牛筋草对百草枯抗性水平初探
2016-12-19沈雪峰李颖慧韩承畴
沈雪峰,胡 芳,陈 勇,李颖慧,韩承畴
(华南农业大学杂草研究室,广东 广州 510642)
牛筋草对百草枯抗性水平初探
沈雪峰,胡 芳,陈 勇*,李颖慧,韩承畴
(华南农业大学杂草研究室,广东 广州 510642)
为探讨牛筋草Eleusineindica(L.) Gaertn对百草枯的抗性水平,测定了百草枯对抗性和敏感生物型牛筋草的抗性水平、叶绿素荧光参数和抗性基因的扩增。结果表明,百草枯对抗性和敏感生物型牛筋草地上部鲜重抑制中浓度(ED50)分别为0.1029和6.1120 kg/hm2,抗性和敏感生物型ED50的比值为59.48;经1 kg/hm2百草枯处理牛筋草3 h后,抗性生物型叶绿素荧光参数逐渐恢复,敏感生物型逐渐降低,而0.1 μmol/L硝酸盐+1 kg/hm2百草枯处理的抗性生物型未出现恢复现象。推测转运体参与了百草枯在抗性生物型牛筋草体内的转运。经特异性引物,抗性生物型获得扩增片段1021 bp,而敏感生物型未能获得。经BLAST比对,该EST片段的核苷酸序列与高粱假设蛋白的同源性为96 %,该蛋白序列来源于氨基酸阳离子转运体,与玉米的氨基酸阳离子转运体基因高度相似片段为96 %。
牛筋草;百草枯;抗性水平;转运体;叶绿素荧光
牛筋草(EleusineindicaL.)属禾本科属1年生草本植物,多生于较湿润的农田、果园或路旁,广布世界各地,已被列为世界十大恶性杂草之一,危害非常严重[1-2]。百草枯(1,1-二甲基-4,4-二氯联吡啶化合物,paraquat)属于典型的光合系统I抑制剂类除草剂[3],已被广泛用于防除田间、果园和荒地等一年生或多年生杂草,目前我国已成为全球百草枯生产和使用的第一大国[4]。自1980年在台湾省发现第一例抗百草枯的苏门白酒草(Conyzasumatrensis)以来[5],世界范围内抗百草枯的杂草生物型不断出现[6]。经调查发现[2],我国华南地区许多长期使用百草枯的果园,百草枯对牛筋草的防除效果已经很差,给当地的果园管理带来极大不便。2010年,在马来西亚连续使用多年的百草枯对果园牛筋草的防除效果很不理想,进一步的研究发现其GR50是敏感性的3.6倍[1]。百草枯的抗性机制十分复杂,包括抑制转运、隔离到液胞、增强氧自由基清除酶的活性等。Szigeti[7]在拟南芥基因组中分离到一个类似大肠杆菌PotE转运体的同源基因AtPotE,能被百草枯诱导,其表达丰度在抗性植株中较敏感植物中的高,可能定位在液泡中。Jori et al.[8]等通过比较百草枯处理后抗性和敏感生物型加拿大飞蓬(ConyzacanadensisL. Cronq)mRNA表达的差异,发现两条仅在抗性生物型中表达的EST片段,分别与EmrE,氨基端阳离子转运体(CAT4)的氨基酸同源性在98 %,表明转运体可能参与了细胞内百草枯的运输。目前国内尚未见有关百草枯抗性生物型牛筋草的研究报道。本研究旨在探索牛筋草对百草枯的抗药性水平,明确抗药性生物型和敏感生物型牛筋草受百草枯和转运体抑制剂处理后,植株内部叶绿素荧光参数的变化,通过转运体特异性引物扩增,研究抗性和敏感牛筋草中转运体的基因表达,从而为延缓抗药性杂草的发展、治理抗性杂草提供理论依据。
1 材料与方法
1.1 供试材料
1.1.1 药剂 42 %的百草枯(paraquat)母液,湖北仙隆化工股份有限公司。
1.1.2 杂草种子 牛筋草(EleusineindicaL.)抗药性生物型(R)和敏感生物型(S),分别于2006年5月采自广州市番禺区大岗镇柑橘园(该区百草枯已连续使用20年)和华南农业大学农场荒地(从未使用过除草剂)。
