细胞边缘的丝状伪足及片状伪足也有所减少,在形态学上表现为促进SMMC-7721细胞的形态发生EMT向MET的转换。为了说明这种转变与细胞迁移和侵袭能力的改变是否存在必然的联系,本研究通过RT-qPCR以及Western blot检测了E-cadherin和N-cadherin的表达情况,后两者的实验结果进一步证明了细胞的EMT过程受到抑制以致EMT向MET的转换。由此证明,杨梅素具有抑制SMMC-7721细胞迁移及侵袭的作用。
有研究表明EMT与肿瘤细胞的浸润过程密切相关[6]。在EMT发生过程中,上皮标志物E-cadherin(E-钙粘蛋白)表达水平会下调,间质标志物N-cadherin(N-钙粘蛋白)表达水平会上调[4],最终导致肿瘤细胞的迁移和侵袭等运动能力增强,使肿瘤细胞更易于发生原位浸润或转移至体内的其他部位,在新的组织或器官中重新定位[7]。EMT除影响肿瘤进展的发生机制外,还与抗肿瘤药物的敏感性以及病人预后等相关。Jenchlinger等[15]发现,在MAPK 介导下发生EMT时,可出现MDR2(多药耐药基因2)的高表达。故研究EMT的发生机制对发现新的肿瘤治疗靶点至关重要。
目前陆续有一些报道显示杨梅素具有抗肿瘤作用,该药或者能够影响肺癌及前列腺癌细胞的增殖[16-17],或者通过抑制肝细胞恶化、降低肝癌发病率[10]。在此基础上,本文对杨梅素抗肿瘤作用的分子机制进行了初步探讨,研究结果对基础和临床上关于该药的后续深入开发和进一步挖掘提供了理论依据。但是由于SMMC-7721细胞是迁移能力较低的细胞,因此,为了更好的探究杨梅素抑制肝细胞转移亦或抗肿瘤作用,还需要其他的多种手段和措施的反复验证。

Fig 4 Effect of myricetin on migration and invasion of SMMC-7721 cells
A:Cell wound healing assay detected effect of myricetin on cell migration of SMMC-7721 cells;B~D:Transwell cell migration assay and cell invasion assay detected effect of myricetin on migration and invasion of SMMC-7721 cells;*P<0.05vscontrol

