荔枝花芽分化对其源叶光合作用及碳氮物质代谢的影响
2016-12-19张红娜苏钻贤陈厚彬
张红娜,苏钻贤,陈厚彬
(1.中国热带农业科学院南亚热带作物研究所/农业部热带果树生物学重点实验室,广东 湛江 524091;2.华南农业大学园艺学院,广东 广州 510642)
荔枝花芽分化对其源叶光合作用及碳氮物质代谢的影响
张红娜1,苏钻贤2,陈厚彬2
(1.中国热带农业科学院南亚热带作物研究所/农业部热带果树生物学重点实验室,广东 湛江 524091;2.华南农业大学园艺学院,广东 广州 510642)
为研究荔枝花芽分化对其源叶光合及同化产物合成的影响,以妃子笑荔枝为材料,比较分析同一生境下成花枝条与未成花枝条叶片的光合参数和碳氮物质的变化。结果表明,荔枝成花枝条叶片的净光合速率(2.39 μmol/m2·s)极显著地高于未成花枝条的(1.60 μmol/m2·s);同时,成花枝条的气孔导度、胞间CO2浓度、蒸腾速率和水分利用率均高于未成花枝条。荔枝花芽分化阶段,成花枝条叶片中的可溶性总糖含量和C/N比值明显高于未成花枝条,而总氮含量则是未成花枝条明显高于成花枝条。妃子笑荔枝花芽分化期间,其成花枝条潜在的光合能力要高于无花枝条;而源叶同化产物合成能力增加,则有助于为花芽发育提供充足的物质基础,以保证荔枝成花的顺利进行。
荔枝;花芽分化;光合作用;碳氮物质
植物“源”叶片的光合作用为“库”组织的代谢活动及其生长发育提供了动力[1]。“库”组织对光合产物的需求也对“源”叶片的光合作用有很大影响[2-4]。长期以来,果实作为强“库”对“源”叶片光合作用的影响是研究植物“库源”关系的重要课题[5-11]。目前,一种观点认为“库”组织的存在能提高“源”叶片的光合速率,并可加速叶片中的光合产物向外转运[7-9];但另一种观点认为改变“库源”关系对“源”叶片的光合速率无显著影响[10-11],说明“库源”之间的相互作用会因植物或外界条件的不同而有所不同。花芽分化是果树生长重要的生理活动之一,同时也是复杂的形态建成过程,花芽分化过程是否对果树叶片的光合作用产生影响受到研究者的广泛关注[12-13]。
荔枝(Litchi chinensis Sonn.)属无患子科荔枝属常绿果树,其花属于混合花序,花序长度在10~40 cm之间,正常花穗小花数量200~1 500朵,最多者可达4 000多以上,花量巨大[14]。荔枝花芽的生理分化过程无需消耗大量的碳素营养,但以白点为转折点的花发端实际上启动了一系列大量消耗碳素营养的生物过程,如花芽形态分化、花序发育、花器官构建等[14]。因此,荔枝的花芽作为一个“库”组织,在其发育过程中需要有一个稳定的“源”为其提供养分。已有研究表明,荔枝“源”叶的光合速率会受到果实“库”组织的影响,疏果处理会导致荔枝叶片光合速率明显下降,且离果实越近的叶片光合速率越高[15]。但是荔枝花芽的存在是否会对其“源”叶的光合速率及碳氮物质的积累产生影响,这个问题值得进一步研究。因此,阐明这些问题对弄清荔枝开花过程中的能量代谢以及如何提高开花质量有重要的理论与实践意义。