1.1.3 主要仪器 3WP-2000型行走式喷雾塔,农业部南京农业机械化研究所研制生产;PAM-2500叶绿素荧光仪,德国WALZ公司生产。
1.2 试验方法
1.2.1 不同牛筋草种群对百草枯敏感性水平测定 采用温室盆栽法培养。将去壳的种子播种于盛有营养土的花盆(d=9 cm)中,每盆10粒,10次重复,置于30±2 ℃,光照强度为800 μE·m-2s-1,相对湿度为80 %的温室中培养,定期浇水。4次重复。
待幼苗长至6~7叶期,采用生测喷雾塔进行茎叶喷雾,测定R和S型牛筋草对百草枯的敏感性。喷头孔径0.3 mm,喷雾压力0.3 MPa,雾滴直径100 μm,喷头流量90 mL·min-1。喷施剂量依次为0、0.075、0.15、0.3、0.6、1.2、2.4、4.8 kg a.i. hm-2。以清水为对照,4次重复。喷药7 d后,统计死亡率并称取地上部鲜重。数据采用SPSS 18.0软件进行处理,以植株抑制率的几率值(Y)和百草枯浓度的对数值(x)建立回归方程,计算ED50。
1.2.2 抗性和敏感牛筋草叶绿素荧光参数的测定 幼苗培养方式同1.2.1节。待植株长至分蘖期随机分成2组,其中一组喷药浓度为1 kg a.i. hm-2;另一组经转运体抑制剂(0.1 μmol·L-1KNO3)处理1 h后,也同样喷施1 kg a.i. hm-2的百草枯,之后每隔1 h使用0.1 μmol·L-1的KNO3处理1次。分别于喷施百草枯后0、1、2、3、4、5、6、12、24和48 h分别测定R和S型牛筋草叶片叶绿素荧光参数[9]。各测定10株,每植株测10个叶片,每隔1 h记录PSII原初光能转化效率(Fv/Fm)值,重复5次。
1.2.3 抗性和敏感牛筋草抗性基因扩增 利用Multalin软件,对水稻、小麦、大麦、玉米等多种植物品系的20条CAT4核苷酸序列进行比对,根据保守区域进行兼并引物设计引物为F(5’-AAGGKRTTGCCTGGTTGA-3’),R(5’-GGAGGKACATATCTRAG TA-3’)。PCR总反应体系为50 μl,其中,包括DNA模板2 μl,5×PCR Buffer 5 μl,下游特异性PCR引物2 μl,上游特异性PCR引物2 μl,TaKaRa ExTaq®0.5 μl,Mg2+3 μl,ddH2O 35.5 μl。PCR反应程序:95 ℃预变性5 min;95 ℃变性30 s;55 ℃退火30 s;72 ℃延伸1.5 min,33个循环;72 ℃后延伸7 min。利用含有EB的1 %的琼脂糖凝胶检测扩增结果,Gel Doc 2000凝胶成像系统观察。
2 结果分析
2.1 抗性和敏感生物型牛筋草对百草枯的敏感性差异
由表1可知,百草枯对牛筋草R、S生物型存活率的抑制中浓度ED50值分别为6.1120和0.1029 kg/hm2,抗性与敏感生物型的ED50比值为59.48。该结果表明牛筋草抗性生物型对百草枯的敏感性明显降低,抗药程度高。
2.2 抗性和敏感生物型牛筋草叶片荧光参数变化
由图1 a可知,经1 kg/hm2百草枯茎叶处理后,R生物型牛筋草的Fv/Fm值呈单峰曲线,峰谷出现在药后3 h,随后呈现上升,趋于平稳;而S生物型牛筋草的Fv/Fm值呈下降趋势,且在百草枯处理48 h后,Fv/Fm值急剧下降至0。经硝酸盐+百草枯处理后,R和S生物型牛筋的Fv/Fm值表现相似,均呈现下降趋势(图1 b),说明硝酸盐抑制了R生物型牛筋草活性。
