咖啡黑(枝)小蠹成虫触角感受器的扫描电镜观察
2017-05-30孙世伟刘爱勤王政桑利伟苟亚峰孟倩倩高圣风
孙世伟 刘爱勤 王政 桑利伟 苟亚峰 孟倩倩 高圣风


摘 要 对咖啡黑(枝)小蠹(Xylosandrus compactus)雌、雄成虫触角感受器进行了扫描电镜观察。结果表明:咖啡黑(枝)小蠹雌、雄成虫之间触角形态、感受器类型和分布均无显著差异。成虫触角具有5种感受器:毛形感受器(Ⅰ、Ⅱ)、锯齿形感受器、锥形感受器、芽形感受器和刺形感受器,其中毛形感受器数量最多,占总感受器的57%,刺形感受器数量最少,仅3%。各种感受器在触角各节上的分布数量显著不同,柄节、梗节感受器数量较少,分别约占感受器总数的4%和3%;鞭节分布的感受器数量最多,约占93%,其中约85%的感受器集中分布在锤状部末端。
关键词 咖啡黑(枝)小蠹;触角;感受器
中图分类号 S571.2 文献标识码 A
Abstract The antennae structures of female and male Xylosandrus compactus were observed with a scanning electron microscope. The results showed that there were no significant differences in the antennae morphological characters, sensilla types and their distribution patterns between sexes.The sensilla was identified for five types: sensilla trichodea(Ⅰ, Ⅱ), zigzag sensilla, sensilla basiconica, bud-shaped sensilla and sensilla chaetica. Sensilla trichodea was the dominant type, which accounted for more than 57%, while the number of sensilla chaetica was the least, only 3%. Most of the sensilla(about 93%)was distributed on the funicle of the antenna, of which about 85% of the sensilla was mainly distributed on the club.There were less sensilla on scapus and pedicel, only about 4% and 3%, respectively.
Key words Xylosandrus compactus; antenna; sensilla
doi 10.3969/j.issn.1000-2561.2017.04.017
咖啡黑(枝)小蠹(Xylosandrus compactus),異名Xyleborus compactus,Xyleborus morstatti Haged,又名黑色枝小蠹、楝枝小蠹,属鞘翅目(Coleoptera)小蠹科(Scolytidae)Xylosandrus属,是为害咖啡生产的重要害虫[1-3]。咖啡结果枝条被害后因水分及营养供应不足,导致干枝瘪果。中国国内于20世纪70年代在海南万宁首次发现其为害,随着咖啡种植面积的扩大,咖啡黑(枝)小蠹的为害日趋严重;发生严重时枝条受害率为29.7%~42.6%,最高达82.5%,植株受害率100%。该虫主要为害中粒种咖啡,小粒种咖啡受害较轻[4-5]。
