脂多糖诱导巨噬细胞中核受体协同抑制因子启动子甲基化对炎症因子的调控作用
2017-05-15郭小莉刘霞雷传江王关嵩王建春
郭小莉 刘霞 雷传江 王关嵩 王建春
·论著·
脂多糖诱导巨噬细胞中核受体协同抑制因子启动子甲基化对炎症因子的调控作用
郭小莉 刘霞 雷传江 王关嵩 王建春
目的探讨核受体协同抑制因子(NCOR)在脂多糖(LPS)诱导巨噬细胞炎症反应中的作用及其调控机制。方法1 μg/ml的LPS分别处理小鼠巨噬细胞RAW264.7 24 h和48 h,应用Western blot和Real time-PCR检测NCOR的表达水平以及肿瘤坏死因子-α(TNF-α)、白细胞介素-6(IL-6) mRNA水平,荧光素酶报告基因检测核因子-κB (NF-κB)的启动子活性。LPS处理细胞48 h后,应用MSP检测NCOR启动子是否发生甲基化以及Western blot检测DNMT3b的表达变化。Real time-PCR检测5′-aza和LPS联合处理细胞后NCOR mRNA的表达水平;转染DNMT3b siRNA后,分别应用Western blot和Real time-PCR检测DNMT3b的表达水平,以及DNMT3b siRNA和LPS联合作用下NCOR、TNF-α、IL-6的表达水平和NF-κB的启动子活性。结果LPS干预细胞24、48 h后,NCOR蛋白和mRNA表达显著下调(P<0.05),而TNF-α、IL-6 mRNA表达水平、DNMT3b 蛋白的表达水平以及NF-κB的启动子活性显著上升(P<0.05)。MSP检测说明LPS可介导NCOR的启动子甲基化。用5′-aza和LPS联合处理细胞后NCOR mRNA水平较LPS组有显著上升(P<0.05)。采用DNMT3b siRNA可显著下调DNMT3b 蛋白和mRNA水平,并可部分逆转LPS介导的抑制NCOR表达的效应,抑制TNF-α、IL-6的表达水平和NF-κB的启动子活性(P<0.05)。结论NCOR启动子的甲基化是LPS介导巨噬细胞炎症反应发生、发展的关键步骤,其可作为治疗ALI/ARDS的潜在靶点。
核受体协同抑制因子; DNA 甲基化; 核转录因子κB; 炎症因子
急性肺损伤(acute lung injury, ALI)和急性呼吸窘迫综合征(acute respiratory distress syndrome, ARDS)严重威胁着人民的生命安全[1-3]。尽管新型药物及机械通气等措施得到广泛的应用,但是ALI/ARDS的病死率仍居高不下[4]。严重的炎症反应如脓毒血症是引起ALI/ARDS主要原因之一[5]。而在这个过程中,受到内毒素刺激的肺泡巨噬细胞通过大量释放炎症介质,在肺组织损伤中发挥着关键的作用[5-6]。因此,深入探究肺巨噬细胞在炎症反应中的分子机制,可为ALI/ARDS的防治提供新的防治策略和靶点。
核受体协同抑制因子(nuclear receptor corepressor, NCOR)是一种核转录抑制因子,其通过调控多种转录因子发挥作用。Hong等[7]研究发现下调NCOR表达可通过促进过氧化物酶体增生物激活受体γ(peroxisome proliferator-activated receptor gamma, PPARγ)的表达,进而发挥负性调节间充质干细胞向脂肪细胞的分化。其他研究也表明NCOR在调控肿瘤的相关生物学特性方面发挥着重要的作用[8-9]。而在炎症反应中,通过抑制NCOR的表达,可促进炎症因子释放的作用,但是否存在其他机制尚不清楚[10]。
DNA甲基化是一种常见的基因修饰途径之一。其主要是在DNA甲基转移酶(DNA methyltransferases, DNMTs)的催化下,完成对染色质结构、DNA构象、DNA稳定性及DNA与蛋白质相互作用方式的改变,控制基因表达,进而引起疾病的发生、发展[11-14]。既往研究表明在ALI大鼠模型中上调DNMT3b、DNMT1等,可引起PPAR γ等基因的启动子甲基化,进而在ALI的进展中发挥着重要作用[15-16]。本研究旨在探讨LPS是否可诱导NCOR的甲基化,及其在NCOR甲基化中的机制,及在巨噬细胞介导的炎症反应中的作用。
