姜黄素对非小细胞肺癌A459细胞增殖及CDH13甲基化状态的影响
2017-05-25赵莹李英姿
赵莹,李英姿
(徐州市中医院,江苏徐州221009)
姜黄素对非小细胞肺癌A459细胞增殖及CDH13甲基化状态的影响
赵莹,李英姿
(徐州市中医院,江苏徐州221009)
目的 观察姜黄素对非小细胞肺癌(NSCLC)A549细胞DNA甲基化转移酶(Dnmts)活性和抑癌基因人H-钙黏蛋白(CDH13)的影响,探讨姜黄素的去甲基化机制。方法 将A549细胞分为两组,观察组加入40 μmol/L姜黄素和培养液,对照组仅加入培养液。采用MTT法检测两组细胞增殖能力,RT-PCR方法检测CDH13 mRNA表达,甲基化特异性PCR(MSP)检测CDH13启动子CpG岛的甲基化状态,酶活性连续循环比色法检测Dnmt1、Dnmt3b活性。结果 观察组和对照组的细胞增殖OD值分别为0.638±0.184、0.859±0.040,观察组低于对照组(P<0.05)。两组CDH13 mRNA的相对表达量分别为0.64±0.09、0.43±0.12,观察组高于对照组(P<0.05)。对照组出现甲基化扩增条带,无去甲基条带;观察组除甲基化条带外,出现去甲基化条带。观察组和对照组每mg核蛋白中Dnmt1的活性分别为(0.153±0.016)、(0.770±0.014)μmol/min,Dnmt3b的活性分别为(0.137±0.020)、(0.179±0.013)μmol/min,观察组Dnmt1、Dnmt3b活性均较对照组下降(P均<0.05)。结论 姜黄素可通过降低Dnmts活性来抑制CDH13的甲基化状态,从而抑制A549细胞增殖。
非小细胞肺癌;姜黄素;甲基化转移酶;人H-钙黏蛋白;细胞增殖
DNA异常甲基化是诱发肺癌的重要原因。DNA甲基化是通过三种DNA甲基化转移酶(Dnmts)Dnmt1、Dnmt3a、Dnmt3b的催化实现的,其中Dnmt1、Dnmt3b的转录和活性增加是引起抑癌基因启动子CpG岛高甲基化的原因之一[1]。由于CpG岛甲基化导致抑癌基因失活是一个可逆转的表观遗传学基因修饰过程,去甲基化修饰可直接恢复抑癌基因的功能[2],因此Dnmts成为肿瘤治疗的热点靶向分子。阻断Dnmts能够阻止启动子CpG岛的甲基化,抑癌基因重新表达,从而抑制肿瘤生长。姜黄素是一种有效的抑癌中药提取物,对抑癌基因具有潜在的去甲基化作用[3,4]。2015年9月~2016年5月,我们应用姜黄素处理非小细胞肺癌(NSCLC)A549细胞,由于A549细胞中抑癌基因人H-钙黏蛋白(CDH13)是完全甲基化状态[5],因此以CDH13为信号,观察姜黄素的去甲基化效果,同时观察Dnmt1、Dnmt3b活性的变化,探讨姜黄素对A549细胞的去甲基化机制。
1 材料与方法
1.1 材料 A549细胞株购自上海细胞生物所。姜黄素粉剂为美国Sigma公司产品(纯度为98%,将其加入二甲基亚砜配置成10 mmol/L的溶液,-20 ℃避光存储,用前加入RPMI1640稀释至40 μmol/L)、通用型基因甲基化检测试剂盒、Dnmt1和Dnmt3b活性检测试剂盒购自上海Genmed公司;总RNA提取试剂盒TRIzol Reagent为美国Invitrogen公司产品;RT-PCR试剂盒为TaKaRa公司产品;DNA抽提试剂盒为美国Omega Bio-tec公司产品;RPMI1640培养基、核蛋白提取试剂盒及实验相关引物合成由南京凯基生物公司提供。
1.2 细胞培养 A549细胞株培养于含10%小牛血清、100 U/mL青霉素和100 U/mL链霉素的RPMI1640培养基中,培养条件为37 ℃、5% CO2。每2日换液1次,每3~4日消化传代1次。取对数生长期细胞用于实验。