小蠹科昆虫的化学信息物质,包括具有引诱活性的寄主气味、昆虫的聚集信息素、性信息素、利己素、利它素以及一些非寄主气味等。小蠹虫在对其寄主的搜寻、定向、攻击为害过程中,化学信息物质起着关键的作用[6-9]。研究表明,小蠹虫寄主选择行为是一个复杂过程,在寻找寄主阶段,多数小蠹虫能直接被寄主气味引诱,通过嗅觉感受器官在复杂生态系统内识别寄主树木挥发性化合物,从而达到对寄主树木的准确定位[10-12]。因此,化学感受器在小蠹虫寄主选择过程中发挥着十分重要的作用。毋亚梅等[13]、王玉刚等[14]、陈辉等[15]、杨群芳等[16]、范丽华等[17]、李宗波等[18]分别利用扫描电镜对松纵坑切梢小蠹(Tomicus piniperda L.)、红脂大小蠹(Dendroctonus valens)、华山松大小蠹(Dendroctonus armnadi)、光滑足距小蠹[Xylosandrus germanus(Blandford)]、脐腹小蠹(Scolytus schevyrewi)、小粒材小蠹(Xyleborus saxeseni)触角感受器进行了观察,表明小蠹虫触角上分布着大量的化学感受器,在寄主选择过程中发挥着十分重要的作用。触角是昆虫最主要的感受器官,研究触角感受器的形态、结构和分布是探索昆虫嗅觉行为和寄主识别机制的基础。
目前对咖啡黑(枝)小蠹的研究主要集中在其生物学习性、田间发生规律及化学防治技术等方面[2-5],尚未见到利用化学信息物质开展该虫寄主选择机制、信息素合成应用等相关报道。本文采用扫描电镜技术对咖啡黑(枝)小蠹成虫触角感受器类型和分布特点进行研究,旨在揭示咖啡黑(枝)小蠹成虫触角感受器的超微结构,为进一步探索其寄主选择行为机制提供理论依据。
1 材料与方法
1.1 材料
1.1.1 供试昆虫 咖啡黑(枝)小蠹成虫采集于海南省万宁市中国热带农业科学院香料饮料研究所咖啡试验基地,剪取受害咖啡枝条带回实验室饲养,饲养条件:环境温度T=(25±1)℃,相对湿度RH=(70±5)%。在体视镜下剖析枝条以获取成虫。
1.1.2 实验设备 体视镜(OLYMPUS SZX16),超声波清洗器(ELMA E120H),扫描电镜(卡尔蔡司ZEISS ΣIGMA场发射扫描电镜)。
1.2 方法
1.2.1 样品的制备与观察 取咖啡黑(枝)小蠹雌、雄成虫各30头,放入垫有湿滤纸的培养皿中供其自由爬行48 h,清水洗涤、晾干后放入4%戊二醛中,置于4 ℃条件下固定24 h,0.2 mol/L磷酸缓冲液清洗3次,每次1 min,然后用乙醇逐级脱水(30%、50%、70%、80%、90%和无水乙醇),乙酸异戊酯处理2次,每次20 min,CO2临界点干燥处理4 h,然后于体视镜下进行粘台,镀金,将经过上述步骤处理后的样品进行扫描电镜观察。
1.2.2 触角感受器的鉴定 感受器的类型主要参考Schneider[19]的方法进行命名和归类。
1.3 数据分析
触角感受器的分布特点及数量特征在扫描电镜照片中观察、统计,感受器百分比=(某种感受器数量/感受器的总数量)×100%。
2 结果与分析
2.1 触角的形态特征
