亚热带和热带森林内小型啮齿动物觅食行为差异*
2017-01-04郎政伟巩合德
郎政伟,巩合德
(1.长宁县桃坪乡人民政府, 四川 长宁644300;2.西南林业大学 生态旅游学院,云南 昆明 650224)
亚热带和热带森林内小型啮齿动物觅食行为差异*
郎政伟1,巩合德2
(1.长宁县桃坪乡人民政府, 四川 长宁644300;2.西南林业大学 生态旅游学院,云南 昆明 650224)
利用哀牢山5种主要依靠啮齿动物扩散的变色锥、硬壳柯、木果柯、顺宁厚叶柯和滇润楠种子作为研究材料,研究哀牢山亚热带中山湿性常绿阔叶林和西双版纳热带雨林林内小型啮齿动物觅食行为的差异。结果显示,西双版纳啮齿动物收获5种试验种子的速度比哀牢山更快(Wald statistic=1 456.397,df=1,P<0.001);西双版纳啮齿动物原地取食和搬运后贮藏种子比例均高于哀牢山;种子被收获率与种子大小在哀牢山呈负相关(R=-0.669,P=0.050),而在西双版纳呈正相关(R=0.048,P=0.006);两地种子原地被取食率与种子大小均呈负相关性,两地原地贮藏率与种子大小均未呈现相关性,两地种子被搬运率与种子大小均呈正相关性,两地种子搬运后被取食率与种子大小均呈负相关性,两地种子搬运后被贮藏率与种子大小均呈负相关性。哀牢山和西双版纳两地种子平均扩散距离没有显著差异,但两地所有被扩散种子个体大小与其扩散距离均呈正相关性(哀牢山,R=0.131,P<0.001;西双版纳,R=0.313,P<0.001)。试验结果表明两地森林内啮齿动物的觅食行为具有相似的选择模型,但西双版纳啮齿动物可能对种子大小具有更明显的选择差异。
种子大小;啮齿动物;觅食行为;哀牢山;西双版纳
当前国内外对啮齿动物觅食的研究主要集中在单一森林内啮齿动物的觅食行为及其影响因素[1~12],或者更多地研究同一森林生态系统不同生境中啮齿动物的觅食行为[13~16],鲜有研究不同森林内啮齿动物觅食行为的差异。因此,本研究选择哀牢山亚热带中山湿性常绿阔叶林和西双版纳热带雨林两个森林生态系统作为研究地点,选取哀牢山5种主要依靠小型啮齿动物扩散种子的壳斗科和樟科树种种子作为研究材料,通过了解两地林内啮齿动物觅食行为差异,进一步认识不同森林内啮齿动物对植物种子的觅食偏好。
1 研究区概况
研究在2014年1月开展。研究地点位于云南省哀牢山国家级自然保护区景东县徐家坝地区,地理位置为24°32′N,102°01′E,海拔2 450m。根据哀牢山森林生态系统定位研究站的长期监测资料,研究区域的平均年降雨量为1 931mm,干季、雨季分明,雨季(5-10月)的降雨量占年降雨量的85%左右。年平均蒸发量为1 485mm,年平均温度为11.3℃,最热月(7月)气温为16.4℃,最冷月(1月)气温为5.4℃,无霜期200天左右。研究地点的亚热带中山湿性常绿阔叶林保护完好,主要由壳斗科(Fagaceae)、茶科(Theaceae)、樟科(Lauraceae)和木兰科(Magnoliaceae)的树种组成。主要植物种类有硬壳柯(Lithocarpushancei)、木果柯(Lithocarpusxylocarpus)、变色锥(Castanopsiswattii)、南洋木荷(Schimanoronhae)等[17]。红颊长吻松鼠(Dremonysrufigenis)、社鼠(Niviventerconfucianus)、安氏白腹鼠(Niviventerandersoni)和中华姬鼠(Apodemusdraco)等是林内主要的啮齿类种子取食者和扩散者[18]。哀牢山试验所在地点为中国科学院哀牢山生态站6hm2样地,该样地的建设遵循美国史密森热带研究所的热带森林研究中心1980年在巴拿马Barro Colorado Island建立的50hm2样地的技术规范[19]。整个样地东西长200m,南北长300m,被划分为150个20m×20m的样方。