1 材料与方法
1.1 试验材料
试验在华南农业大学荔枝种质资源圃进行。供试品种为妃子笑,树龄9~10年,常规管理,随机选取健壮、长势及发育状况基本一致的3株作为试验用树。每株树在树冠外围东南西北向随机选取当年生枝梢10枝,进行定枝定位观察顶端花芽分化的物候期,于荔枝末次枝梢开始现白点时,每个单株从东、南、西、北4个方向选取长势相同的有白点和无白点枝条各5个,3株重复,共60个枝条,每个枝条分别做好标记。跟踪观察每个枝条顶芽的生长状况,1个月后确定所选枝条的成花情况。后续观察确认本试验中所选的有白点枝条均发育成为有花枝条,而无白点枝条的顶芽仍然保持未萌动状态。记白点与无白点枝条为成花枝条与未成花枝条两种处理。
1.2 试验方法
在出现白点后的第0、5、10、15和20 d上午9:00~11:00,对标记好的枝条中部成熟叶片进行活体测定光合特性和叶绿素指数,每次测定5个重复数据,并于当天分别选取各植株标记好的成熟叶片用冰盒保存带回实验室。采集的叶片用清水、蒸馏水清洗干净,置于烘箱中105℃杀青15 min后65℃烘干24 h,再用粉碎机粉碎,过0.15 mm孔径筛,装入封口袋并置于干燥器中备用。
1.3 测定指标及方法
1.3.1 光合特性测定 在出现白点后10 d上午9:00~11:00,对标记好的枝条中部成熟叶片测定气体交换参数和叶绿素指数。气体交换参数采用美国Li-cor公司生产的Li-6400便携式光合测定系统进行测定。测定时光合有效辐射(PAR)设为1 200 μmol/m2·s、大气CO2浓度(Ca)设为400 μmol/mol。大气温度(Ta)、叶温(TL)、相对湿度(RH)等参数均采用仪器自控系统控制,随外界环境变化而变化。同时使用SPAD-502型号便携式叶绿素仪对叶绿素指数进行测定。
1.3.2 碳水化合物含量测定 可溶性总糖(以干重计)含量采用蒽酮-硫酸法[16]测定。淀粉含量测定采用碘显色法[17]测定。总蛋白含量(以干重计)测定采用凯式定氮法[18]测定。
1.4 统计分析
试验数据采用Excel 2010、SigmaPlot10.0和SPSS16.0软件进行数据处理及相关性分析,用t检验法对各参数进行显著性检验。
2 结果与分析
2.1 光合参数比较分析
从表1可以看出,荔枝成花枝条叶片的净光合速率(Pn,2.39 μmol/m2·s)极显著高于未成花枝条叶片(1.60 μmol/m2·s);成花枝条的气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)和水分利用率(WUE)均高于未成花枝条,其中Gs达到显著差异,Tr达到极显著差异。