1.3 细胞增殖能力观察 采用MTT法。取对数生长期细胞,调整细胞浓度为1×105/mL,接种于96孔培养板,每孔100 μL。将细胞分为两组,每组设6个复孔。观察组加入30 mL培养液及40 μmol/L姜黄素30 μL,对照组加入30 mL培养液。孵育24 h后,弃培养液,重复以上步骤二次处理细胞,加入3 mL清洗液清洗。收集细胞,加入MTT液(5 g/L)10 μL,37 ℃孵育4 h后加入100 μL二甲基亚砜孵育15 min,振荡器振荡10 min充分溶解,于分光光度仪上测定各孔490 nm处的吸光度OD值,减去空白孔的OD值,作为观察结果。
1.4 CDH13 mRNA和甲基化检测以及Dnmts活力检测
1.4.1 细胞培养与分组处理 取对数生长期细胞接种于培养瓶中,37 ℃培养箱孵育24 h。将细胞分为观察组和对照组,观察组加入30 mL培养液及40 μmol/L姜黄素30 μL,对照组加入30 mL培养液,孵育24 h后,弃培养液;重复以上步骤二次处理细胞,加入3 mL清洗液清洗,收集细胞用于以下实验。
1.4.2 CDH13 mRNA表达检测 采用半定量RT-PCR方法。两组分别收集1×106个细胞,加入TRIzol提取总RNA,逆转录获得cDNA。使用CDH13特异性引物进行PCR扩增,以β-actin作为内参。CDH13引物序列上游5′-GCAAGTCAGTTCAGACTCCAGCC-3′,下游5′-AGGCCTTTTGACTGTAATCACACC-3′,退火温度60 ℃,产物片段为393 bp。β-actin引物序列:上游5′-GAGAGGGAACTCGTGCGTGAC-3′,下游5′-CATCTGCTGGAAGGTGGACA-3′,退火温度55 ℃,产物片段为452 bp。取反应产物行琼脂糖凝胶电泳,凝胶成像系统分析。计算目的基因与β-actin的光密度比值作为CDH13 mRNA的相对表达量。实验重复3次,取平均值。
1.4.3 CDH13启动子CpG岛的甲基化检测 采用甲基化特异性PCR(MSP)方法。收集两组细胞,使用DNA提取试剂盒提取基因组DNA,产物经琼脂糖凝胶电泳,紫外分光光度计鉴定。使用甲基化检测试剂盒对提取的DNA进行磺化修饰和纯化,转化后的DNA样本溶于20 μL TE溶液中,-20 ℃保存。按照甲基化检测试剂盒说明书进行MSP检测,反应体系共25 μL,包括转化后的DNA模板1 μL(100 ng),15 mL扩增液,CDH13甲基化或去甲基化上下游引物各1 μL,双蒸水7 μL补齐反应体系总量至25 μL。反应条件:95 ℃预变性5 min;95 ℃ 30 s,退火30 s,72 ℃ 30 s,35个循环;72 ℃延伸5 min。CDH13甲基化引物序列:上游5′-TCGCGGGGTTCGTTTTTCGC-3′,下游5′-GACGTTTTCATTCATACACGCG-3′;CDH13去甲基化引物序列:上游5′-TTGTGGGGTTTGTTTTTTGT-3′,下游5′-AACTTTTCATTCATACACACA-3′。退火温度54 ℃,产物片段均为243 bp。取反应产物行琼脂糖凝胶电泳。以人类胎盘组织DNA经甲基化酶SssⅠ转化后作为甲基化的阳性对照,健康人淋巴细胞DNA作阴性对照,双蒸水作空白对照。
1.4.4 Dnmt1、Dnmt3b活性检测 使用核蛋白提取试剂盒提取两组细胞中的核蛋白,考马斯亮蓝法测定提取液蛋白浓度。用Dnmt1、Dnmt3b活性酶连续循环比色法定量检测试剂盒,取20 μg核蛋白检测处理前后细胞中Dnmt1、Dnmt3b的活性。Dnmts的特异性活性=样本总活性-样本非特异性活性。1个单位活性代表Dnmts在37 ℃,pH 8.0条件下,1 min可转移1 μmol S-腺苷甲硫氨酸甲基到半链甲基化寡核苷酸分子上。计算每mg核蛋白所含Dnmts的单位活性。实验重复3次,取平均值。