对照试验于2014年3月开展。研究地点位于云南省西双版纳州勐腊县勐仑自然保护区,地理位置为21°57′ N,101°12′E,海拔约810m。研究样地内热带季节性雨林和季风常绿阔叶林镶嵌交错分布。在季节性雨林内,群落上层优势种为绒毛番龙眼(Pometiatomentosa)、千果榄仁(Tenninaliamyriocarpa),中层树种有云南玉蕊(Barringtoniamacrostachya)、大叶白颜树(Gironnierasubaequalis)等,下层主要树种有细罗伞(Ardisiatenera)、崖豆树(Millettiateptobotrya)等[20]。季风常绿阔叶林内乔木主要由短刺锥(Castanopsisechidnocarpa)、滇银柴(Aporusayunnanensis)、红花木犀榄(Olearosea)、截头石栎(Lithocarpustruncatus)、红木荷(Schimawallichii)和枹丝锥(C.calathiformis)组成(植物群落数据由西双版纳热带植物园热带雨林生态系统研究站提供)。根据以往的一些研究,西双版纳热带森林中分布的小型啮齿动物有社鼠、针毛鼠(Niviventerfulvescens)、红刺鼠(Maxomyssurifer)、王鼠(Maxomysrajah)、黄胸鼠(Rattusflavipectus)、屋顶鼠(Rattusrattus)、锡金小鼠(Muspahari)、丛林小鼠(Musfamulus)、小泡巨鼠(Leopoldamysedwardsi)、青毛鼠(Berylmysbowersi)、赤腹松鼠(Callosciuruserythroeus)、明纹花鼠(Tamiopsmacclellandi)和隐纹花鼠(Tamiopsswinhoei)等[21~24]。
2 材料与方法
2.1 试验设计
种子成熟季节(2013年10月),于哀牢山试验样地内随机收集成熟、新鲜、完好的变色锥、硬壳柯、木果柯、顺宁厚叶柯(Lithocarpuspachyphyllus)和滇润楠(Machilnsyunnanensis)种子。所有种子带回试验室保存于4℃的冰柜里以备试验使用。试验前对试验种子进行浸水处理,筛选成熟、颗粒饱满、无蛀虫的种子并晾干。对每一颗试验种子,进行单粒称重(精确到0.01g)、钻孔(直径0.6mm),并用1根长约15cm的细铁丝连接1白色塑料标签(3.5cm×2.5cm)将种子编号[25]。利用黑色油性笔对每1个标签进行唯一的字母和数字编号,以便于试验检查。当啮齿动物把带有标签的种子贮藏于浅层土壤中,一部分标签仍暴露于地表以利于种子命运追踪。标签标记对于啮齿动物的种子扩散和贮藏没有显著的影响[25~26]。
在哀牢山试验点样地内每个树种随机选取20棵试验树种作为母树,共100棵母树;西双版纳试验点以间距20m设置5个样带,每条样带上间隔20m选择1棵样地内优势树种作为母树,每条样带10棵母树,共50棵母树。以母树为圆心,在半径0.5m的圆环上等距离设置4个种子释放点,每个释放点依照直径15cm的圆环放置试验种子10粒。西双版纳试验点试验种子按照随机间隔的方法放置,避免同一样带上相邻两棵母树为同种试验种子。哀牢山共释放4 000粒试验种子,每个树种种子800粒;西双版纳共释放2 000粒种子,每个树种种子400粒。释放后种子的命运检查方法参照Wang and Chen的方法并有所改动[26],共检查12次释放地点种子的命运,分别为释放种子后的第1天、2天、3天、4天、6天、8天、12天、16天、20天、28天、36天。种子搜查范围为以种子释放地点为圆心半径为25m的范围。当找到被搬走的种子,记录种子离释放地点的距离(精确到0.01m)和相对于释放地点的角度(精确到5°)。基于目前的研究[9,27],将种子命运作以下分类:(1)原地留存和被收获(即所有被啮齿动物原地取食、原地贮藏和搬离释放点的种子);(2)被收获种子分为原地取食(仅剩余标签和残存的种子碎屑)、原地贮藏和被搬运3类;(3)被搬运的种子分为搬运后取食、搬运后贮藏(包括仅搬运在地表未贮藏种子)和消失(搜索范围内未找到的种子,因此无法判定种子命运)3类。