材料和方法
一、实验材料
DMEM高糖培养基和胎牛血清(无内毒素)购自GIBCO公司;脂多糖(lipopolysaccharide, LPS)和5-氮胞苷2′-脱氧胞苷(5-Aza-2′-deoxycytidine,5′-aza)和TRIzol reagent购自sigma公司;兔源NCOR单克隆抗体、兔源DNMT3b单克隆抗体、兔源GAPDH多克隆抗体以及山羊抗兔HRP-Ig(H+L)二抗购自江苏睿瀛生物科技有限公司;逆转录试剂盒购自Thermo fisher公司;定量PCR mix购自罗氏公司;DNA提取试剂盒、DNA重亚硫酸盐转化试剂盒和甲基化特异性PCR试剂盒均购自天根生化科技有限公司。pNF-κB-luc报告基因质粒、G418和荧光素酶报告基因检测试剂盒购自江苏碧云天生物技术有限公司。DNMT3b siRNA和阴性对照(negative control, NC)由广州锐博生物科技有限公司设计和合成;转染试剂lipofiter购自汉恒(上海)生物科技有限公司。
二、研究方法
1. 小鼠巨噬细胞RAW264.7的培养:小鼠巨噬细胞RAW264.7由第三军医大学生物测试中心提供。RAW264.7用DMEM+10%FBS的培养基进行培养。当细胞融合度达到90%时使用细胞刮刀将细胞刮下来,离心后重新进行传代。细胞传代后到达对数生长期进行相关实验。
2. LPS处理RAW264.7细胞:使用1 μg/ml的LPS分别处理对数生长期的RAW264.7细胞0 h、24 h和48 h,然后进行后续试验[16]。
3. 5′-aza处理RAW264.7细胞:使用1 μM的5′-aza分别处理对数生长期的RAW264.7细胞48 h,然后进行后续试验。
4. pNF-κB-luc报告基因质粒的稳定转染:将消化下来的RAW264.7细胞传代到10 cm2培养皿中,培养12 h使细胞融合度达到80%。弃去培养基加入10 ml的DMEM。将24 μg pNF-κB-luc质粒加入1 ml DMEM中,充分混匀。再取一个EP管,加入1 ml的DMEM后,再加入72 μl lipofiter,充分混匀。将上述两管静置5 min后,混合在一起,充分混匀后再静置15 min。将上述混合液均匀滴入细胞培养基中,培养6 h后换成还有DMEM+10%FBS的完全培养基继续培养48 h。之后要用1 mg/ml的G418持续筛选1个月,获得稳定表达NF-κB-luc-RAW264.7细胞。
5. 甲基化特异性PCR(methylating-specific PCR, MSP):将处理后的RAW264.7细胞用PBS洗涤1次后,用细胞刮刀将细胞刮下,离心。用DNA提取试剂盒提取细胞的DNA,使用DNA重亚硫酸盐转化试剂盒将DNA样品中未甲基化胞嘧啶转化为尿嘧啶。非甲基化引物(unmethylation primer):上游:5′-TTGTTTGTGTTATTTATGTTTATGA -3′,下游: 5′-CAATACTTACTACTACCCACATCAAT-3′。甲基化引物(methylation primer): 上游:5′-TTGTTTGTGTTATTTGTGTTTACGA-3′,下游: 5′-ATACTTACTACTACCCGCGTCGAT-3′,见表1。将上述样本就行MSP,扩增条件:95 ℃ 3 min预变性,95 ℃ 20 s,60 ℃ 20 s,72 ℃ 20 s进行35个循环,再以72 ℃ 5 min进行延伸。将上述产物进行琼脂糖电泳。