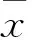
2 结果
2.1 两组细胞增殖能力比较 观察组和对照组的相对OD值分别为0.638±0.184、0.859±0.040,观察组低于对照组(P<0.05)。
2.2 两组CDH13 mRNA表达检测 观察组与对照组CDH13 mRNA的相对表达量分别为0.64±0.09、0.43±0.12,观察组高于对照组(P<0.05)。
2.3 两组CDH13甲基化状态比较 见图1。空白对照无甲基化和去甲基化产物;阴性对照可见去甲基化扩增条带(1u,243 bp);阳性对照可见甲基化扩增条带(2m,243 bp)。对照组可见甲基化扩增条带(4m),无去甲基条带;观察组除甲基化条带(3m)外,出现去甲基化条带(3u)。提示CDH13的启动子CpG岛在姜黄素的作用下发生了去甲基化。

注:0为空白对照,1为阴性对照,2为阳性对照,3为观察组,4为对照组。M为Marker,m为甲基化扩增,u为非甲基化扩增。
图1 MSP检测细胞处理前后CDH13基因启动子的甲基化状态
2.4 两组Dnmts活性比较 观察组和对照组每mg核蛋白中Dnmt1的活性分别为(0.153±0.016)、(0.770±0.014)μmol/min,Dnmt3b的活性分别为(0.137±0.020)、(0.179±0.013)μmol/min,观察组Dnmt1、Dnmt3b活性均较对照组明显下降(P均<0.05)。
3 讨论
DNA的甲基化和去甲基化修饰是重要的表观遗传学调控方式。DNA甲基化通过Dnmts催化实现,其中Dnmt1是主要的维持甲基化酶,Dnmt3b则催化从头甲基化,它们与正常的胚胎发育、衰老和肿瘤发生关系密切[2]。研究表明,Dnmts在肿瘤组织和肿瘤细胞系中呈过度表达[6],抑制或选择性沉默Dnmts可抑制肿瘤生长[7]。这种表观遗传学调控是可逆转的,因此寻找有效的去甲基化药物能够为肿瘤的靶向治疗提供新的方向。目前发现的去甲基化药物包括核苷类、氨基苯甲酸类、肼类、茶多酚类等,以及中药中新发现的砒霜(三氧化二砷)和姜黄素[8]。美国FDA已经批准上市了两种核苷类去甲基化药物,用于治疗骨髓异常增生类疾病[7]。但由于核苷类对正常细胞的细胞毒作用,影响了其临床应用。姜黄素作为一种低毒的中药提取物,为寻找低毒高效的抗肿瘤药物提供了新思路。
姜黄素是从莪术、姜黄等中草药中提纯出来的一种植物多酚,具有抗氧化、镇痛、抗炎和抗肿瘤等作用[3]。研究显示,姜黄素能够抑制胰腺癌[9]、乳腺癌[10]、结肠癌[11]、肺癌[12]、肝癌[13]等肿瘤生长,其抑癌机制包括调节信号传导途径、诱导细胞凋亡[13]、调控细胞周期[14]等。姜黄素是一种潜在的去甲基化药物,其对抑癌基因的去甲基化作用已引起人们的高度关注。研究显示,姜黄素可能具有多靶点的抗肿瘤机制。Xu等[12]发现,姜黄素衍生物可以诱导肺癌细胞自噬,促进其凋亡,这与抑制Hedgehog信号传导途径有关,而对正常肺泡上皮细胞无毒性作用。Li等[9]发现,姜黄素可通过下调NF-κB及其下游调控因子抑制人胰腺癌细胞生长,诱导其凋亡。Ke等[14]发现,姜黄素可下调Aurora-A,抑制乳腺癌细胞的有丝分裂,使其停滞在S期及G2/M期,从而阻止乳腺癌肺转移;姜黄素还可以增加乳腺癌细胞对顺铂、长春瑞滨等化疗药物的敏感性。