2.2 啮齿动物调查
试验前一周在研究区域采用捕鼠笼活捕法[28],以了解试验区域主要的小型啮齿动物种类和种群数量规模。采用滇润楠种仁作为诱饵,将种子挂在40cm×14cm×15cm的捕鼠笼内。总共释放49个捕鼠笼,按照每个捕鼠笼间隔10m的距离在7条样带上设置一个7m×7m的捕捉网格。调查时间为7天,每天对捕鼠笼检查两次(08:00和17:00)并查看和重置受损诱饵,对捕捉到的啮齿动物进行编号、拍照并带回试验室暂时饲养,在调查结束后原地释放所有被捕的啮齿动物。西双版纳试验地点受自然保护区条件限制,未使用捕鼠笼活捕法进行啮齿动物调查,而是安装红外相机以记录啮齿动物种类。在试验样地内随机选择6棵母树放置6台红外相机,相机固定在母树离地面40cm处,并对准试验种子释放点,拍摄时间为7天。
2.3 数据处理
利用Cox regression analysis 检验种子大小、树种对释放种子留存比例的影响;Independent sample t-test 用于比较被收获种子和原地剩余种子的大小差异,同时应用于检验搬运种子和原地取食种子、搬运后取食和贮藏种子的大小差异;One-way ANOVA用于检验原地取食、原地贮藏和搬运种子之间的大小差异,同时用于搬运后各树种间种子大小和扩散距离的差异检验;Pearson相关性用于种子命运与其种子大小以及种子大小与其扩散距离的相关性检验。所有数据都在SPSS 18.0软件上进行分析。
3 结果与分析
3.1 主要啮齿动物种类
哀牢山捕捉期白天未捕捉到啮齿动物,夜间共捕捉到7只啮齿动物;其中4只社鼠(57.1%),3只中华姬鼠(42.9%)。西双版纳红外相机拍摄总相片数为384张,拍摄到啮齿动物照片数为108张,占总照片数量的28.1%;在拍摄到啮齿动物照片中鼠类为57只,松鼠5只;西双版纳试验点根据照片辨识鼠类主要为社鼠、针毛鼠和红刺鼠,松鼠类为赤腹松鼠。
3.2 收获和原地留存种子
西双版纳试验点(以下简称西双版纳)5种试验种子被啮齿动物收获的速度明显比哀牢山试验点(以下简称哀牢山)快(Wald statistic=1 456.397,df=1,P<0.001)。试验结束时,哀牢山试验种子原地留存比例高于西双版纳(51.4% VS.7.7%)。试验结果显示,变色锥(Wald statistic=428.038,df=1,P<0.001)、滇润楠(Wald statistic=500.740,df=1,P<0.001)、顺宁厚叶柯(Wald statistic=504.010,df=1,P<0.001)、木果柯(Wald statistic=402.277,df=1,P<0.001)和硬壳柯(Wald statistic=261.252,df=1,P<0.001)5种试验种子在西双版纳被啮齿动物收获的速度均比哀牢山快(图1)。试验结束时,变色锥和滇润楠在西双版纳没有种子原地留存,在哀牢山分别还有10.1%、22.0%的种子原地留存;顺宁厚叶柯在哀牢山原地留存的种子比例(38.6%)高于西双版纳(5.5%);木果柯在哀牢山原地留存的种子比例(88.1%)远高于西双版纳(0.3%);硬壳柯在哀牢山原地留存的种子比例(97.1%)高于西双版纳(32.7%)。
