瑞丽山龙眼幼苗光合特性研究
2017-05-30马加芳李贵祥柴勇和丽萍孟广涛
马加芳 李贵祥 柴勇 和丽萍 孟广涛
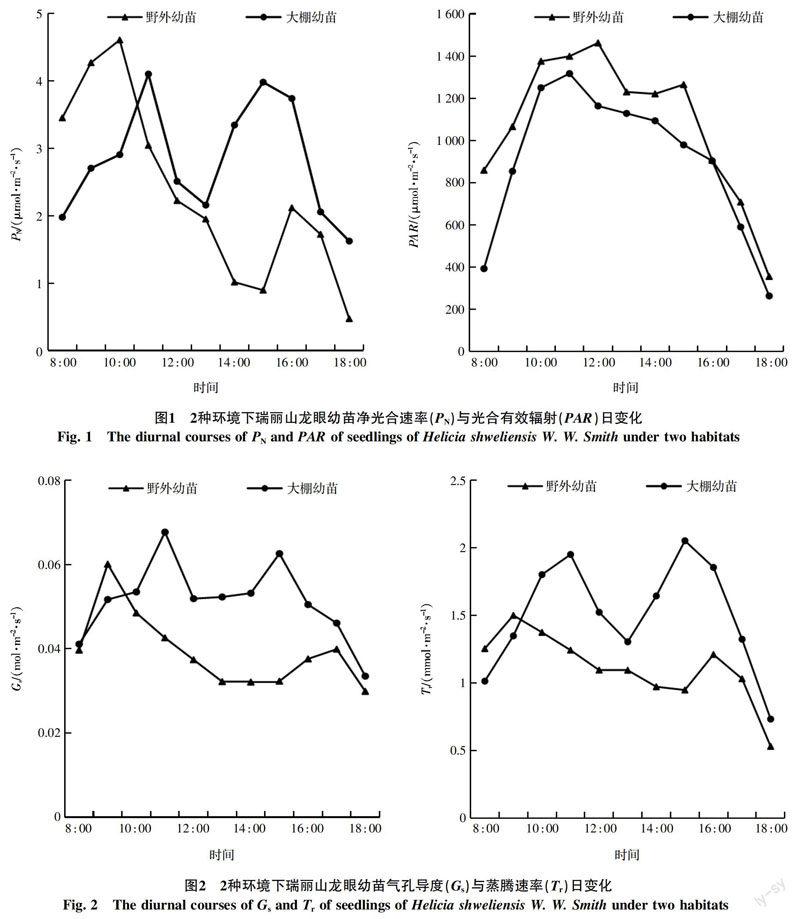
摘 要 使用Li-6400XT便携式光合仪测定高黎贡山南段2种环境(野外、大棚)下生长的我国特有種植物瑞丽山龙眼幼苗的光合特性,结果表明:①2种环境下生长的幼苗净光合速率(PN)日变化出现“双峰”曲线,有“午休”现象,但峰值出现的时刻与大小不同,野外幼苗在10 : 00、16 : 00出现高峰值,分别为4.608、2.118 μmol/(m2·s),大棚幼苗在11 : 00、15 : 00出现2个高峰值,分别为4.102、3.982 μmol/(m2·s);蒸腾速率(Tr)、气孔导度(Gs)变化趋势与净光合速率变化趋势一致,也有“午休”现象;水分利用率(WUE)无明显变化规律;胞间CO2浓度(Ci)与净光合速率日变化相反。②净光合速率与环境因子偏相关性分析表明,环境因子:光合有效辐射(PAR)、大气CO2浓度(Ca)、空气温度(Ta)、相对湿度(RH)对幼苗PN值影响均为:PAR>Ca>Ta>RH;③两地幼苗光饱和点(LSP)均较高,野外为2 674.995 μmol/(m2·s)、大棚幼苗为2 228.754 μmol/(m2·s),光补偿点(LCP)分别为38.250、8.322 μmol/(m2·s)。
关键词 瑞丽山龙眼;幼苗;光合特性;不同环境
中图分类号 Q945.11;S667.2 文献标识码 A
Abstract The photosynthetic characteristics of Helicia shweliensis W. W. Smith seedlings of two habitats(wild, nursery)in the southern section of Gaoligong Mountains were measured by Li-6400XT Portable Photosynthesis System. Research indicated that: ①The diurnal courses of net photosynthetic rate(PN)of the two seedlings occurred“double peak”curve and“lunch break”phenomenon, but the peak time was different of the two seedlings, that for the wild appeared at 10 : 00 and 16 : 00 was 4.608 μmol/(m2·s)and 2.118 μmol/(m2·s), and that for the nursery appeared at 11 : 00 and 15 : 00 was 4.102 μmol/(m2·s)and 3.982 μmol/(m2·s). Transpiration rate(Tr), stomatal conductance(Gs)trends were consistent with the net photosynthetic rate trends. Water use efficiency(WUE)had no significant variation. Intercellular CO2 concentration(Ci)was opposite to the net photosynthetic rate. ②The net photosynthetic rate and environmental factors correlation analysis showed that: PN was influenced by environmental factors in the order from high to low: light intensity(PAR), the atmospheric CO2 concentration(Ca), air temperature(Ta), relative humidity(RH). ③The light saturation point(LSP)of the two seedlings were high. That for the wild was 2 674.995 μmol/(m2·s), and that for the nursery was 2 228.754 μmol/(m2·s). The light compensation point(LCP)for the two was 38.250 μmol/(m2·s), 8.322 μmol/(m2·s), respectively.