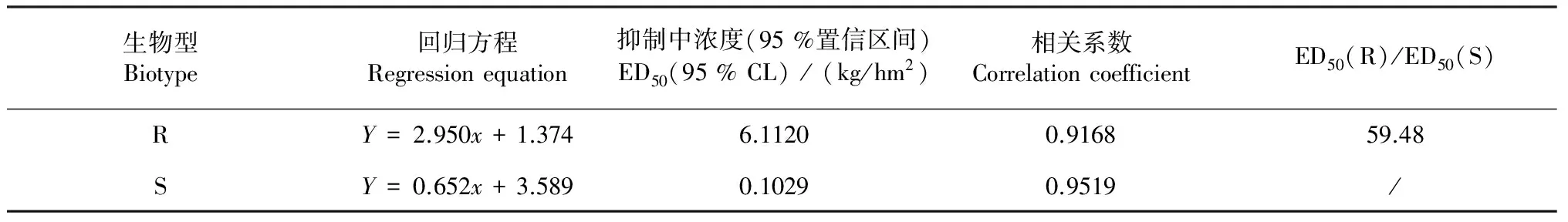
表1 牛筋草抗性和敏感生物型对百草枯的敏感性差异
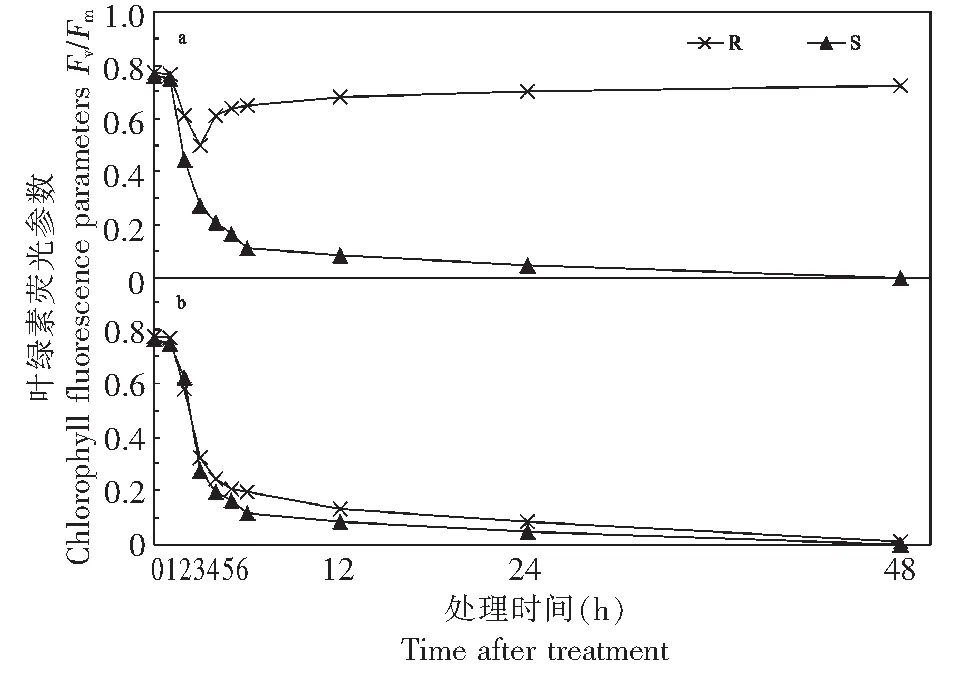
a:1 kg/hm2百草枯处理R和S生物型牛筋草;b:1 kg/hm2百草枯和0.1 μmol/L硝酸盐处理R和S生物型牛筋草a: Resistant and susceptible biotypes of E. indica treated by 1 kg/hm2 paraquat; b: Resistant and susceptible biotypes of E. indica treated by 0.1 μmol/L nitrates and 1 kg/hm2 paraquat图1 百草枯和硝酸盐对R和S生物型牛筋草叶片Fv/Fm值的影响Fig.1 The change trend of Fv/Fm value in resistant/susceptible E. indica after paraquat and nitrates treatment
2.3 抗性和敏感生物型牛筋草总DNA扩增与序列分析
通过保守区域设计兼并引物,分别对R和S生物型牛筋草总DNA进行扩增,结果从R生物型牛筋草中扩增到1条1021 bp的核苷酸片段,在S生物型牛筋草中未得到该片段(图2)。经BLAST比对,显示与高粱假设蛋白(Sorghumbicolorhypothetical protein)同源性为96 %,与玉米LOC100285193基因(ZeamaysLOC100285193),全长cDNA克隆玉米mRNA(Zeamaysfull-length cDNA clone ZM_BFb0068I04 mRNA)同源性均为95 %,与短柄草LOC100846653基因(BrachypodiumdistachyonLOC100846653)同源性为94 %,而上述序列均与转运体有关。同时,对PCR产物测序结果进行序列分析,结果表明,该序列部分片段与玉米、二穗短柄草、拟南芥酸阳离子转运体等基因片段同源。
3 结论与讨论
百草枯的主要作用部位是植物叶绿体光系统PSⅠ的光合膜系统,其作用机理是与光系统PSⅠ中的电子受体竞争,捕获电子传递链中的电子,抑制正常的电子传递,同时产生过氧化物和自由基,破坏质膜、蛋白质、核酸等结构而造成植物死亡[3, 10]。本研究结果证实,取自于华南农业大学农场荒地的S生物型及R生物型牛筋草的ED50值分别为0.1029和6.1120 kg/hm2已经远高于百草枯的推荐剂量0.600 kg/hm2;与S生物型牛筋草相比,R生物型牛筋草的相对抗性水平达到59.48倍。这与Seng et al.[1]对马来西亚R生物型牛筋草研究的结果相似。

(S1~S4: S生物型牛筋草,M: DL2000 DNA Marker,R1~R4: R生物型牛筋草)S1-S4: Susceptible biotypes, M: DL2000 DNA Marker, R1-R4: Resistant biotypes图2 抗性和敏感生物型牛筋草DNA扩增产物Fig.2 DNA-blot analysis of paraquat-sensitive and -resistant E.indica individuals