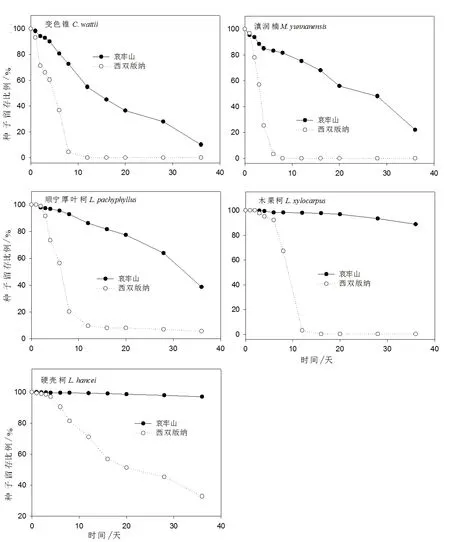
图1 哀牢山和西双版纳各种试验种子在种子释放点的留存比例
Fig.1 Survival curve (proportion)of the tagged seeds after placement at seed releasing plots for each species in Ailao Mountains and Xishuangbanna
试验结果表明,5种试验种子被啮齿动物收获的种子比例与种子大小在哀牢山呈负相关性(R=-0.669,P=0.050),而在西双版纳呈正相关性(R=0.048,P=0.006)(图3)。啮齿动物在哀牢山选择收获个体更大的变色锥种子(Independent-Samplest-test:t=2.310,P=0.023),在西双版纳所有变色锥种子都被收获;滇润楠在哀牢山并未发现被收获种子和原地留存种子之间的大小显著差异性(P>0.05),在西双版纳所有滇润楠种子都被收获;啮齿动物在哀牢山收获个体更大的顺宁厚叶柯种子(t=2.609,P<0.001),但在西双版纳并未发现这种大小选择差异性(P>0.05);啮齿动物在哀牢山偏向于收获个体更小的木果柯种子(t=3.190,P=0.001),而在西双版纳未发现这种大小选择差异性(P>0.05);硬壳柯种子在哀牢山和西双版纳都未发现啮齿动物对收获种子和原地留存种子之间的大小选择差异性(P>0.05)。
3.3 原地取食和搬运种子
所有被啮齿动物收获的种子大部分都被啮齿动物搬运和原地取食,而少量种子被原地贮藏(4.6%,n=1 952,哀牢山,n值为所有被收获种子数;0.4%,n=1 846,西双版纳),由于原地贮藏种子样本量少故未进行数据的统计分析。试验结果显示,哀牢山5种试验种子被啮齿动物原地取食种子比例低于西双版纳(14.9% VS.39.5%);哀牢山原地贮藏种子比例高于西双版纳(2.2% VS.0.4%);哀牢山搬运种子比例低于西双版纳(31.5% VS.52.4%)(图2)。
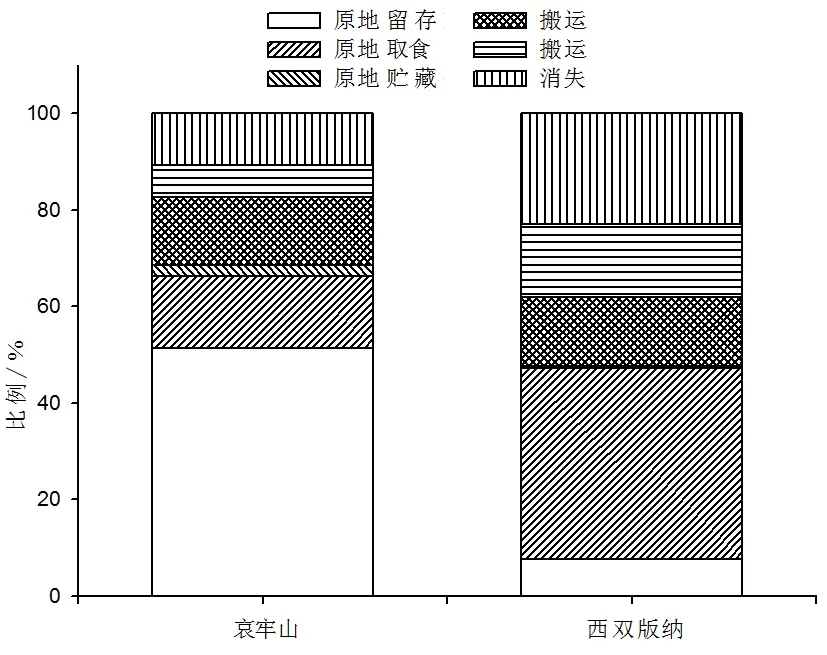
图2 哀牢山和西双版纳种子命运比较
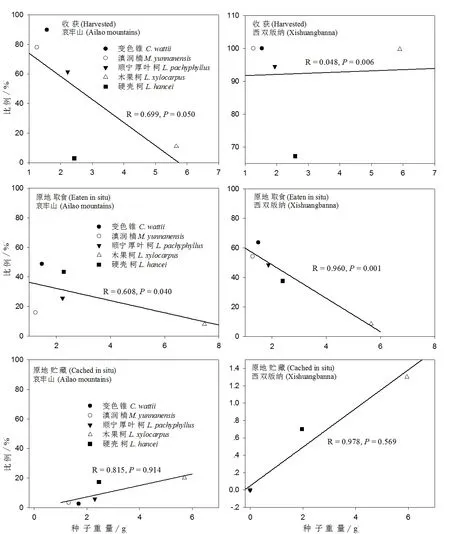
图3 种子大小与种子命运的关系
5种试验种子,被原地取食比例与种子大小在哀牢山(R=0.608,P=0.040)和西双版纳(R=0.960,P=0.001)都呈显著负相关(图3);啮齿动物原地贮藏种子的比例与种子大小在哀牢山和西双版纳两地未呈现显著相关性(P>0.05)(图3);被啮齿动物搬运的种子比例与种子大小在哀牢山(R=0.094,P=0.047)和西双版纳(R=0.964,P=0.012)都呈正相关性(图4)。
3.4 搬运后取食和贮藏种子
5种试验种子,在哀牢山被啮齿动物搬运后取食(14.1% VS.14.4%)、搬运后贮藏(6.6% VS.15.0%)和消失(10.8% VS.23.0%)的种子比例均低于西双版纳(图2)。种子被搬离释放点后,啮齿动物对种子只有取食和贮藏两种策略,但是试验中有一部分种子在搜查中未找到(定义为消失的种子),这类种子因为其不确定的种子命运所以未加入到数据的统计分析中。5种试验种子,被啮齿动物搬运后取食的种子比例与其种子大小在哀牢山(R=0.775,P=0.017)和西双版纳(R=0.337,P=0.026)都呈负相关性(图4);被啮齿动物搬运后贮藏的种子比例与其种子大小在哀牢山(P>0.05)未呈现显著相关性,在西双版纳(R=0.272,P=0.021)呈显著负相关性(图4)。
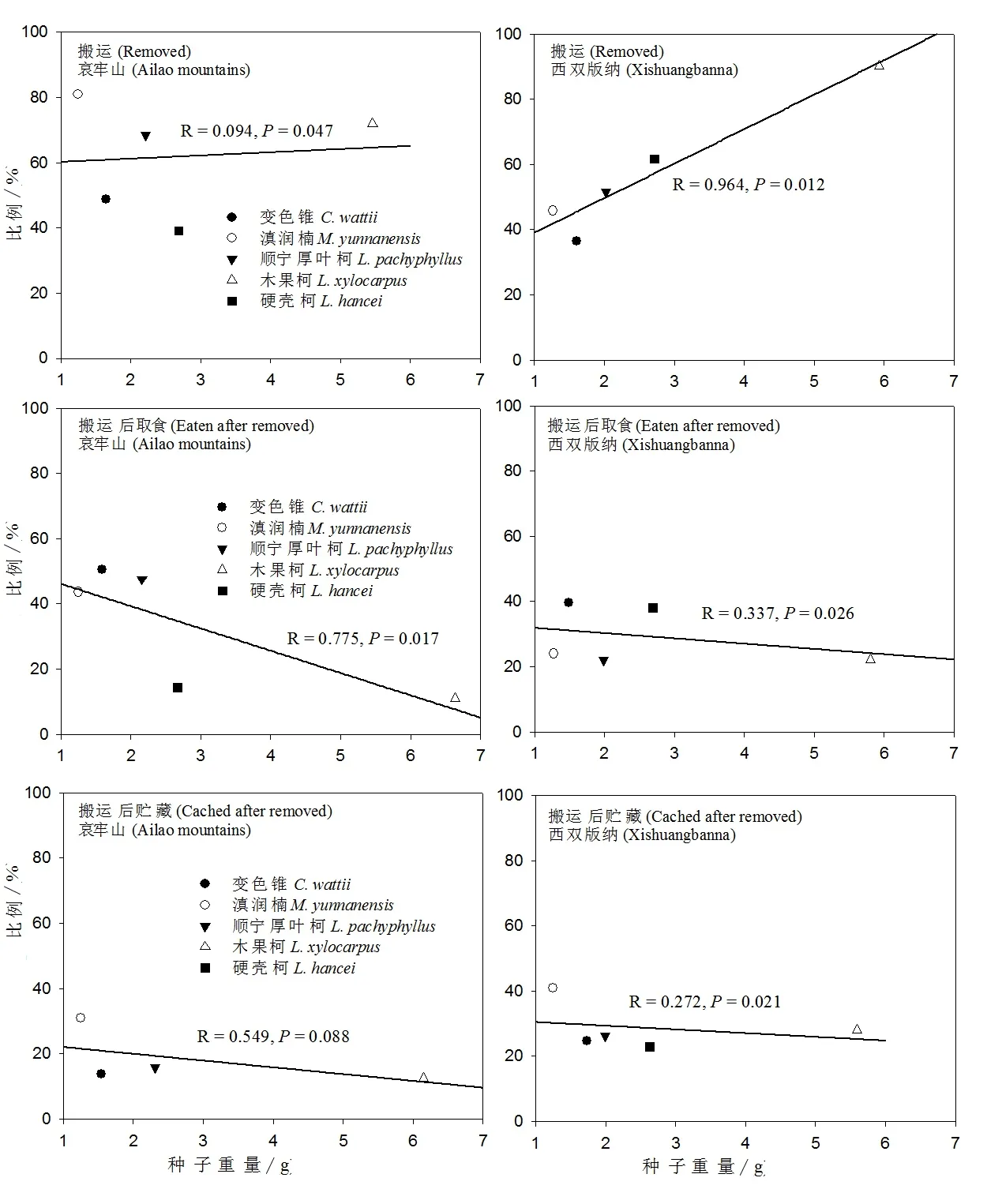
图4 种子大小与种子命运的关系