本研究应用40 μmol/L姜黄素处理A549细胞株后,发现细胞增殖受到抑制,甲基化特异性PCR提示细胞中完全甲基化的抑癌基因CDH13启动子CpG岛的甲基化状态部分逆转,CDH13 mRNA的转录水平提高。提示姜黄素对A549细胞株的抑制作用与其去甲基化作用有关,在此过程中Dnmt1和Dnmt3b的催化活性下降,说明姜黄素可能通过抑制Dnmt1和Dnmt3b的催化活性发挥去甲基化作用,Dnmts活性下降提示姜黄素在蛋白水平抑制了Dnmts的作用。Liu等[4]研究认为,姜黄素的去甲基化作用可能通过与Dnmt1的巯基共价结合而实现的,Dnmt1的甲基转移位点被姜黄素占据,导致其转移甲基的效率下降,妨碍了基因启动子的甲基化修饰,从而起到去甲基化作用。Liu等[15]发现,姜黄素的衍生物双去甲氧基姜黄素可以使A549细胞株中的抑癌基因WIF-1表达增加,具有较强的去甲基化作用。Chamani等[10]发现,使用姜黄素衍生物处理肝癌细胞株后,Dnmts的转录水平提高,抑癌基因miR-34家族miR-34a、miR-34b和miR-34c的转录也明显上升。以上研究表明,姜黄素对癌细胞株的去甲基化作用可能是通过多种途径实现的。
综上所述,姜黄素具有去甲基化作用,能够提高抑癌基因CDH13的表达,降低甲基化转移酶Dnmts的活性。由于肿瘤的发生发展及抗癌药物的作用机制极其复杂,体外肿瘤细胞株的研究不能完全复制体内肿瘤细胞的生活状态。因此,姜黄素的去甲基化作用还需要更多的实验支持。此外,姜黄素的生物利用度低、稳定性差、在体内代谢快等缺陷,在一定程度上影响了其应用。姜黄素的多种天然和合成的衍生物,如去甲氧基姜黄素、双去甲氧基姜黄、含氮杂环姜黄素等,较姜黄素更稳定,抗肿瘤作用也得到了实验验证[14]。我们下一步的研究将着重于姜黄素及其衍生物在动物体内的去甲基化作用,期待以姜黄素作为前体的衍生物成为一种高效、低毒的靶向治疗药物,在肿瘤治疗领域发挥更重要的作用。
[1] Rhee I, Bachman KE, Park BH, et al. DNMT1 and DNMT3b cooperate to silence genes in human cancer cells[J]. Nature, 2002,416(6880):552-556.
[2] Klutstein M, Nejman D, Greenfield R, et al. DNA Methylation in Cancer and Aging[J]. Cancer Res, 2016,76(12):3446-3450.
[3] Shehzad A, Lee J, Lee YS. Curcumin in various cancers[J]. Biofactors, 2013,39(1):56-68.
[4] liu Z, Xie Z, Jone W, et al. Curcumin is a potent DNA hypomethylation agent[J]. Bioorg Med Chem Lett, 2009,19(3):706-709.
[5] Daskalos A, Oleksiewicz U, Filia A, et al. UHRF1-mediated tumor suppressor gene inactivation in nonsmall cell lung cancer[J]. Cancer, 2011,117(5):1027-1037.
[6] Schmidt WM, Sedivy R, Forstner B, et al. Progressive up-regulation of genes encoding DNA methyltransferases in the colorectal adenoma-carcinoma sequence[J]. Mol Carcinog, 2007,46(9):766-772.
[7] Ghoshal K, Bai S. DNA methyltransferases as targets for cancer therapy[J]. Drugs Today(Barc),2007,43(6):395-422
[8] 陈淑瑜.DNA去甲基化药物研究现状[J].齐齐哈尔医学院学报,2013,34(4):559-561.