Key words Helicia shweliensis W. W. Smith; seedlings; photosynthetic characteristics; different habitats
doi 10.3969/j.issn.1000-2561.2017.04.009
瑞丽山龙眼(Helicia shweliensis W. W. Smith)属于山龙眼科(Proteaceae),山龙眼属(Helicia),多为乔木或灌木,是中国特有种,仅分布于云南西部海拔2 000 m左右的较小范围内,在研究中国本土植物地理分布中具有一定意义。近年来由于现有植被破坏严重,加之数量较少,导致其陷入了濒危境地。从国内研究来看,瑞丽山龙眼因为分布较为特殊,目前还未见该植物任何方面的相关研究报道。
光合作用的研究在理论上和实践上都有很大意义[1],根据对一部分濒危植物的更新能力与濒危机制的研究,生长环境的特殊性,光适应范围的宽窄,光合作用的强弱等对于植物的存活有一定影响[2-4]。国内研究重点植物的光合特性报道较多[5-8],多从植物的光合日变化、光响应机制、CO2响应机制等方面进行研究,同时植物的光合作用会受植物自身因素和外界环境的影响,故对于不同环境条件下的植物光合作用的研究也比较常见[9-11]。对于瑞丽山龙眼的光合特性报道还未见,故本研究在瑞丽山龙眼生长旺盛期对其光合作用进行了初步测定,并分析了影响光合作用的主要环境因子,旨在为其濒危机制的研究提供方向及科学保护提供一定指导。
1 材料与方法
1.1 材料
试验于2016年5月中旬的晴朗天氣在云南省高黎贡山南段(东经98°45′,北纬24°50′)的大棚和野外荒草地进行,两地均属于亚热带高原季风气候,海拔2 000~2 400 m,年平均气温约15℃,年平均降水量约1 650 mm,分配不均匀,多集中在4~9月,干湿季分明[12]。其中,试验野外地以肾蕨(Nephrolepis auriculata L. Trimen)为优势种,伴生有芒萁(Dicranopteris dichotoma)、黄毛草莓(Fragaria nilgerrensis Schltdl)等,无灌木与乔木,处于迎风坡,一天内温差较大;试验大棚平均温度20 ℃,湿度70%以上,遮荫网密度15%,温差幅度较小。
测量仪器选用美国生产的Li-6400便携式光合作用仪。
大棚内幼苗为三年生实生苗,苗圃常规管理,定时浇水通风;在野外选择与大棚内生长大小一致的实生幼苗材料进行试验。选两地幼苗各3株,平均植高35 cm,要求所选叶片大小一致,叶片完整,分别对每一植株的第3至5叶位进行标记。
1.2 方法
1.2.1 光合作用日变化测定 试验时间为8 : 00~18 : 00,分别对标记的植株叶片依次进行测量,1次/h,每片叶子仪器标记3次。分别得到光合参数净光合速率(PN)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、光合有效辐射(PAR)、水分利用率(WUE=PN/Tr)。