3.5 扩散距离
5种试验种子在哀牢山的平均扩散距离(5.46±0.41m)大于西双版纳(2.90±0.35m)(P<0.001)。哀牢山5类种子的扩散距离没有显著的差异性(F3 807=2.449,P=0.062;硬壳柯因为只有一颗种子具有扩散距离,因此未进行数据统计分析)。这些种子的平均扩散距离为木果柯(6.02±1.49m,mean±SE,n=14)、硬壳柯(7.83m,n=1)、顺宁厚叶柯(4.74±0.27m,n=209)、变色锥(4.48±0.20m,n=216)、滇润楠(4.10±0.19m,n=372)(图5)。西双版纳木果柯种子(7.31±0.51m,mean±SE,n=181)的平均扩散距离显著大于顺宁厚叶柯(4.16±0.37m,n=94)、滇润楠(3.87±0.28m,n=119)、变色锥(3.60±0.32m,n=94)和硬壳柯(2.22±0.15m,n=101)(F4 584=24.740,P<0.001)(图5)。
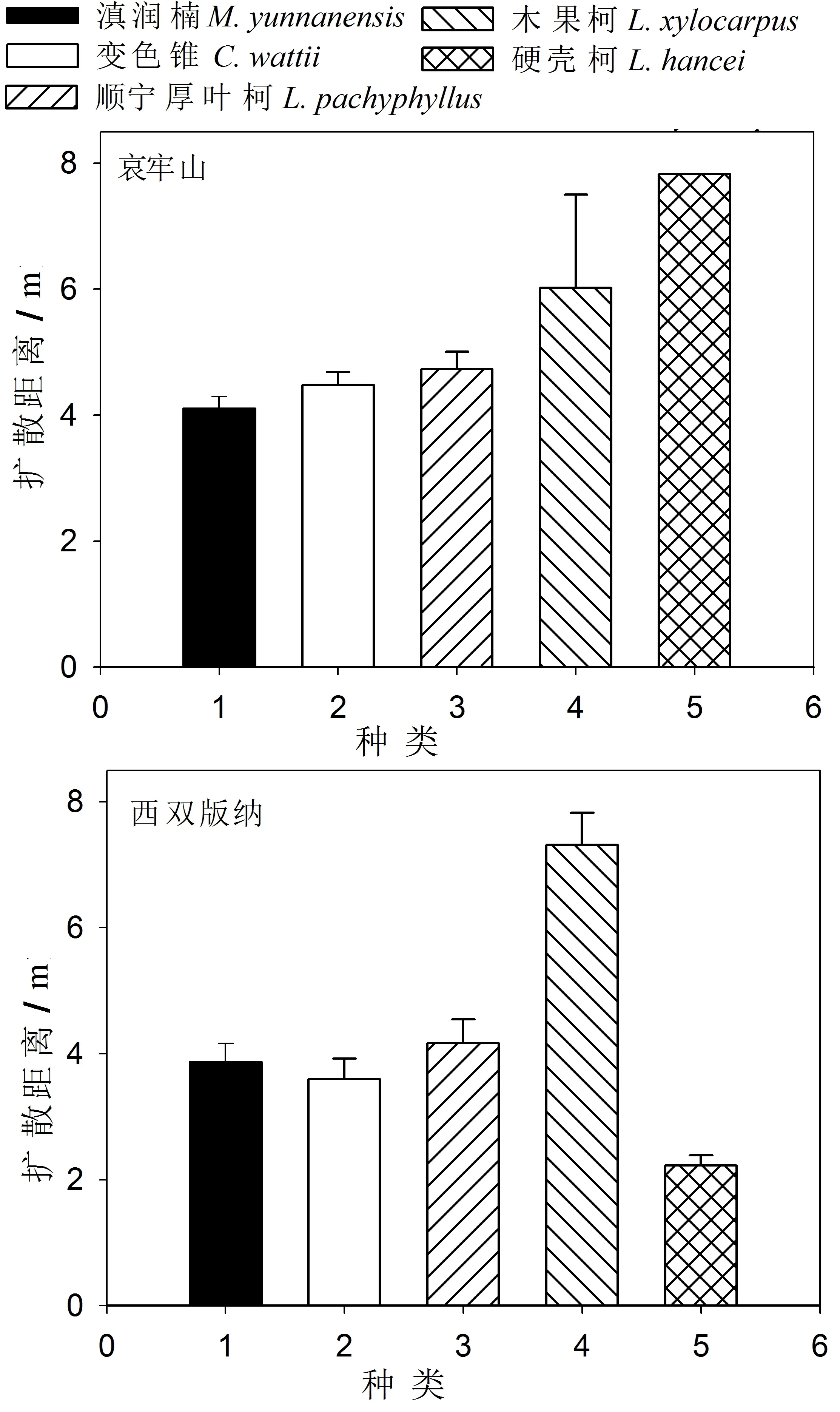
图5 搬运种子扩散距离
试验结果显示,变色锥的种子大小与其扩散距离在哀牢山(Pearson’s correlation:R=0.177,P<0.001)和西双版纳(R=0.048,P=0.007)都呈现正相关性;滇润楠的种子大小与其扩散距离在哀牢山呈显著正相关(R=0.125,P=0.015),但在西双版纳两者并未呈现明显相关性(P>0.05);顺宁厚叶柯的种子大小与其扩散距离在西双版纳呈显著正相关(R=0.023,P=0.007),但在哀牢山两者并未呈现明显相关性(P>0.05);木果柯的种子大小与其扩散距离在西双版纳呈显著正相关(R=0.101,P<0.001),但在哀牢山两者并未呈现明显相关性(P>0.05);硬壳柯的种子大小与其扩散距离在西双版纳呈显著正相关(R=0.005,P<0.001),但在哀牢山两者并未呈现明显相关性(P>0.05)。通过整合哀牢山和西双版纳两地5种试验种子被搬运种子的数据,哀牢山(R=0.131,P<0.001)和西双版纳(R=0.313,P<0.001)被搬运种子大小与其扩散距离都呈显著正相关性。
4 结论与讨论
由研究结果可知,西双版纳试验点5种植物种子被啮齿动物收获的速度显著比哀牢山快,两地啮齿动物对种子的觅食选择都存在明显的种间差异,而这种差异体现在对不同大小种子的选择上。森林内啮齿动物发现植物种子后,一般面临多项抉择:第一,是取食、贮藏或忽视这些种子;第二,如果取食,是原地取食还是搬运到安全地方取食;第三,如果贮藏,是原地贮藏还是搬运到其他地方贮藏,以备后期取食[12,27]。本试验中,两地林内啮齿动物对小种子植物种类的变色锥、顺宁厚叶柯和硬壳柯倾向于选择收获、搬运个体较大的种子,最终更多地贮藏这些种子。对种子个体大小差异较小的滇润楠,两地啮齿动物都未表现出明显的大小选择差异性。许多研究也证实啮齿动物觅食过程中种子大小和觅食选择之间这种相似的相关性[3~4,26,29]。然而,对于最大种子个体种类的木果柯,两地啮齿动物都选择收获和搬运个体较小的种子。这一矛盾的结果说明,两地啮齿动物在面临种子大小(代表能量含量)和种子处理时间两个问题时,都选择一个有利于最大净能量回报的权衡。这表明哀牢山和西双版纳两地林内啮齿动物在植物种子大小选择上存在相似的模式。
一些研究表明,啮齿动物的个体大小会对啮齿动物对不同大小种子的选择产生影响[30~31],而且不同啮齿动物对植物种子的处理能力也存在差异。Hodgkison等的研究表明不同地点啮齿动物对同类种子(果实)的觅食选择存在显著差异,而且这种发展的觅食行为不受地理起源的影响[32]。哀牢山和西双版纳两地森林内虽然存在共同分布的啮齿动物种类(如社鼠),但仍存在较大的种类差异。本试验结果显示,两地虽然存在啮齿动物的种类和个体大小差异,但两地啮齿动物在种子大小选择上存在共同的觅食选择偏好。