Fig 5 Effect of myricetin on actin cytoskeleton rearrangement of SMMC-7721 cells


综上所述,杨梅素可能是干扰细胞骨架的重塑进而促进细胞发生EMT向MET转换来抑制细胞的迁移和侵袭。本实验初步探讨杨梅素对肝癌细胞迁移和侵袭的抑制作用,为杨梅素进一步开发利用提供实验基础和理论依据,但具体作用机制仍需进一步研究。
(致谢:感谢我的师姐郑雪,师妹马宏昕、任静娜,师弟饶本龙对我的实验提供很多的帮助与支持。)
[1] 陈万青, 郑荣寿, 曾红梅, 等. 2011年中国恶性肿瘤发病和死亡分析[J]. 中国肿瘤, 2015, 24(1):1-10.
[1] Chen W Q,Zheng R S,Zeng H M,et al. Report of cancer incidence and mortality in China,2011[J].ChinaCancer, 2015, 24(1):1-10.
[2] El-Serag H B. Hepatocellular carcinoma[J].NEnglJMed, 2011, 365(12):1118-27.
[3] Chen L, Chan T H, Yuan Y F, et al. CHD1L promotes hepatocellular carcinoma progression and metastasis in mice and is associated with these processes in human patients[J].ClinInvest, 2010, 120(4):1178-91.
[4] Huber M A, Kraut N, Beug H. Molecular requirements for epithelial-mesenchymal transition during tumor progression[J].CurrOpinCellBiol, 2005, 17(5): 548-58.
[5] Bi X, PohI N M, Qian Z, et al. Decorin- mediated inhibition of colorectal cancer growth and migration is associated with E-cadherininvitroand in mice[J].Carcinogenesis, 2012, 33(2): 326-30.
[6] Lee K W, Kang N J, Rogozin E A, et al. Myricetin is a novel natural inhibitor of neoplastic cell transformation and MEK1[J].Carcinogenesis, 2007, 28(9): 1918-27.
[7] Park H H, Lee S, Son H Y, et al. Flavonoids inhibit histamine release and expression of proinflammatory cytokines in mast cells[J].ArchPharmRes, 2008,31(10):1303-11.
[8] Liu I M, Tzeng T F, Liou S S, et al. Improvement of insulin sensitivity in obese Zucker rats by myricetin extracted from Abelmoschus moschatus[J].PlantaMed,2007, 73(10):1054-60.
[9] Maheshwari D T, Yogendra Kumar M S, Verma S K, et al. Antioxidant and hepatoprotective activities of phenolic rich fraction of Seabuckthorn(Hippophae rhamnoides L.) leaves[J].FoodChemToxicol, 2011, 49(9):2422-8.
[10]Labbé D, Provencal M, Lamy S, et al. The flavonols quercetin, kaempferol, and myricetin inhibit hepatocyte growth factor-induced medulloblastoma cell migration[J].JNutr, 2009, 139(4):646-52.
[11]郑作文, 阎 莉. 藤茶双氢杨梅树皮素抗肝癌H22的实验研究[J]. 中华中医药学刊, 2006, 24(9):1627-9.
[11]Zheng Z W, Yan L. The Epermiental Study of Ampelops in on Antitumor Effect[J].ChinaArchTraditChinMed, 2006,24(9):1627-9.
[12]李 蓉, 黎小兵, 敬 敏, 等. 漆树黄酮逆转人肝癌细胞HepG2上皮间质转化的实验研究[J]. 中国药理学通报, 2011, 27(1):91-5.
[12]Li R, Li X B, Jing M, et al. Effect of fisetin on epithelial-mesenchymal transition in hepatocellular carcinoma cell line HepG2[J].ChinPharmacolBull, 2011, 27(1):91-5.
[13]庞慧芳, 覃 华, 赵 秋, 等. 姜黄素对胰腺癌细胞上皮间质转化的影响[J]. 世界华人消化杂志, 2014, 22(18):2565-71.
[13]Pang H F, Qin H, Zhao Q, et al. Curcumin inhibits TGF-β1-induced epithelial mesenchymal transition of pancreatic cancer cells[J].WorldChinJDigestol,2014, 22(18):2565-71.
[14]Wicki A, Lehembre F, Wick N, et al. Tumor invasion in the absence of epithelial-mesenchymal transition: Podoplanin-mediated remodeling of the actin cytoskeleton[J].CancerCell, 2006,9(4):261-72.
[15]Jechlinger M, Grunert S, Tamir I H, et al. Expression profiling of epithelial plasticity in tumor progression[J].Oncogene, 2003, 22(46):7155-69.
[16]Lu J, Papp L V, Fang J, et al. Inhibition of mammalian thioredoxin reductase by some flavonoids: implications for myricetin and quercetin anticancer activity[J].CancerRes, 2006, 66(8):4410-8.
[17]De Leo M, Braca A, Sanogo R, et al. Antiproliferative activity of Pteleopsis suberosa leaf extract and its flavonoid components in human prostate carcinoma cells[J].PlantaMed, 2006, 72(7):604-10.
Effect of myricetin on migation and invasion of hepatoma SMMC-7721 cells
ZHU Lei, SHENG Wei-gan, XU Zheng-xin
(SchoolofMedicine,YangzhouUniversity,YangzhouJiangsu225001,China)
Aim To investigate the effects of myricetin on the migration and invasion of hepatoma SMMC-7721 cells.Methods SMMC-7721 cells were treated with different concentrations of myricetin. Migration and invasion of SMMC-7721 cells were examined by Wound healing and Transwell, and the expression of E-cadherin and N-cadherin was detected by RT-qPCR and Western blot.Results Myricetin may inhibit the viability of SMMC-7721 cells via different concentration(10~40 μmol·L-1); furthermore, myricetin might also inhibit the migration and invasion of hepatoma SMMC-7721 cells. Meanwhile with the increasing of myricetin concentration, both filopodia and lamellipodia formation was reducd, and 7721 cells displayed in more integrity. And data from RT-qPCR and Western blot demonstrated that myricetin may up-regulate the expression of E-cadherin, simultaneously, down-regulate N-cadherin in SMMC-7721 cells.Conclusion Myricetin may influence the cell migration and invasion through up-regulating the expression of E-cadherin, simultaneously, down-regulating N-cadherin and activate cytoskeleton remolding of SMMC-7721.
EMT; myricetin; SMMC-7721 cell; cell migration; cell invasion; cytoskeleton
时间:2017-5-25 17:44 网络出版地址:http://kns.cnki.net/kcms/detail/34.1086.R.20170525.1744.032.html
2017-01-15,
2017-03-22
国家自然科学基金面上项目(No 31071171);遗传工程国家重点实验室开放课题(SKLGE-1405)
朱 磊(1985-),男,硕士生,研究方向:药理学,E-mail:395923269@qq.com; 许正新(1964-),男,博士,教授,硕士生导师,通讯作者,E-mail:xuzhengxin405@126.com
10.3969/j.issn.1001-1978.2017.06.016
A
1001-1978(2017)06-0823-07
R284.1;R329.24;R73-37;R735.702.2