6. DNMT3b siRNA转染RAW264.7细胞:将3条DNMT3b siRNA和对照组 (NC)用RNase-free H2O 配制成20 nM的储存液后,分装、冻存备用。使用终浓度为50 pM DNMT3b siRNA转染细胞的简要操作如下:①将5×104/孔 RAW264.7接种到24孔板中,培养12 h;②2.5 μl 浓度为20 nM的DNMT3b siRNA加入到50 μl DMEM中,轻轻混匀;③2.4 μl的lipofiter 加入到50 μl DMEM中,轻轻混匀;④上述两组混悬液静置5 min后,再将它们混合在一起,充分混匀,静置15 min;⑤将100 μL混合试剂加入含有细胞的孔板中,轻轻混匀,DNMT3b siRNA的转染终浓度为50 pM;⑥将培养板置于37 ℃、5% CO2、饱和湿度下的培养箱中培养48 h后进行PCR和Western blot等检测。NC的转染同前。
7. 荧光素酶活性的检测:将作为内参pRL-TK质粒和DNMT3b siRNA/NC 共同转染稳定表达NF-κB-luc的RAW264.7细胞24 h后,再用1 μg/ml LPS干预24 h。将细胞用PBS洗涤1~2次,然后加入200 μl报告基因细胞裂解液,充分裂解细胞后,按照说明书加入溶解荧光素酶检测试剂,孵育后并进行检测。
8. Real time-PCR检测NCOR、DNMT3b、IL-6和TNF-α mRNA的表达:收集细胞后,用TRIzol提取总RNA,并逆转录为cDNA. cDNA采用BIO-RAD CFX96系统进行实时荧光定量PCR检测NCOR、DNMT3b、IL-6和TNF-α mRNA的表达,以β-actin作为内参。每个待测基因设4个复孔。数据通过BIO-RAD CFX96系统进行处理,按照公式RQ=2-ΔΔCT计算各组间的倍数关系。
9. Western blot检测NCOR、DNMT3b、IL-6和TNF-α mRNA的表达:收集细胞,加入300 μl蛋白裂解液,在冰上充分裂解,然后4 ℃、以离心半径8 cm,12 000 r/min离心20 min。取上清液,测定蛋白浓度,并将所有样本调整到相同浓度后,加入5×loading buffer,沸水中煮10 min使其变性。制备分离胶为8%的SDS-PAGE凝胶,每孔加样40 μg进行电泳。300 mA湿转150 min后,将PVDF膜室温封闭1 h,应用NCOR(1︰2 000)、DNMT3b(1︰2 000)及GAPDH抗体(1︰10 000)的抗体4 ℃孵育过夜;再孵育放入辣根酶标记山羊抗兔的二抗(1︰50 000),用ECL化学发光法检测NOCR和DNMT3b的表达水平。所得蛋白条带图片,使用Quality one软件对图像进行分析,以GAPDH的光密度值作为内参来校正各自目的蛋白的光密度值。

表1 基因引物序列
三、统计学方法
各实验至少重复3次,使用SPSS 17.0对数据进行分析,两组间进行t检验,多组之间进行单因素方差分析,P<0.05为具有统计学差异。
结 果
一、 LPS处理RAW264.7细胞后NCOR、IL-6和TNF-α表达以及NF-κB活性的变化
应用1 μg/ml LPS分别干预RAW264.7细胞0 h、24 h和48 h后,Western blot和real time-PCR检测发现如图1A和1B所示,LPS干预24 h和48 h后,RAW264.7细胞中NCOR蛋白和mRNA表达水平显著低于0 h组(P<0.05),但24 h和48 h组NCOR蛋白和mRNA表达水平无显著差异(P>0.05)。荧光素酶报告基因结果显示如图1C所示,LPS干预RAW264.7细胞后24 h、48 h,NF-κB 的活性较0 h组显著升高(P<0.05),并且48 h组NF-κB 的活性显著高于24 h(P<0.05)。此外,炎症因子TNF-α和IL-6 mRNA的表达水平随着LPS作用时间延长而升高(图1D),呈时间依赖(P<0.05)。