试验结果表明,西双版纳试验点的变色锥、顺宁厚叶柯、木果柯和硬壳柯4种植物种子大小与其扩散距离都呈现显著的相关性,而哀牢山试验点只有变色锥和滇润楠两种种子呈现这种相关性。国内外一些研究的结果也证实种子大小与其扩散距离存在正相关性[4~5,26,33]。但是,Theimer在研究Beilschmiediabancroftii种子时并未发现种子大小和扩散距离之间的相关性[34]。Wang等在研究中指出,种子大小与其扩散距离的这种相关性可能仅存在于一定的种子大小阀值之中,超出这一范围啮齿动物会因搬运更大种子增加了能量消耗而减少自己的净能量回报[26]。本试验中,西双版纳试验点仅有滇润楠种子未呈现种子大小与其扩散距离的相关性,同时大种子种类的木果柯扩散距离大于其他4种种子,说明西双版纳林内啮齿动物对植物种子可能具有更加明显的大小选择差异。此外,除了种子大小,其他的种子性状(如营养物、单宁含量等)也会影响啮齿动物对种子的扩散距离[6,26],这也可能是本研究中其他种类的种子没有发现种子大小与其扩散距离呈现相关性的原因。
在研究地点内,啮齿动物是许多种子植物主要的种子扩散者,啮齿动物在植物种群扩展和森林更新中发挥了重要作用。但是,本试验中并未考虑到其他种子性状的潜在影响,也没有将西双版纳主要依靠啮齿动物扩散种子的树种加入到对比试验中,同时两地不同的森林植被类型、气候条件差异和季节食物丰富度也可能对试验结果产生一定影响。因此,需要更进一步的研究来了解这一对比差异和多样种子性状对啮齿动物觅食行为差异的影响。
致谢:感谢中国科学院西双版纳热带植物园王博副研究员对本文提出的宝贵意见。感谢普洱学院陈泰浩和谭爱飞同学在野外试验和试验室工作中的辛勤工作,感谢中国科学院哀牢山生态站对本研究野外试验工作的大力支持和帮助。
[1]Vander Wall S B.Food hoarding in animals[M].Chicago:University of Chicago Press,1990:1-43.
[2]Brewer S W.Predation and dispersal of large and small seeds of a tropical palm[J].Oikos,2001,92:245-255.
[3]Jansen P A,Bongers F,Hemerik L.Seed mass and mast seeding enhance dispersal by a neotropical scatter-hoarding rodent[J].Ecological Monographs,2004,74:569-589.
[4]Xiao Z S,Zhang Z B ,Wang Y S.Effects of seed size on dispersal distance in five rodent-dispersed fagaceous species[J].Acta Oecologica-International Journal of Ecology,2005,28(3):221-229.
[5]Moore J E,McEuen A B,Swihart R K,etal.Determinants of seed removal distance by scatter-hoarding rodents in deciduous forests[J].Ecology,2007,88:2529-2540.