近年来,Fv/Fm反映了PSⅡ反应中心的原初光能转换效率[9],被用来研究植物对逆境响应,是光合作用光抑制的显著特征,作为判断是否发生光抑制的标准[11]。在胁迫条件下,Fv/Fm值会显著降低[12]。本研究发现,随着百草枯处理时间的延长,S生物型牛筋草PSⅡ活性中心受损逐渐加重,Fv/Fm呈快速下降趋势,说明百草枯对S生物型牛筋草产生了不可逆的胁迫作用,植株受到严重的光抑制,光合系统严重受损,光合作用受阻,敏感牛筋草最终死亡,而R生物型牛筋草经百草枯处理后,Fv/Fm呈现单峰曲线,表现为先降低后升高,之后趋于平稳,说明其光合系统受到影响较小,对光能的利用效率始终保持稳定,叶片PSⅡ的潜在活性和原初光能转化效率都很高,进一步说明抗性牛筋草PSⅡ活性中心未受损,植株能够正常地进行电子传递及光合作用,说明叶绿体机能复活,而叶绿体机能活性恢复过程是由于百草枯被转运到代谢不活跃的细胞区间造成的[10, 13]。
前人[14]研究报道:用真核生物蛋白活性抑制剂放线菌酮(Cycloheximide)和百草枯同时处理加拿大莴苣百草枯抗性植株被破坏;而单独使用放线菌酮并不能使植物致死。表明百草枯能够进入植株叶绿体中,并进行瞬时的功能抑制,推测存在某类其活性能被放线菌酮抑制的百草枯诱导蛋白,其功能是携带百草枯并将其隔离而解毒。本研究发现,经0.1 μmol/L硝酸盐+1 kg/hm2百草枯处理后,R和S生物型牛筋的Fv/Fm值均呈现下降趋势,说明硝酸盐抑制了R生物型牛筋草活性。这表明硝酸钾抑制了位于质膜上ATPases的作用,使转运体失去了来自质膜两侧的质子浓度梯度提供的能量,无法完成百草枯的转运过程,使抗性牛筋草失去功能活性恢复过程,表现出与敏感牛筋草相同的现象。
Su et al.[15]等在百草枯抗性植物中还检测到一个与氨基端阳离子转运体CAT4同源的EST片段,它与定位于膜的H+-ATP酶亚基同源。选择性转运体抑制/恢复试验发现,拟南芥氨基酸转运CAT4,参与百草枯和其它毒性化合物的胁迫应答[16],使用不同的转运体抑制剂处理百草枯抗性莴苣也观察到了同样的现象[17]。本研究选用特异性引物分别对R和S生物型牛筋草进行目的片段扩增,在R生物型牛筋草中扩增得到氨基端阳离子转运体同源序列片段,且其EST片段的核苷酸序列与高粱假设蛋白同源性为96 %,该蛋白序列来源于氨基酸阳离子转运体,与玉米的氨基酸阳离子转运体基因高度相似片段的相似性为96 %。
[1]Seng C T, Lun L V, San C T, et al. Initial report of glufosinate and paraquat multiple resistance that evolved in a biotype of goosegrass (Eleusineindica) in Malaysia [J]. Weed Biology and Management, 2010, 10: 229-233.
[2]AN J, SHEN XF, MA QB, et al. Transcriptome profiling to discover putative genes associated with paraquat resistance in goosegrass (EleusineindicaL.) [J]. PLoS ONE, 2014, 9(6): e99940.
[3]李颖慧, 陈 勇. 杂草对百草枯的抗药性机制研究进展[J]. 生态学杂志,2012, 31(1): 194-199.
[4]Bromilow R H. Paraquat and sustainable agriculture [J]. Pest Management Science, 2004, 60(4): 340-349.