图1 LPS处理RAW264.7细胞后NCOR、IL-6和TNF-α表达以及NF-κB活性的变化,1 μg/ml LPS分别处理RAW264.7细胞0 h、24 h、48 h;注: A. Western blot检测NCOR蛋白的表达水平;B: Real time-PCR检测NCOR mRNA的表达水平;*与0 h比较,P<0.05;C:荧光素酶报告基因检测NF-κB启动子活性;*与0 h比较,P<0.05,^与24 h比较,P<0.05;D: Real time-PCR检测IL-6和TNF-α mRNA的表达水平;*与0 h比较,P<0.05,^与24 h比较,P<0.05
二、LPS处理RAW264.7细胞后对NCOR基因启动子甲基化的作用
应用MSP检测发现,见图2A: 1 μg/ml LPS干预RAW264.7细胞48 h后NCOR基因出现甲基化,提示LPS处理可诱导NCOR的启动子甲基化。同时,利用Real time-PCR检测NCOR mRNA的表达,见图2B:发现LPS处理48 h后,NCOR mRNA的表达较对照组显著降低(P<0.05)。但在使用DNA甲基化清除剂5′-aza和LPS同时处理后,NCOR mRNA的表达水平较对照组显著降低(P<0.05),但较LPS组的表达水平有显著的提升(P<0.05)。此外,还检测了DNMT3b的蛋白水平(图2C),发现其表达水平在LPS处理后较对照组显著升高(P<0.05)。
三、下调DNMT3b对NCOR表达的影响
RAW264.7细胞分别转染了3条siRNA和NC 48 h后,用Real time-PCR和Western blot检测发现,3条DNMT3b siRNA均可以下调DNMT3b mRNA和蛋白的表达(P<0.05),其中DNMT3b siRNA-2的下调作用最为显著(图3A和3B)。因此,接下来的实验均使用该条siRNA。进一步研究发现单独转染DNMT3b和NC,NCOR mRNA 和蛋白表达水平无显著的差异(P>0.05)(图3C和3D)。但RAW264.7细胞转染DNMT3b siRNA 24 h后,再用LPS处理24 h后,NCOR的表达水平较NC+LPS组显著降低,但仍然较单独siRNA组显著降低(P<0.05)。



图2 LPS处理RAW264.7细胞对NCOR 启动子甲基化的影响;注:A:MSP检测1 μg/ml LPS处理细胞48 h后,NCOR 启动子甲基化情况。U代表非甲基化NCOR DNA片段;M代表甲基化NCOR DNA片段;B:Real time-PCR检测NCOR mRNA的表达水平。*与control组比较,P<0.05;^与LPS组比较,P<0.05;C: Western blot检测1 μg/ml LPS处理细胞48 h后 DNMT3b蛋白的表达水平
四、下调DNMT3b对NF-κB活性及对IL-6和TNF-α表达的影响
利用荧光素酶报告基因检测NF-κB启动子活性, Real time-PCR检测IL-6和TNF-α mRNA的表达水平,发现单独DNMT3b siRNA组、NC+LPS组和DNMT3b siRNA +LPS组的NF-κB启动子活性及IL-6、TNF-α mRNA的表达水平均显著高于单独NC组(P<0.05)。DNMT3b siRNA+LPS组的NF-κB启动子活性及IL-6、TNF-α mRNA的表达水平较NC+LPS组明显降低(P<0.05),但仍高于单独DNMT3b siRNA组(P<0.05) (图4A、B)。



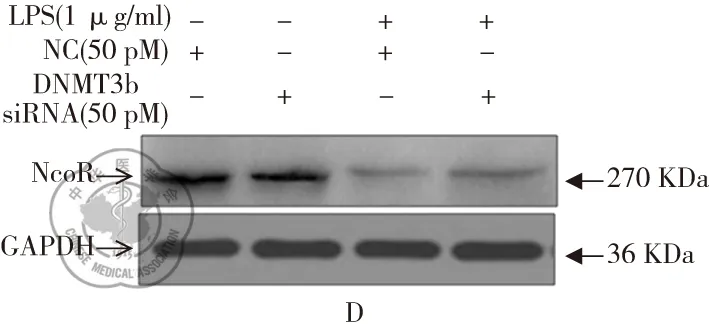
图3 下调DNMT3b对NCOR表达的影响;注:A和B: Real time-PCR和 western blot检测DNMT3b siRNA及NC转染RAW264.7细胞后,NCOR mRNA和蛋白的表达水平,*与NC组比较,P<0.05;C和D: DNMT3b siRNA转染24 h后再用1 μg/ml LPS处理24 h,Real time-PCR和 western blot检测NCOR mRNA和蛋白的表达水平,*与NC组比较,P<0.05;^DNMT3b siRNA组比较,P<0.05;#NC+LPS组比较,P<0.05