咖啡黑(枝)小蠹雌、雄成虫触角(图1、 2)结构相似,均呈膝状,由柄节(scapus)、梗节(pedicel)和鞭节(funicle)3部分组成。其中,柄节棒锤状,梗节珠状,均为1节;鞭节分5个亚节,前4亚节呈套筒状,逐渐变粗,最后一亞节膨大成锤状部(club)。锤状部侧面扁平,正面近圆形。触角各节表面上均有明显的瓦棱状花纹。雌、雄成虫触角平均长度分别为(283.76±19.52)μm、(208.21±11.40)μm。
2.2 感受器的类型
通过扫描电镜观察,在咖啡黑(枝)小蠹成虫触角上共发现5类感器,分别为毛形感受器、锯齿形感受器、锥形感受器、芽形感受器和刺形感受器。其中,毛形感受器分为毛形感受器Ⅰ、毛形感受器Ⅱ2个亚型。此外,在锤状部末端还观察到一种表皮孔。
2.2.1 毛形感受器Ⅰ(sensilla trichodeaⅠ) 直立或沿触角轴微弯曲,细长,从基部到端部逐渐变细,末端钝圆,(图3,STⅠ),长度为(14.20±1.23)μm。数量最多,约占全部感受器的51%,集中分布于鞭节,尤以锤状部末端最为明显,与锯齿形、芽形等感受器混生。
2.2.2 毛形感受器Ⅱ(sensilla trichodeaⅡ) 直立或微弯曲,毛发状,整体细长而端部较尖,长度为(27.77±6.00)μm。(图4,ST Ⅱ)。主要分布于锤状部,尤以锤头部的近端处最为明显,占总感受器数量的6%。
2.2.3 锯齿形感受器(zigzag sensilla) 直立或与触角轴呈10°~60°角,形似毛发,每个感受器上具有5~12个不等锯齿,长度为(39.87±6.65)μm(图3,图5,ZS)。约占全部感受器数量的33%,在柄节、梗节和鞭节均有分布,以锤状部数量最多,柄节和梗节有少量分布。
2.2.4 锥形感受器(sensilla basiconica) 直立,形似锥子,生于柄节或锤状部,(图6,SB),长度为(6.91±1.25)μm。数量较少,约占全部感受器的4%。
2.2.5 芽形感受器(bud-shaped sensilla) 直立或微弯曲,形似禾谷种子的新芽,基部较粗大,顶端尖,着生于触角表面的凹窝中,长度为(4.08±0.70)μm(图7,BSS)。数量较少,仅分布于锤状部的末端,常与锥形感受器混生。
2.2.6 刺形感受器(sensilla chaetica) 外形刚直如刺,直立于触角表面,基部膨大,末端急尖。感受器长度雌虫为(6.93±3.05)μm(图8,SC)。数量少,分布于柄节和头部连接处的凹陷处及锤状部近端处。
2.2.7 表皮孔(cuticular pore) 分布于锤状部末端不着生感受器的空白区域内的一些稀疏小孔,直径小于1 μm。(图4,图7~8,CP)。多单生或并生于毛形、芽形等感受器附近。
2.3 感受器的分布规律
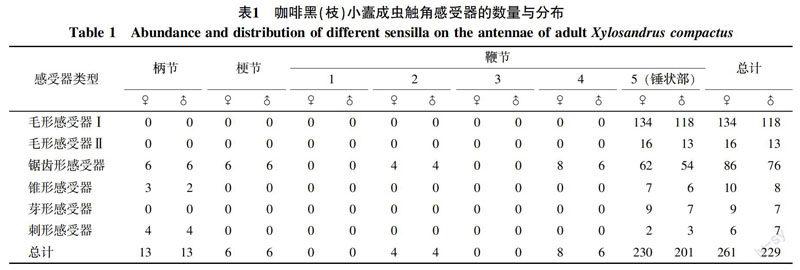
咖啡黑(枝)小蠹雌、雄成虫触角感受器的数量和分布特点见表1。由表1可知,咖啡黑(枝)小蠹雌、雄成虫触角感受器类型、数量和分布特点无明显差异。雌、雄成虫触角感受器类型相同,均有毛形、锯齿形、锥形、芽形和刺形5种类型,感受器的平均数量分别为261和229个,以毛形感受器最多,占感受器总数的57%以上,其次为锯齿形感受器,占总数的33%左右,这2种感受器的数量占绝对优势。其余锥形感受器、芽形感受器、刺形感受器数量合计占总数的10%左右。各类感受器在触角各节上的分布数量存在显著变化,柄节、梗节感受器数量较少,分别约占感受器总数的5% 和2%;鞭节分布的感受器数量最多,约占93%,其中约85%的感受器集中分布在锤状部末端。