[5]张朝贤, 倪汉文, 魏守辉, 等. 杂草抗药性研究进展 [J]. 中国农业科学,2009, 42(4): 1274-1289.
[6]安 静, 沈雪峰, 陈 勇. 多胺及其转运体在杂草对百草枯抗性机制中的作用研究进展 [J]. 农药学学报, 2013, 15(4): 363-371.
[7]Szigeti Z. Mechanism of paraquat resistance-from the antioxidant enzymes to the transporters [J]. Acta Biologica Szegediensis, 2005,49(1-2): 177-179.
[8]Jóri B, Soós V, Szeg D, et al. Role of transporters in paraquat resistance of horseweedConyzacanadensis(L.) Cronq [J]. Pesticide Biochemistry and Physiology, 2007, 88(1): 57-65.
[9]杨彩宏, 田兴山, 冯 莉, 等. 牛筋草对草甘膦的抗药性 [J]. 中国农业科学, 2012, 45(10): 2093-2098.
[10]倪万潮, 郭书巧. 转运体在百草枯抗性中的应用 [J]. 分子植物育种, 2009, 7(5): 1010-1014.
[11]陈建明, 俞晓平, 程家安. 叶绿素荧光动力学及其在植物抗逆生理研究中的应用 [J]. 浙江农业学报, 2006, 18(1): 51-55.
[12]付连双, 谢甫绨. 土壤水分对越冬期间冬小麦叶绿素荧光参数的影响[J]. 东北农业大学学报, 2011, 42(4): 36-41.
[13]Norman M A, Fuerst E P, Smeda R J, et al. Evaluation of paraquat resistance mechanisms inConyza[J]. Pesticide Biochemistry and Physiology, 1993, 46(3):236-249.
[14]Darkó é, Lehoczki E, Szigeti Z. Nuclear coded protein can be involved in the mechanism of paraquat resistance [J]. Biologia. Plantarum, 1994, 36: S342.
[15]Su Y H, Frommer W B, Ludewig U. Molecular and functional characterization of a family of amino acid transporters fromArabidopsis[J]. Plant Physiology, 2004, 136(2): 3104-3113.
[16]Yerushalmi H, Lebendiker M, Shuldiner S. EmrE, an Escherichia coli 12-kDa multidrug transporter, exchanges toxic cations and H+and is soluble in organic solvents [J]. The Journal of Biological Chemistry, 1995, 270(12): 6856-6863.
[17]Szigeti Z. Mechanism of paraquat resistance-from the antioxidant enzymes to the transporters[J]. Acta Biologica Szegediensis, 2005, 49(1-2): 177-179.
(责任编辑 李山云)
Preliminary Study on Resistance Level ofEleusineindicato Paraquat
SHEN Xue-feng, HU Fang, CHEN Yong*, LI Ying-hui, HAN Cheng-chou
(Weed Research Laboratory, South China Agricultural University, Guangdong Guangzhou 510642, China)
To investigate the resistance level ofEleusineindicato paraquat, the susceptible level ofEleusineindicato paraquat, values ofFv/Fmand amplification of resistance genes was conducted. The results showed that the paraquat concentration causing 50 % inhibition of survival rate (ED50) in resistant (R) and susceptible (S) biotypes ofE.indicawas 0.1029 and 6.1120 kg/hm2, and the relative resistance index [ED50(R) / ED50(S)] was 59.48. The values ofFv/Fmwas recovered in R biotype, but decreased with time in S biotype 3 h after spraying paraquat. However, it wasn’t recovered in R biotype after treating with 0.1 μmol/L nitrate + 1 kg/hm2paraquat. It was speculated that the paraquat was transported in R biotype by transporter. Provided primers specific to the PCR template sequence, the DNA of 1021 bp gets from R biotype, but S biotype was not. The nucleotide sequence of EST fragments was 96 % homologous toSorghumbicolorhypothetical protein with BLAST. Protein sequence alignments was 96 % homologous toZeamaysgene of transporter revealed one amino acid position, where organic cation transporters.
Eleusineindica; Paraquat; Resistance level; Transporter; Chlorophyll fluorescence
1001-4829(2016)08-1875-04
10.16213/j.cnki.scjas.2016.08.021
2015-10-22
国家自然科学基金资助项目(31272054,31471788);公益性行业(农业)科研专项(201303031)
沈雪峰(1982-),男,河南商丘人,讲师,主要从事作物栽培与生理研究,E-mail: xuefengshen@126.com,*为通讯作者。
S451.1
A