讨 论
本研究发现LPS可诱导巨噬细胞RAW264.7中NCOR的启动子甲基化,进而下调NCOR的表达,提升NF-κB的活性,导致炎症因子TNF-α、IL-6表达显著上调。进一步研究表明DNMT3b参与了NCOR的启动子甲基化。下调DNMT3b可部分逆转LPS介导的NCOR甲基化,并抑制NF-κB的活性和炎症因子TNF-α、IL-6表达的水平。上述研究说明,LPS介导NCOR的DNA 甲基化是促进巨噬细胞产生和释放炎症介质的关键环节。
失控性炎症反应是ALI/ARDS发生、发展的重要环节,而巨噬细胞在其中发挥着重要的作用[17]。当内毒素刺激肺泡巨噬细胞时,LPS将激活细胞膜表面的Toll样受体及下游信号通路NF-κB,产生大量的炎症因子如TNF-α、IL-6和IL-1β以及ROS,进而导致组织损伤。本研究也发现了同样的现象:在LPS刺激下,RAW264.7细胞的NF-κB的启动子活性显著增加以及炎症因子TNF-α和IL-6表达上调。上述结果说明本研究LPS诱导巨噬细胞炎症反应的模型是成功的,为进一步的研究奠定了基础。


图4 下调DNMT3b对NF-κB活性及IL-6和TNF-α表达的影响;注:A: 荧光素酶报告基因检测NF-κB启动子活性;B: Real time-PCR检测IL-6和TNF-α mRNA的表达水平,*与NC组比较,P<0.05;^DNMT3b siRNA组比较,P<0.05;#NC+LPS组比较,P<0.05
NCOR作为核转录抑制因子在炎症反应中发挥着重要作用。在正常状态下,NCOR与一些炎症通路的基因相结合,使它们处于抑制状态[18]。当炎症通路被激活后,NCoR被降解或从炎症基因的启动子上游离出来,消除对NF-κB、AP1等的转录抑制作用,进而释放大量炎症因子,加剧炎症反应[17,19]。本研究中,LPS处理后,NCoR的表达显著下降,而NF-κB的活性以及炎症因子的表达水平显著升高。而当NCOR的表达水平在DNMT3b siRNA处理下有所上调时,NF-κB的活性以及炎症因子的表达水平同样受到了部分抑制,这与文献报道一致。上述研究说明LPS可调控NCOR的表达和定位,其可能的机制是通过激活泛素化途径发挥作用[20-21]。有趣的是,本研究发现在LPS处理的巨噬细胞中NCOR发生了启动子甲基化。当使用DNA甲基化清除剂5′-aza和LPS共同处理巨噬细胞时,NCOR的表达水平较之前的LPS单独处理时有明显的升高,再次说明LPS可诱导NCOR的启动子甲基化,进而抑制其表达。
DNMTs是介导DNA甲基化的关键酶,其主要包括DNMT1、DNMT2和DNMT3三类。这三类酶由于mRNA剪切片段的不同又分为几个亚型(如DNMT3a、3b)[22],不同的亚型发挥着不同的作用[23]。前期研究和其他研究说明在急性肺损伤模型中,DNMT3b在介导PPARγ等基因的启动子甲基化中发挥着重要作用[15-16]。本研究同样发现LPS可诱导DNMT3b的表达。而通过siRNA下调DNMT3b后,可部分显著恢复下调的NCOR表达水平,进而逆转了炎症通路的激活和炎症因子的上调。上述结果说明DNMT3b参与了LPS介导的NCOR 基因启动子甲基化。
综上所述,本研究进一步明确了NCOR在LPS诱导的巨噬细胞炎症反应中的作用及机制,并首次发现了LPS介导NCOR基因的启动子甲基化。研究结果为炎症反应的发生、进展提供了新的理论基础,为ALI/ARDS的防治提供了新的靶点和策略。
1 宋旸, 蒋昊翔, 张永红, 等. 急性呼吸窘迫综合征药物研究进展[J/CD]. 中华肺部疾病杂志(电子版), 2015, 8(6): 769-772.
