Red/ET同源重组技术在载体构建中的应用
2017-03-12郭善军张晓林陈章宝曾吉
郭善军 张晓林 陈章宝 曾吉
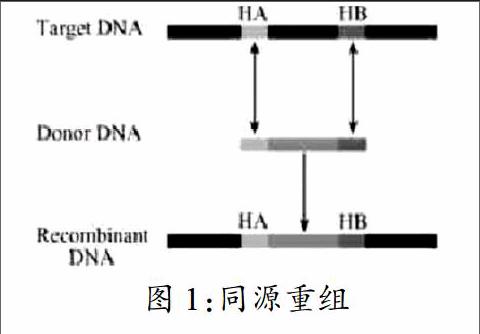
摘 要 Red/ET同源重组技术是以原有的重组技术为基础,利用Rac噬菌体ET系统或者 噬菌体的Red系统进行外源基因重组的重组方法。该方法使同源重组过程变得更方便快捷。本文对Red/ET同源重组技术的原理,改进和载体构建方面的应用做了综述。
关键词 Red/ET同源重组 噬菌体 载体构建
中图分类号:Q392 文献标识码:A
2001年人类基因组测序完成,基因中所含有的大量的功能信息越来越受到人们的关注。为了解这些复杂的人类密码的含义,对单个基因功能的研究也显得越来越重要。然而,对于真核生物基因片段而言,其序列中除了包含具有功能、能表达的外显子之外,还含有很多调控序列等内含子,如此一个基因片段则可能大至几十或上百Kb。虽然现今有相应的载体,如BAC(人工染色体)和PAC(P1人工染色体)能装载如此大量的基因信息,但是要找到这些基因片段的限制性内切位点,并在体外对其进行长距离的片段扩增的难度比较大。
Red/ET重组技术是以传统的同源重组技术为基础,但是,相对于普通的同源重组技术而言,它具有简单、快速、高效等特点,而且整个重组过程不需要经过体外扩增阶段,这样可以避免外界环境引起的不必要的突变。
1传统同源重组
自然发生的同源重组是两个DNA分子之间进行片段交换,从而使序列重排。同源重组分析最早开始于1956年。在大肠杆菌中,A John Clark发现了两种与同源重组有关的酶,RacA和RecBCD。在同源重组发生的时候,RecA结合DNA单链或双链,对双链DNA进攻,把原来配对的互补链分开,并尝试自身与被入侵DNA链配对。当配对成功后,RecA蛋白脱落。或者在RecBCD的引导下,使目标DNA解旋,以便使外源DNA分子插入并在RecA的帮助下进行重组。当两条链发生重组时,会产生holliday中间体,该中间体在重组完成时由RuvAB和RecG蛋白进行拆分。至此,形成两条含有异源序列的双链DNA。
在质粒或噬菌体转入大肠杆菌的实验中,当同源区段小于75bp时会使重组率显著降低。但是实际上,在RecA重组系统中,要求同源臂的长度在1kb左右,且反应条件要求比较苛刻。
2 Red/ET同源重组技术
Red/ET同源重组系统,其实是Red同源重组系统和ET同源重组系统的总称,其中ET系统于1998年由Stewart等在大肠杆菌中诱导的Rac噬菌体中发现,随后又在 噬菌体中发现了Red系统。在这两个系统中,整个重组过程由DNA重组酶参与而不需要限制性内切酶,且对同源臂长度的要求低,僅需35-50bp的同源序列,而传统的RecA同源重组则需要大约1kb的同源序列。并且该系统介导的整个同源重组过程都是在细内进行,避免了因胞外反应引起的不必要突变。
2.1 ET同源重组系统
该系统于1998年由Stewart等在大肠杆菌中诱导的Rac噬菌体中发现,其所需的单侧同源臂长度仅为35-50 bp,该噬菌体通过表达蛋白RecE和RecT来介导重组反应。其中RecE蛋白有5′-3′外切酶活性,能够从5′-3′端依次切下双链DNA上的碱基,使DNA分子形成3′粘性末端。RecT是一种单链结合蛋白,保护单链核苷酸不被降解,能在退火时指导单链入侵。
在ET系统中,带有同源臂的外援DNA分子可以直接通过同源臂找到同源区域,然后进行DNA的互换。
2.2 Red同源重组系统
Red重组系统也是由Stewart的实验小组在 噬菌体中发现,它是由Red 和Red 两种蛋白组成的同源重组系统。Red 由三个蛋白质亚基组成,中间的空隙能消化DNA分子,与ET系统中的RecE蛋白相似,它具有5′-3′端核酸外切酶活性,能形成3′粘性末端。Red 也是一种单链结合蛋白,作用相似于ET系统中的RecT蛋白。
3 重组原理及操作
3.1 Red/ET同源重组的原理
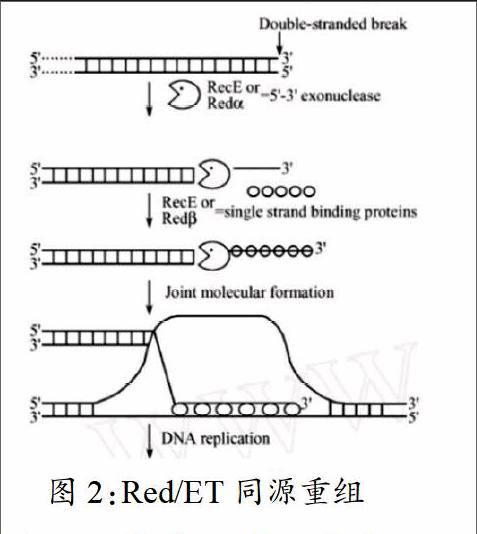
根据Red/ET中Red 及RecE、Red 及RecT的功能,推测了该同源重组系统的作用机理。