各类感受器在成虫触角柄节、梗节和鞭节上的分布具有不同特点。柄节和梗节感受器数量少且类型相对单一,梗节仅有锯齿形1种感受器;鞭节则分布有全部5种感受器,但第1、3亚节没有感受器分布。此外,5种感受器在各节上的分布明显不同,锯齿形感受器在柄节、梗节、鞭节均有分布,锥形、刺形感受器分布在柄节和锤状部,而毛形和芽形感受器集中分布于锤状部。
3 讨论
咖啡黑(枝)小蠹雌雄成虫触角均由柄节、梗节和鞭节3部分组成,其中鞭节后半部分特化为锤状部;锤状部是感受器集中分布的区域。触角的整体形态、触角各节上感受器分布等特征与其他小蠹科昆虫相似[20-21]。咖啡黑(枝)小蠹成虫触角感受器在锤状部的排列方式与华山松大小蠹、红脂大小蠹等小蠹科不同属有所不同[14-17],咖啡黑(枝)小蠹感受器密集散生在锤状部的末2段,而后两者的毛形感受器在锤状部环绕触角排成界限分明的3 条带。
在咖啡黑(枝)小蠹成虫触角上共发现5类感受器,分别为毛形感受器、锯齿形感受器、锥形感受器、芽形感受器和刺形感受器。触角感受器类型与其他小蠹科昆虫存在属内相似性和种间差异性[13-18,22],同属的光滑足距小蠹分布有与其相同的5类感受器,且各类感受器的数量与分布特点存在较高的相似性;不同属的小粒材小蠹、红脂大小蠹、华山松大小蠹除分布有毛形、锯齿形、芽形和刺形4种感受器外,华山松大小蠹还分布有栓锥形感受器,小粒材小蠹有板形、锥形感受器,且占优势数量的感受器类型和分布规律也不相同。咖啡黑(枝)小蠹属多雌一雄型,新羽化的成虫在隧道内交配后,雌成虫飞出另找新的寄主场所,雄成虫继续生活在老隧道内直到死亡,雄成虫并不参与寄主定位[5]。光滑足距小蠹雌、雄成虫羽化后在原生长发育的坑道中交配,受精后的雌成虫分散外出,重新寻找寄主蛀孔侵入,仅雌成虫入侵寄主,雄成虫并不参与寄主定位和入侵[16]。咖啡黑(枝)小蠹与光滑足距小蠹为同属食菌小蠹,均为一雄多雌类型,说明两者的触角感受器类型相同、分布特点相似,在寄主选择和入侵行为等方面存在高度相似性。这与陈辉等[15]在华山松大小蠹成虫触角感受器的扫描电镜观察讨论的观点一致:小蠹虫触角感受器的类型和分布规律与寄主树木挥发性物质的识别和反应密切相关,具有相同感受器类型和分布规律的小蠹虫可能意味着具有相似的寄主感受行为和选择行为。
在咖啡黑(枝)小蠹成虫触角锤状部发现的表皮孔,与开口于圆形凸起上的腔锥形感器或瓶形感器不同,它的孔口直接开于触角表面,这与青杨脊虎天牛(Xylotrechus rusticus L.)触角第3~9鞭节上的表皮孔形态描述相似[23],分布在刺形感器基部附近,咖啡黑(枝)小蠹锤状部存在的表皮孔则分布于锯齿形感受器、毛形感受器等基部附近。类似的表皮孔在鞘翅目蚁甲科(Pselaphidae)、叶甲科(Chrysomelidae)中的某些种类中被描述[24]。有研究表明,这些表皮孔可能是信息素腺体,能够分泌触角和感器的滑润剂[25]。咖啡黑(枝)小蠹成虫触角表面存在的表皮孔和各种类型感受器的功能及在成虫寄主选择中的作用,还需进一步进行微观结构和电生理方面的研究。
参考文献
[1] 刘爱勤, 桑利伟, 孙世伟, 等. 热带香辛饮料作物主要病虫害防治图谱[M]. 北京: 中国农业出版社, 2013.
[2] 林延谋, 符悦冠, 刘凤花, 等. 咖啡黑小蠹的发生规律及药剂防治研究[J]. 热带作物学报, 1994, 15(2): 79-86.
[3] 吳跃开, 余金勇, 李晓虹, 等. 园林植物新害虫——黑色枝小蠹的调查[J]. 亚热带植物科学, 2007, 36(1): 13-16.
[4] 董云萍, 龙宇宙, 孙 燕. 咖啡高产栽培技术[M]. 北京: 中国农业出版社, 2009.