2 Rubenfeld GD, Caldwell E, Peabody E, et al. Incidence and outcomes of acute lung injury[J]. N Engl J Med, 2005, 353(16): 1685-1693.
3 Zambon M, Vincent JL. Mortality rates for patients with acute lung injury/ARDS have decreased over time[J]. Chest, 2008, 133(5): 1120-1127.
4 Villar J, Sulemanji D, Kacmarek RM. The acute respiratory distress syndrome: incidence and mortality, has it changed?[J]. Curr Opin Crit Care, 2014, 20(1): 3-9.
5 Opal SM, Dellinger RP, Vincent JL, et al. The next generation of sepsis clinical trial designs: what is next after the demise of recombinant human activated protein C?*[J]. Crit Care Med, 2014, 42(7): 1714-1721.
6 Matthay MA, Ware LB, Zimmerman GA. The acute respiratory distress syndrome[J]. J Clin Invest, 2012, 122(8): 2731-2740.
7 Hong-Wei G, Lan L, De-Guo X, et al. NCoR negatively regulates adipogenic differentiation of mesenchymal stem cells[J]. In Vitro Cell Dev Biol Anim, 2015, 51(7): 749-758.
8 Wong MM, Guo C, Zhang J. Nuclear receptor corepressor complexes in cancer: mechanism, function and regulation[J]. Am J Clin Exp Urol, 2014, 2(3): 169-187.
9 Heldring N, Nyman U, Lönnerberg P, et al. NCoR controls glioblastoma tumor cell characteristics[J]. Neuro Oncol, 2014, 16(2): 241-249.
10 Jennewein C, Kuhn AM, Schmidt MV, et al. Sumoylation of peroxisome proliferator-activated receptor gamma by apoptotic cells prevents lipopolysaccharide-induced NCoR removal from kappaB binding sites mediating transrepression of proinflammatory cytokines[J]. J Immunol, 2008, 181(8): 5646-5652.
11 Barrès R, Yan J, Egan B, et al. Acute exercise remodels promoter methylation in human skeletal muscle[J]. Cell Metab, 2012, 15(3): 405-411.
12 Crevillén P, Yang H, Cui X, et al. Epigenetic reprogramming that prevents transgenerational inheritance of the vernalized state[J]. Nature, 2014, 515(7528): 587-590.
13 Qiu W, Baccarelli A, Carey VJ, et al. Variable DNA methylation is associated with chronic obstructive pulmonary disease and lung function[J]. Am J Respir Crit Care Med, 2012, 185(4): 373-381.
14 Chouliaras L, Mastroeni D, Delvaux E, et al. Consistent decrease in global DNA methylation and hydroxymethylation in the hippocampus of Alzheimer′s disease patients[J]. Neurobiol Aging, 2013, 34(9): 2091-2099.
15 Zhang XQ, Lv CJ, Liu XY, et al. Genome wide analysis of DNA methylation in rat lungs with lipopolysaccharide induced acute lung injury[J]. Mol Med Rep, 2013, 7(5): 1417-1424.
16 Lei C, Jiao Y, He B, et al. RIP140 down-regulation alleviates acute lung injury via the inhibition of LPS-induced PPARγ promoter methylation[J]. Pulm Pharmacol Ther, 2016, 37: 57-64.
17 Lin KL, Suzuki Y, Nakano H, et al. CCR2+ monocyte-derived dendritic cells and exudate macrophages produce influenza-induced pulmonary immune pathology and mortality[J]. J Immunol, 2008, 180(4): 2562-2572.