[6]Wang B,Chen J.Seed size,more than nutrient or tannin content,affects seed caching behavior of a common genus of Old World rodents[J].Ecology,2009,90(11):3023-3032.
[7]Lei J J,Shen K,Yi X F.Pericarp thickness and seed size determine acorn dispersal of five rodent-dispersed oak species[J].Acta Theriologica Sinica,2012,32(2):83-89.
[8]Rusch U D,Midgley J J,Anderson B.Rodent consumption and caching behavior selects for specific seed traits[J].South African Journal of Botany,2013,84:83-87.
[9]Wang B,Yang X L.Teasing apart the effects of seed size and energy content on rodent scatter-hoarding behavior[J].PLoS One,2014,9(10):e111389.
[10]Gong H D,Tang C Z,Wang B.Post-dispersal seed predation and its relations with seed traits:a thirty-species-comparative study[J].Plant Species Biology,2015,30(3):193-201.
[11]陈帆,陈进.华山松种子大小对啮齿动物贮藏行为的影响及其时空变化[J].动物学研究,2011,32(4):435-441.
[12]常罡,王开锋,王智.秦岭森林鼠类对华山松种子捕食及其扩散的影响[J].生态学报,2012,35(10):3177-3181.
[13]Medellin R A,Gaona O.Seed dispersal by bats and birds in forest and disturbed habitats of chiapas,Mexico[J].Blotroplca,1999,31(3):478-485.
[14]闫兴富,周立彪,刘建利.啮齿动物捕食压力下生境类型和覆盖处理对辽东栎种子命运的影响[J].生态学报,2012,32( 9):2778-2787.
[15]师一粟.片段化生境中苦槠种子的扩散格局[D].上海:华东师范大学资源与环境科学学院,2012.
[16]赵雪峰,路纪琪,乔王铁,等.生境类型对啮齿动物扩散和贮藏栓皮栎坚果的影响[J].兽类学报,2009,29(2):160-166.
[17]邱学忠.哀牢山生态系统研究[M].昆明:云南科技出版社,1998:12-27.
[18]Xiao Z S,Zhang Z B.Behavioural responses to acorn germination by tree squirrels in an old forest where white oaks have long been extirpated[J].Animal Behaviour,2012,83:945-951.
[19]Condit R.Tropical forest census plots:methods and results from Barro Colorado Island,Panama and comparison with other plot[M].Berlin:Springer,1998.