以上步骤在典型的晴天中重复测量3 d。
1.2.2 光响应曲线的测定 光响应曲线测定试验时间为9 : 00~11 : 00。进行光响应测定时,控制叶温25 ℃,CO2浓度400 μmol/(m2·s),空气相对湿度50%,流速500 μmol/s,光合有效辐射梯度从高至低为2 000、1 500、1 000、500、250、120、60、30、15、0 μmol/(m2·s)。光响应曲线拟合模型表达式为PN=α·(1-β·PAR)·PAR/(1+γ·PAR)-Rd[13-14];幼苗光饱和点表达式为LSP={[(β+α)/β]1/2-1}/γ[15];幼苗光补偿点LCP,即当净光合速率为0时,所对应的光合有效辐射[16-17](α为曲线初始斜率,β为修正系数,γ为α与最大净光合速率比值,Rd为暗呼吸速率)。
以上步骤重复测量3 d。
1.3 数据处理
数据采用Microsoft Excel 2010绘图,SPSS Statistics 19.0进行相关分析处理。
2 结果与分析
2.1 净光合速率(PN)与光合有效辐射(PAR)日变化
由图1可以看出,2种不同环境下生长的瑞丽山龙眼幼苗净光合速率都在中午时段出现了明显的“午休”现象,日变化均呈现出“双峰”曲线,野外幼苗在10 : 00与16 : 00出现高峰值,分别为4.608、2.118 μmol/(m2·s),两峰值出现的时间相差长,即“午休”时间长;大棚幼苗在11 : 00与15 : 00出现2个高峰值,分别为4.102、3.982 μmol/(m2·s),两值出现的时间跨度小,即“午休”时间短;两地幼苗在18 : 00的净光合速率最小,前者为0.473 μmol/(m2·s),后者为1.624 μmol/(m2·s)。野外环境的光合有效辐射与大棚环境光合有效辐射相比较高,二者的日变化均呈现出“单峰”曲线,在8 : 00~11 : 00,光合有效辐射值快速上升,11 : 00~15 : 00变化幅度较小,16 : 00之后,光合有效辐射值逐渐下降;其中,野外光合有效辐射值在12 : 00达到最大值,为1 642.370 μmol/(m2·s),大棚光合有效辐射值在11 : 00达到最大值,为1 317.160 μmol/(m2·s),二者的最低光合有效辐射值都在18 : 00后,均低至300 μmol/(m2·s)左右。
2.2 气孔导度(Gs)与蒸腾速率(Tr)日变化
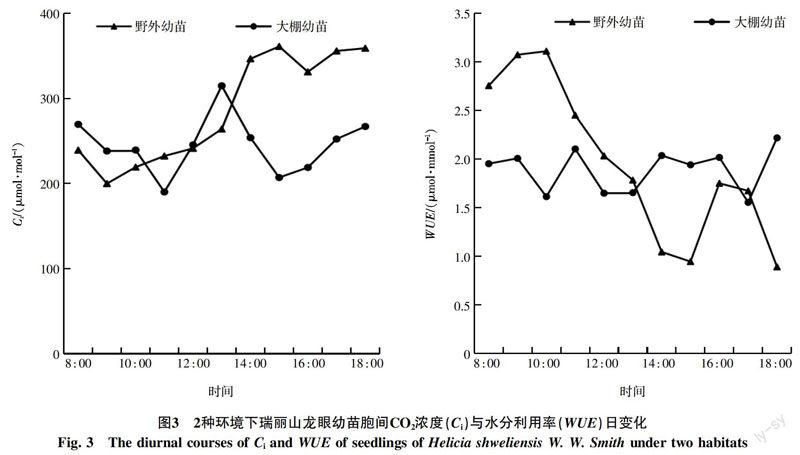
气孔导度与蒸腾速率也是植物光合特性的重要参数,一般情况下,二者的日变化趋势与净光合速率的日变化趋势相似。图2中,两环境下幼苗的变化趋势都不同程度上呈现“双峰”型,其中大棚幼苗的双峰趋势较为明显。气孔导度日变化中,野外幼苗在9 : 00出现一天中的最高值,为0.060 mol/(m2·s),17 : 00出现下午时段的小高峰值,为0.039 mol/(m2·s);大棚幼苗的2个高值出现在11 : 00和15 : 00,分别为0.068、0.063 mol/(m2·s)。二者的最低值都出现在18 : 00,分别为0.029、0.033 mol/(m2·s)。