18 Medzhitov R, Horng T. Transcriptional control of the inflammatory response[J]. Nat Rev Immunol, 2009, 9(10): 692-703.
19 Ogawa S, Lozach J, Jepsen K, et al. A nuclear receptor corepressor transcriptional checkpoint controlling activator protein 1-dependent gene networks required for macrophage activation[J]. Proc Natl Acad Sci U S A, 2004, 101(40): 14461-14466.
20 Lu R, Hu X, Zhou J, et al. COPS5 amplification and overexpression confers tamoxifen-resistance in ERα-positive breast cancer by degradation of NCoR[J]. Nat Commun, 2016, 7: 12044.
21 Glass CK, Saijo K. Nuclear receptor transrepression pathways that regulate inflammation in macrophages and T cells[J]. Nat Rev Immunol, 2010, 10(5): 365-376.
22 Xu F, Mao C, Ding Y, et al. Molecular and enzymatic profiles of mammalian DNA methyltransferases: structures and targets for drugs[J]. Curr Med Chem, 2010, 17(33): 4052-4071.
(本文编辑:黄红稷)
郭小莉,刘霞,雷传江,等. 脂多糖诱导巨噬细胞中核受体协同抑制因子启动子甲基化对炎症因子的调控作用[J/CD]. 中华肺部疾病杂志(电子版), 2017, 10(2): 157-162.
Lipopolysaccharide modulation the promoter methylation of nuclear receptor corepressor regulated inflammation mediators in macrophages
GuoXiaoli,LiuXia,LeiChuanjiang,WangGuansong,WangJianchun.
DepartmentofRespiratoryDisease,XinqiaoHospital,ThirdMilitaryMedicalUniversity,Chongqing400037,China
WangJianchun,Email:wjc5577@126.com
Objective To investigate the role and potential mechanism of nuclear receptor corepressor (NOCR) at lipopolysaccharide(LPS) mediation inflammation response in macrophages. Methods Western blot, realtime-PCR and luciferase assays was used to detect the expression of NCOR, interleukin-6(IL-6) and tumour necrosisfactor(TNF-α) and promoter activity of nuclear factor-κB (NF-κB) when RAW264.7 cells were treated with 1 μg/ml LPS. The promoter methylation of NCOR was analyzed by methylating-specific PCR(MSP) analysis, and the protein of DNMT3b was evaluated by western blot after cells stimulation with 1 μg/ml LPS for 48 h. Real time-PCR was also used to evaluate the expression of NCOR mRNA when RAW264.7 was treated with 1 μg/ml LPS combine with 1μM 5′-aza. After DNMT3b siRNA was transfected into RAW264.7 for 48 h, the mRNA and protein of DNMT3b was detected by western blot and real time-PCR. Additional, the expression of NCOR, TNF-α and IL-6 and NF-κB activity was analyzed after RAW264.7 was treated with 1 μg/ml LPS combine with DNMT3b siRNA. Results The expression of NCOR mRNA and protein was significantly decreased (P<0.05) and the expression of TNF-α and IL-6 mRNA, DNMT3b protein and activity of NF-κB was elevated (P<0.05) after cells was treated with 1 μg/ml LPS for 24 and 48 h, respectively. MSP assay showed LPS could modulate the NCOR promoter methylation. The expression of DNMT3b mRNA and protein was decreased after cells were transfected by DNMT3b siRNA. Additionally, down-regulation of DNMT3b was partly reversed LPS modulation the inhibition of NCOR, and decreased the expression of TNF-α and IL-6 mRNA, and activity of NF-κB (P<0.05). Conclusion NCOR promoter methylation is key step of occurrence and development of LPS mediation inflammation response. It might be a potential target for acute lung injury / acute respiratory distress syndrome (ALI/ARDS) treatment.
Nuclear receptor corepressor; DNA methylation; Nuclear factor-k-gene binding protein; Inflammatory factor
10.3877/cma.j.issn.1674-6902.2017.02.009
国家自然科学基金资助项目(81170066)
400037 重庆,第三军医大学新桥医院呼吸内科
王建春,Email: wjc5577@126.com
R563
A
2017-02-07)