[5] 中国热带农业科学院, 华南热带农业大学. 中国热带作物栽培学[M]. 北京: 中国农业出版社, 1998.
[6] 陈 辉. 化学信息素对小蠹虫的入侵危害的调控[J]. 林业科学, 2003, 39(6): 154-158.
[7] 闫争亮. 小蠹科害虫化学信息物质及其对侵害寄主等行为的影响[J]. 西部林业科学, 2006, 35(3): 22-33.
[8] Byers J A. Host-tree chemistry affecting colonization in bark beetles, in CardeRT and Bell W J (eds) Chemical Ecology of Insects[M]. New York: Chapman and Hal, 1995: 154-213.
[9] Elkinton J S, Wood D L. Feeding and boring behavior of the bark beetle Ips paraconfusus(Coleoptera: scolytidae) on the Bark of a Host and Non-host Trees Species[J]. Can Entomo, 1980, 112: 797-809.
[10] 陈 辉, 李宗波. 植物挥发性化合物在小蠹虫寄主选择中的作用[J]. 福建林学院学报, 2006, 26(1): 87-91
[11] 苗振旺, 张钟宁, 王培新, 等. 外来入侵害虫红脂大小蠹对寄主挥发物的反应[J]. 昆虫学报, 2004, 47(3): 360-364.
[12] 周 楠, 毋亚梅, 张立新, 等. 松小蠹成虫对其聚集信息化合物的触角电位反应[J]. 云南林业科技, 2000, 9(3): 38-42.
[13] 毋亚梅, 周 楠, 张立新, 等. 松纵坑切梢小蠹成虫触角的扫描电镜观察[J]. 西南林学院学报, 2000, 20(1): 40-47.
[14] 王玉刚, 张 真, 王鸿斌, 等. 红脂大小蠹成虫触角扫描电镜的观察. 林业科学研究, 2003, 16(6): 726-730.
[15] 陈 辉, 李宗波, 唐 明. 华山松大小蠹成虫触角感受器的扫描电镜观察[J]. 林业科学, 2006, 42(11): 156-159.
[16] 杨群芳, 韩菊兰, 李 庆. 光滑足距小蠹成虫触角感受器的电镜扫描观察[J]. 昆虫知识, 2010(3): 520-524.
[17] 范丽华, 李永红, 张金桐, 等. 脐腹小蠹成虫触角结构与感受器扫描电镜观察[J]. 林业科学, 2011, 47(5): 87-91.
[18] 李宗波, 欧晓红. 小粒材小蠹成虫触角结构与感受器的扫描电镜观察[J]. 西北林学院学报, 2013, 28(3): 138-141.
[19] Schneider D. Insect antennae[J]. Annu Rev Entomol, 1964, 9: 103-122.
[20] 马瑞燕, 杜家纬. 昆虫的触角感器[J]. 昆虫知识, 2000, 37(3): 179-183.
[21] 殷惠芬, 黄复生, 李兆臃, 等. 中国经济昆虫志: 第29册[M]. 北京: 科学出版杜, 1984: 56-57.
[22] 宋丽文. 两种小蠹虫寄主选择机制研究[D]. 长春: 东北师范大学, 2010.
[23] 程 红. 青杨脊虎天牛触角感器类型及其对植物挥发物的反应电位反应[D]. 哈尔滨: 东北林业大学, 2006.
[24] Weis A, Schonitzer K, Melzer R R. Exocrine glands in the antennae of the carabid beetle, Platynus assimilis (Paykull) 1790 (Coleoptera, Carabidae, Pterostichinae)[J]. Insect Morphol and Embryol, 1999, 28(4): 331-335.
[25] Skilbeck C A, Anderson M. The fine structure of glandular units on the antennae of two species of the parasitoid, aleochara (Coleoptera: Staphylinidae)[J]. Insect Morphol and Embryol, 1994, 23(4): 319-328.