蒸腾速率日变化中,野外幼苗的蒸腾速率日变化较为平稳,9 : 00出现一天中的最高值,为1.499 mol/(m2·s);但18 : 00出现极速下降,是一天中的最低值,为0.530 mmol/(m2·s)。大棚幼苗的2个高峰值为1.949、2.051 mmol/(m2·s),分别出现在11 : 00和15 : 00左右,其中15 : 00是一天中的最高值,最低值也是出现在18 : 00,为0.732 mmol/(m2·s)。
2.3 胞间CO2浓度(Ci)与水分利用率(WUE)日变化
胞间CO2浓度的日变化趋势与净光合速率变化趋势明显相反,这与濒危植物长序榆幼苗[18]、孑遗植物四合木[19]等植物相似,但从图3中可以看出,相比于大棚幼苗,野外幼苗的变化趋势较为平稳,一天中二者的最低值都出现在9 : 00~12 : 00,野外幼苗为200.006 μmol/mol,大棚幼苗为190.205 μmol/mol。
水分利用率的强弱与植物抗旱能力相关,2种环境下幼苗的水分利用率均未呈现出任何变化规律。相比而言,野外幼苗波动明显,在10 : 00达到一天中的最大值,为3.110 μmol/mmol,之后一直下降,在16 : 00~17 : 00又出现一个小高峰值,为1.752 μmol/mmol,在18 : 00值为0.892 μmol/mmol,是一天中的最低值;大棚幼苗的水分利用率值较平稳,都在1.5~2.5 μmol/mmol间波动。
2.4 光响应曲线
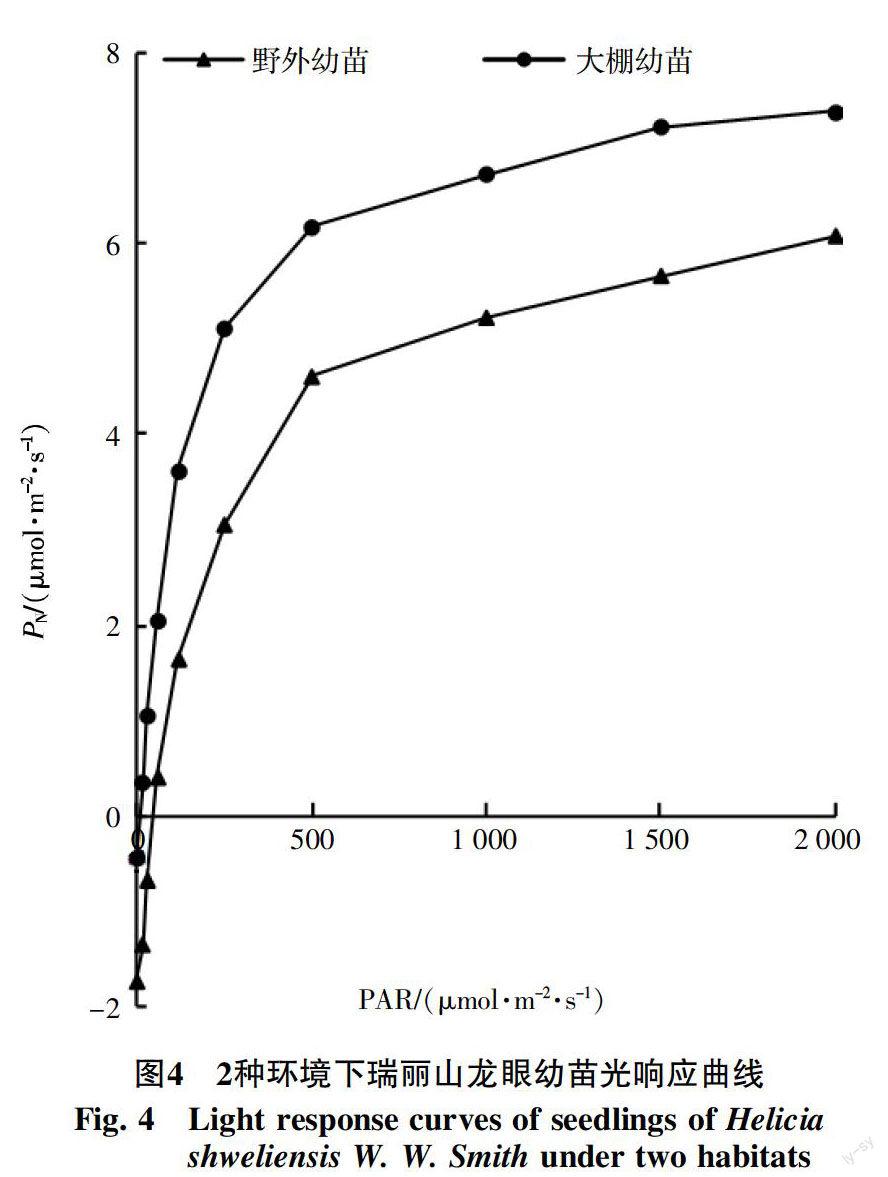
从图4中可知,2种不同环境下生长的幼苗,在光合有效辐射低于250 μmol/(m2·s)时,幼苗的净光合速率呈直线上升;在500~2000 μmol/(m2·s)时,上升趋势逐渐缓慢,但仍在上升,都未出现光抑制现象。且在相同的光合有效辐射下,大棚幼苗的净光合速率值高于野外幼苗。
通过光响应模型计算得知,表1中大棚幼苗的光饱和点(LSP)高于野外幼苗,对于光补偿点(LCP),大棚幼苗低于野外幼苗。
2.5 净光合速率与環境因子相关性分析
光合作用是一个复杂的过程,净光合速率的强弱不但与植物自身因素有关,还会受到植物生长环境中各因子的影响。通过合并计算野外生长和大棚生长的瑞丽山龙眼幼苗净光合速率(PN)与光合有效辐射(PAR)、大气CO2浓度(Ca)、空气温度(Ta)、相对湿度(RH)的偏相关性表明,各环境因子瑞丽山龙眼幼苗的影响大小为PAR(r=0.689,p=0.059)>Ca(r=0.516,p=0.191)>Ta(r=-0.478,p=0.231)>RH(r=0.221,p=0.598)。表2中,同时分析了几个环境因子之间也存在一定相关性,其中光合有效辐射与空气温度相关性最大。