[20]Zhu H.Forest vegetation of Xishuangbanna,south China[J].Forestry Studies in China,2006(8):1-58.
[21]Wu D L,Luo J,Fox B J.A comparison of ground-dwelling small mammal communities in primary and secondary tropical rainforests in China[J].Journal of Tropical Ecology,1996,12:215-230.
[22]陈志平,王应祥,冯庆,等.云南西双版纳片断热带雨林鼠形啮齿类的物种多样性研究[J].动物学研究,1996,17:451-445.
[23]吴德林,奉勇.西双版纳热带雨林地栖小兽群落的初步研究[J].兽类学报,1997,17:227-231.
[24]孟令曾,高秀霞.西双版纳不同地点和生境鼠类及蚂蚁的多样性比较[J].生态学杂志,2007,26:802-809.
[25]Xiao Z S,Jansen P A,Zhang Z B.Using seed-tagging methods for assessing post-dispersal seed fate in rodent-dispersed trees[J].Forest Ecology and Management,2006,223:18-23.
[26]Wang B,Wang G,Chen J.Scatter-hoarding rodents use different foraging strategies for seeds from different plant species[J].Plant Ecology,2012,213:1329-1336.
[27]Wang B,Ye C X,Cannon C H,etal.Dissecting the decision making process of scatter-hoarding rodents[J].Oikos,2013,122:1027-1034.
[28]肖治术,张知彬.啮齿动物的贮藏行为与植物种子的扩散[J].兽类学报,2004,24(1):61-70.
[29]Forget P M,Milleron T,Feer F.Patterns in post-dispersal seed removal by neotropical rodents and seed fate in relation to seed size.In:Newbery,D.M.,Prins,H.T.,Brown,N.D.(Eds.),Dynamics of Tropical Communities[M].Blackwell,1998:25-49.
[30]Munoz A,Bonal R.Are you strong enough to carry that seed? Seed size/body size ratios influence seed choices by rodents[J].Animal Behaviour,2008,76:709-715.
[31]Tamura N,Hayashi F.Geographic variation in walnut seed size correlates with hoarding behaviour of two rodent species[J].Ecological Research,2008,23:607-614.
[32]Hodgkison R,Ayasse M,Haeberlein C,etal.Fruit bats and bat fruits:the evolution of fruit scent in relation to the foraging behaviour of bats in the New and Old World tropics[J].Functional Ecology,2013,27(4):1075-1084.
[33]Vander Wall S B.The effects of seed value on the caching behavior of yellow pine chipmunks[J].Oikos,1995,74:533-537.
[34]Theimer T C.Intraspecific variation in seed size affects scatter-hoarding behaviour of an Australian tropical rain-forest rodent[J].Journal of Tropical Ecology,2003,19:95-98.
Foraging Behavior of Scatter-hoarding Rodent in Subtropical and Tropical Forests
LANG Zheng-wei1,GONG He-de2
(1.The people’s government of Taoping township,Changning county,Changning Sichuan 644300,P.R.China;2.Ecotourism Faculty,Southwest Forestry University,Kunming Yunnan 650224,P.R.China)
In this study,we surveyed rodent seed predation and dispersal of five common tree species (Castanopsiswattii,Lithocarpushancei,Lithocarpusxylocarpus,Lithocarpuspachyphyllus,Machilnsyunnanensis)in a natural subtropical forest located in Ailao Mountain subtropical evergreen broad-leaved forest and Xishuangbanna National Nature Reserve tropical rainforest.Our results showed that the speed at which rodents harvested seeds in Xishuangbanna were faster than that of Ailao Mountain(Wald statistic=1 456.397,df=1,P<0.001).The proportion of seeds that were eaten in situ and cached after removal in Xishuangbanna was higher than those in the Ailao Mountains.The rate of harvested seeds and seed size exhibited a negative correlation in Ailao Mountain(R=-0.669,P=0.050),while a positive correlation was found in Xishuangbanna(R=0.048,P=0.006).In both sites,there was a negative correlation between the proportion of seeds that were eaten in situ and seed size,but there was no significant correlation between the proportion of seeds that were cached in situ and seed size.In both locations,there was a positive correlation between the proportion of removed seeds and seed size,while the proportion of seeds that were eaten and cached after removal exhibited a negative correlation with seed size.No significant differences were found in seed dispersal distance in both locations.By pooling all of the removed seed observations within each site,positive relationships between seed size and dispersal distance were found in both the Ailao Mountains (R=0.131,P<0.001)and in Xishuangbanna (R=0.313,P<0.001).Our results showed that rodents forage similarly at both sites,but rodents in Xishuangbanna may have significant selection processes based on seed size.
seed size;rodents;foraging behavior;Ailao Mountains;Xishuangbanna
10.16473/j.cnki.xblykx1972.2016.06.024
2015-10-13
国家自然科学基金委 ·国家自然科学基金项目(31470494, 31100315)。
郎政伟(1989-),男,硕士研究生,从事生物地理学方向研究。E-mail:langzw1989@163.com
简介:巩合德(1978-),男,副教授,博士,从事森林生态学方向研究。E-mail:472912369@qq.com
Q 959.837
A
1672-8246(2016)06-0133-09