当带有同源臂的外源DNA分子进入到大肠杆菌中时,Red 或RecE发挥其5′-3′端核酸外切酶功能,由5′端开始降解DNA序列,产生的3′粘性末端,这时外源DNA具备了与受体DNA发生重组的条件。产生的粘性末端被RecT或者Red 结合35bp的单链核苷酸序列,同时防止被细胞内存在的核酸内切酶降解。在DNA退火时,含有粘性末端的3′单链进攻双链DNA片段,重组。发生重组的两条DNA链经过剪切和DNA聚合酶的修复作用后,重组DNA形成,外源DNA成功导入受体DNA中。
但是在大肠杆菌中有一种RecBCD蛋白,它具有极强的核酸外切酶活性,能将外源线性DNA降解掉,从而降低重组的成功率。所以在RecBCD蛋白缺失的大肠杆菌中Red/ET同源重组的方法才能最大可能的发挥作用。为了能提高外源DNA在大肠杆菌中的重组率,首先应该抑制RecBCD在大肠杆菌中的表达量,为此将 噬菌体Gam蛋白的基因导入到重组系统中。Gam蛋白能与RecBCD形成RecBCD-Gam符合体,抑制RecBCD的核酸外切酶活性。
3.2操作方式
该重组技术在的操作方式主要有两种。(1)建立含有将 基因的ET系统或Red系统的质粒,在实施重组时将三者共同导入受体分子中,共同表达。(2)直接在受体菌中筛选出能进行Red/ET重组的菌株。前者的重组成功率更高,但是当受体分子不容易接受外源DNA时,后一种方式就显得更加便捷。
4系统优化
最初Red/RT系统采用的是pDAB-ET 依托型质粒载体,在该载体中,recE、recT以及 基因分放在PBAD、EM7和Tn-5启动子下共表达。在之后构建的pDAB-ETg和pDAB-gba载体中,RecE/RecT/Red 和Red /Red /Red 被放在PBAD启动子后面共表达,该启动子受L-阿拉伯糖诱导调控。但是在实际应用过程中却发现了一些问题。
(1)在发生重组后该载体不容易从宿主细胞中去除,影响后续的研究工作。为此,研究人员开发了温控性质粒。其原理是将Red /Red 与 基因放在含有PL启动子和cI857基因的载体上,然后导入大肠杆菌中表达。在温度为30℃时,cI857基因启动PL启动子,使Red系统不表达。当温度为42℃时,cI857基因的作用被抑制,Red系统得以重新表达。
(2)由于工程用大肠杆菌一般都缺乏RecA蛋白,在这样的情况下重组的发生率会降低。在最初的目的中使RecA缺失,其目的是让真个表达系统中外源DNA复制更稳定。但是为了避免RecA缺失对重组率的影响又从新引入了RecA蛋白。
(3) 基因在表达的过程中不易控制,它的过度表达会影响RecBCD的活性,进而影响质粒表达的稳定性(Gam蛋白与RecBCD蛋白结合只是抑制了RecBCD对核酸的降解能力,但仍然能对同源重组起促进作用)。为了解决这样的问题,研究者将 基因和Red/ET系统共表达,从而对 基因进行调控。
此外,有研究发现, 基因上的5′端的非翻译区域对重组效率有很大的影响。将含有5′端的非翻译区突变的基因与Red 和Red 连接,可以很大程度的提高重组效率。
5在载体构建中的应用
2006年Osterrieder等運用Red/ET系统和反筛选系统(I-SceI蛋白)设计出了两步重组方法。首先设计一个含有I-SceI同源臂的基因片段,通过Red/ET系统转入载体之中。构建好的重组载体通过诱导I-SceI蛋白的表达,使双链产生切口。切口处可作为同源臂,进行下一次的异源重组。这种两步重组系统大大地提高了重组的精确性。
在对致病菌基因的研究中,朱善元等对禽致病大肠杆菌的ibeA基因进行缺失,用缺失该基因的大肠杆菌粘附与侵袭人微血管内皮细胞。结果显示,缺失ibeA基因的大肠杆菌与微血管内皮细胞的粘附能力显著下降,侵袭率也显著下降。Murphy等还通过该技术对致病大肠杆菌的有毒基因进行敲除,以为免疫制剂的进一步研究提供理论方法。
兰德松等,利用Red/ET同源重组技术,通过两步重组法构建了禽流感A/Goose/Guangdong/3/96(H5N1)毒株的血凝素(HA)基因的重组火鸡疱疹病毒。他们的研究为新型禽流感重载活性疫苗的开发奠定了基础。
6展望
Red/ET同源重组技术的使用,为外源基基因的重组表达提供了一种快捷、简单的方法。在抗生素的开发中,使抗生素异源表达,提高产量也是研究中的重点。该技术除了能方便的运用于抗生素的异源表达方面,还可以通过对基因簇的修饰、重排、删除等来创造新的抗生素。现在已有该方面的研究。
参考文献
[1] 马先勇,姚开泰.同源重组技术研究进展[J].生物工程进展,1996,16(3):16-23.
[2] 王军平, 张友明. Red/ET重组及其在生物医学中的应用[J].生物工程学报,2000,18(12):502-506.