3 讨论
在夏季晴朗的天气下,通过测定2种环境中生长的瑞丽山龙眼幼苗的各光合参数日变化,发现两地幼苗的净光合速率均为“双峰”曲线,中午会出现“午休”现象,同时两地幼苗的气孔导度与蒸腾速率在一天内的变化趋势与净光合速率的变化趋势存在一致性,9 : 00~11 : 00出现第一次高峰,15 : 00~17 : 00出现第二次高峰,而胞间CO2浓度的日变化则与净光合速率相反,水分利用率则无明显变化趋势。首先,根据And等[20]学者观点,可以认为,造成净光合速率、气孔导度、蒸腾速率变化趋势一致的原因是叶片气孔的闭合。夏季中午时段水分供应紧张,空气温度低,光合有效辐射增强,引起叶片气孔关闭。气孔导度减小,净光合速率也减小。而气孔导度会影响植物对水分的利用,气孔导度大,蒸腾速率增大[21],反之则小,所以三者变化呈现一致。其次,净光合速率与蒸腾速率的改变会制约水分利用率,莫凌等[22]通过研究濒危植物狭叶坡垒就曾表明,水分利用率低是造成该植物生长缓慢的一大原因,但在本研究中,水分利用率无明显变化趋势,具体原因以及是否对瑞丽山龙眼幼苗的生长具有影响需要进一步探究。最后,净光合速率与胞间CO2浓度的变化趋势相反,原因可能是外界CO2进入叶片细胞会存在一定阻力,净光合速率增大时,CO2供应量也增加,但由于存在进入阻力,胞内CO2得不到快速补充,浓度也就不高,反之,净光合速率减小,胞间CO2出现盈余,浓度就高[18]。但总体来说,净光合速率、气孔导度、蒸腾速率、胞间CO2浓度和水分利用率的趋势变化并不是完全由单因子引起,而是几者之间的相互作用,以及外界环境的影响。
探究到外界环境因素对净光合速率的影响,通过相关性分析,外界光合有效辐射,大气CO2浓度,空气温度,相对湿度对两地的幼苗都存在一定的影响。分析可知,光合有效辐射的影响一定存在,整个光合作用的系统动力都源于此,研究表明:濒危植物长叶榧[2]、金花茶[3]、狭叶坡垒[22]等的濒危机制就包括了光合有效辐射不足;CO2的影响同样重要,CO2是净光合速率产生不可或缺的原料;而温度主要通过影响参与光合作用的酶的活性起作用。
在光响应试验中,两地幼苗的光饱和点都在2 000 μmol/(m2·s)以上,说明瑞丽山龙眼属于阳生植物,人工育苗时可增加其的光照。二者的光补偿点野外幼苗高,为45.982 μmol/(m2·s),大棚幼苗为8.032 μmol/(m2·s)。总体来说,幼苗的光补偿点与光饱和点差值大,特别是大棚幼苗,说明瑞丽山龙眼能适应更大的光照幅度,这也与调查发现瑞丽山龙眼能分布在郁闭度较高的密林和较低的疏林中有关。
从整个试验看,两地幼苗生长阶段一致,但大棚幼苗由于生长小环境优越,长势更好,光合特性参数变化趋势也比野外幼苗好;所以,在人工种植时应更加注重瑞丽山龙眼幼苗从大棚移植户外时幼苗对自然环境的适应性,做好炼苗过程。
参考文献
[1] 潘瑞炽. 植物生理学(第7版)[M]. 北京: 高等教育出版社,2012: 70.