[9] Li L, Aggarwal B, Shishodia S, et al. Nuclear factor-kappaB and ikappaB kinase are constitutively active in human pancreatic cells, and their down-regulation by curcumin (diferuloylmethane) is associated with the suppression of proliferation and the induction of apoptosis[J]. Cancer, 2004,101(10): 2351-2362.
[10] Bimonte S, Barbieri A, Palma G, et al. Dissecting the role of curcumin in tumour growth and angiogenesis in mouse model of human breast cancer[J]. Biomed Res Int, 2015(2015),878134.
[11] Half E, Arber N. Colon cancer: preventive agents and the present status of chemoprevention[J]. Expert Opin Pharmacother, 2009,10(2):211-219.
[12] Xu J, Yang H, Zhou X. et al. Autophagy accompanied with bisdemethoxycurcumin-induced apoptosis in non-small cell lung cancer cells[J]. Biomed Environ Sci, 2015,28(2):105-115.
[13] Chamani F, Sadeghizadeh M, Masoumi M, et al. Evaluation of MiR-34 family and DNA methyltransferases 1, 3A, 3B gene expression levels in hepatocellular carcinoma following treatment with dendrosomal nanocurcumin[J]. Asian Pac J Cancer Prev, 2016,17:219-224.
[14] Ke CS, Liu HS, Yen CH, et al. Curcumin-induced Aurora-A suppression not only causes mitotic defect and cell cycle arrest but also alters chemosensitivity to anticancer drugs[J]. J Nutr Biochem, 2014,25(5):526-539.
[15] Liu YL, Yang HP, Gong L, et al. Hypomethylation effects of curcumin, demethoxycurcumin and bisdemethoxycurcumin on WIF-1 promoter in non-small cell lung cancer cell lines[J]. Mol Med Rep, 2011,4(4):675-679.
Effect of curcumin on proliferation and CDH13 methylation status of non-small-cell lung cancer cell line A549
ZHAOYing,LIYingzi
(XuzhouHospitalofTraditionalChineseMedicine,Xuzhou221009,China)
Objective To observe the effect of curcumin on DNA methyltransferase (Dnmts) activity and tumor suppressor gene human H-cadherin (CDH13) in non-small-cell lung cancer (NSCLC) A549 cells and to explore the mechanism of curcumin's demethylation.Methods A549 cells were divided into two groups: the observation group and the control group. Cells in the observation group were cultured with 40 μmol/ L curcumin and the culture medium, while cells in the control group only with the culture medium. MTT assay was used to detect the cell proliferation, and RT-PCR was used to detect the expression of CDH13 mRNA. CpG island methylation status of CDH13 promoter was detected by methylation-specific PCR (MSP), and catalytic activity of Dnmts was detected by enzyme colorimetric determination kit. Results The OD of the observation group was 0.638±0.184, which was lower than that (0.859±0.040) of the control group (P<0.05). The relative expression level of CDH13 mRNA in the observation group was 0.64±0.09, which was higher than that (0.43±0.12) of the control group. There were methylation amplification bands, but no demethylation bands in the control group, and there were both methylation amplification bands and demethylation bands in the observation group. In the observation group and the control group, the activities of Dnmt1 were separately (0.153±0.016) and (0.770±0.014) μmol/min, and the activities of Dnmt3b were (0.137±0.020) and (0.179±0.013) μmol/min, respectively. The activities of Dnmt1 and Dnmt3b in the observation group were lower than those in the control group (allP<0.05). Conclusion Curcumin inhibits the methylation status of CDH13 by decreasing the activity of Dnmts, and thereby inhibits A549 cell proliferation.
non-small-cell lung cancer; curcumin; methyltransferase; human H-cadherin; cell proliferation
江苏省中医药局科技项目(YX1230)。
赵莹(1986-),女,主治医师,主要研究方向为呼吸道肿瘤的诊治。E-mail: njmuzy@126.com
李英姿(1968-),女,主任医师,主要研究方向为肿瘤的中医诊治。E-mail: lblyz652@sohu.com
10.3969/j.issn.1002-266X.2017.06.003
R734.2
A
1002-266X(2017)06-0009-04
2016-08-16)