[2] 王 强, 金泽新, 郭水良, 等. 濒危植物长叶榧的光合生理生态特性[J]. 生态学报, 2014, 34(22): 6 460-6 470.
[3] 韦 霄, 郭 辰, 李吉涛, 等. 金花茶的濒危机制及保育对策[J]. 广西科学院学报, 2016, 32(1): 1-5.
[4] 刘 鹏, 康华靖, 廖承川, 等. 浙江九龙山自然保护区长序榆群落结构特征研究[J]. 浙江师范大学学报(自然科学版), 2007, 30(4): 435-439.
[5] Cui N B, Du T S, Kang S Z, et al. Relationship between stable carbon isotope discrimination and water use efficiency under regulated deficit irrigation of pear-jujube tree[J]. Agricultural Water Management, 2009, 96(11): 1 615-1 622.
[6] Zhang H N, Su P X, Li S J. Indicative effect of the anatomical structure of plant photosynthetic organ on WUE in desert region[J]. Acta Ecologica Sinica, 2013, 33(16): 4 909-4 918.
[7] Barber J. Photosynthetic energy conversion: natural and artificial[J]. Chemical Society Reviews, 2009, 38(1): 185-189.
[8] 李林鋒, 吴小凤, 刘素青. 湛江5种红树林树种光合作用特性及光合固碳能力研究[J]. 广西植物, 2015(6): 825-832.
[9] 林开文, 杨自云, 姜永雷, 等. 小叶女贞秋季光合速率日变化及其与环境因子的相关性[J]. 江苏农业科学, 2016, 44(1): 213-215.
[10] 高 辉, 黎云祥.不同遮荫条件下峨眉岩白菜光合特性初探[J].西南农业学报, 2015, 28(5): 1 992-1 997.
[11] 廖菊阳, 彭春良, 刘 艳, 等. 溪畔杜鹃光合特性及其与环境因子关系研究[J]. 中国农学通报, 2015, 31(13): 135-138.
[12] 袁春明, 孟广涛, 方向京, 等. 珍稀濒危植物长蕊木兰种群的年龄结构与空间分布[J]. 生态学报, 2012, 32(12): 3 866-3 872.
[13] 叶子飘. 光响应模型在超级杂交稻组合-Ⅱ优明86中的应用[J]. 生态学杂志, 2007, 28(8): 1 323-1 326.
[14] 宋佩东. 5种光响应模型对小叶杨的适用性分析[J]. 辽宁林业科技, 2015(3): 9-12.
[15] 钟 楚, 朱 勇. 几种光合作用光响应模型对烟草的适应性分析[J]. 中国农业气象, 2013, 34(1): 74-80.
[16] 叶子飘, 康华靖. 植物光响应修正模型中系数的生物学意义研究[J]. 扬州大学学报(农业与生命科学版), 2012, 33(2): 51-57.
[17] 刘宇锋, 萧浪涛, 童建华, 等. 非直线双曲线模型在光合响应曲线数据分析中的应用[J]. 中国农学通报, 2005, 21(8): 76-79.
[18] 高建国, 徐根娣, 李文巧, 等. 濒危植物长序榆(Ulmus elongata)幼苗光合特性的初步研究[J]. 生态环境学报, 2011, 20(1): 66-71.
[19] 智颖飙, 李红丽, 崔 艳, 等. 孑遗植物四合木(Tetraena mongolica)迁地保护中的光合作用日变化特征与生理生态适应性[J]. 生态环境学报, 2015, 24(1): 14-21.
[20] And G D F, Sharkey T D. Stomatal Conductance and Photosynthesis[J]. Annual Reviews of Plant Physiology, 2003, 33(33): 317-345.
[21] 郭二辉, 胡 聃, 田朝阳, 等. 城市环境下紫丁香与火炬树的光合生理特征研究[J]. 安徽农业大学学报, 2009, 36(4): 639-644.
[22] 莫 凌, 唐文秀, 毛世忠, 等. 珍稀濒危植物狭叶坡垒的光合特性[J]. 福建林学院学报, 2009, 29(4): 357-361.