表1 成花枝条与未成花枝条叶片的净光合速率及部分光合参数
2.2 叶绿素指数(SPAD)分析
SPAD可以表征叶片叶绿素的含量[19]。从图1可以看出,荔枝成花枝条的SPAD值无明显变化,但未成花枝条叶片的SPAD在白点出现后的20 d内始终低于成花枝条,且在10 d左右有明显下降趋势,随后又有所回升。
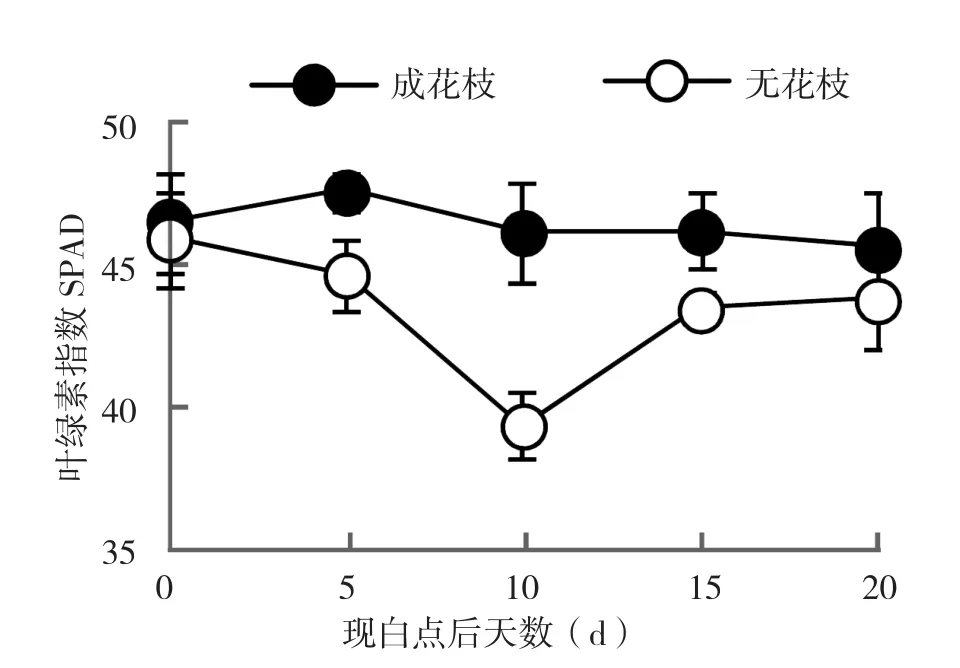
图1 成花枝条与未成花枝条叶片叶绿素指数变化
2.3 碳水化合物和总氮含量分析
2.3.1 可溶性总糖含量 由图2可知,荔枝花芽分化阶段成花枝条和未成花枝条叶片中可溶性总糖含量有较大差异。在白点出现后的一段时间内,荔枝成花枝条叶片中的可溶性总糖含量一直维持在30 mg/g左右的水平。未成花枝条中可溶性总糖含量一直维持在相对较低的水平,现白点后逐渐降低,10 d时达到最低水平,随后又有所上升。
2.3.2 淀粉含量 从图3可以看出,成花枝条与无花枝条中淀粉含量的变化趋势恰好相反。出现白点 5 d后,成花枝条叶片中的淀粉含量急剧下降,10 d后减幅明显下降,20 d时达到最低水平。而无花枝条中淀粉含量在现白点时明显低于成花枝条,5 d后淀粉含量急剧增加,10 d时明显超过对照,随后一直维持在较高水平。2.3.3 总氮含量分析 由图4可知,荔枝花芽分化阶段成花枝条和未成花枝条叶片中总氮含量存在较大差异,白点出现后,未成花枝条叶片中总氮的含量趋于稳定;而成花枝条叶片的总氮含量明显低于未成花枝条,且含量随着成花的发生继续下降,在20 d时达到最低水平。
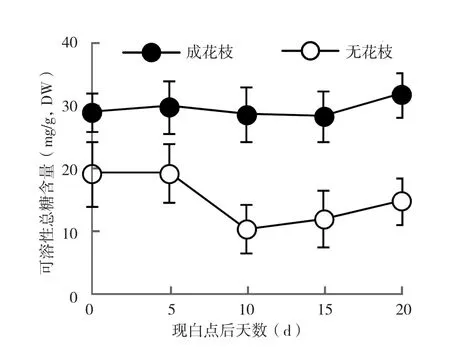
图2 成花枝条与未成花枝条叶片可溶性糖含量变化
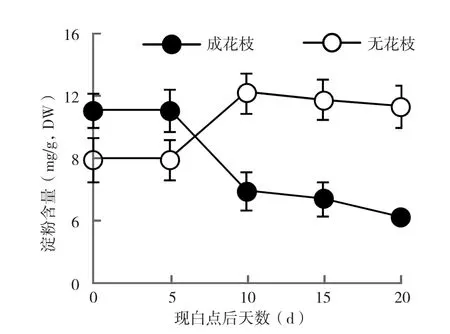
图3 成花枝条与未成花枝条叶片淀粉含量变化
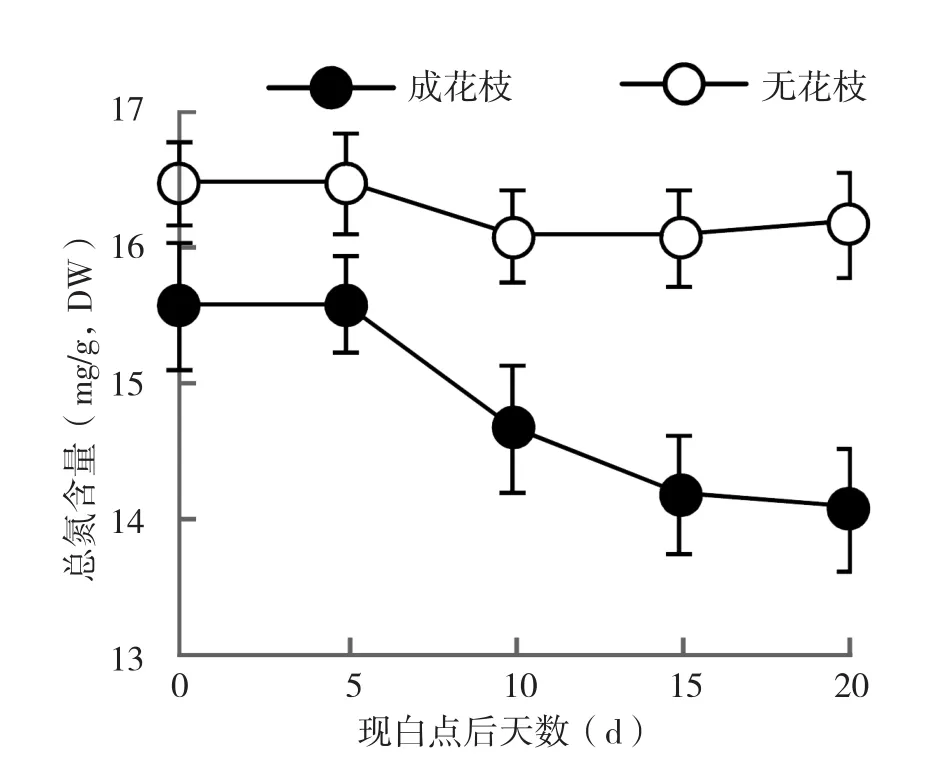
图4 成花枝条与未成花枝条叶片总氮含量变化
2.3.4 C/N比值 C/N比是反映植物营养情况和生长状态的重要指标。从图5可以看出,荔枝花芽分化阶段成花枝条和未成花枝条叶片中C/N比值有一定差别,但变化趋势基本一致,即先降低又升高;白点出现后荔枝成花枝条叶片中的C/N比值明显高于未成花枝条,说明高C/N比值是影响荔枝成花的一个重要因素。
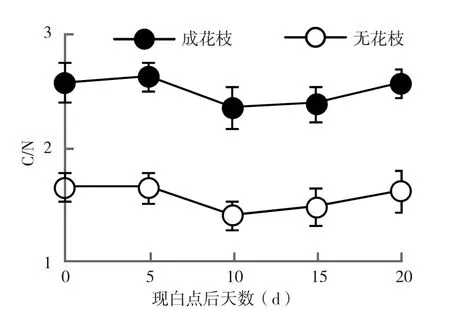
图5 成花枝条与未成花枝条叶片C/N比变化
3 结论与讨论
以往一些关于葡萄[1]、桃[20-21]、砂糖桔[22]等果树的研究表明,果实的存在会提高叶片的光合效率。当摘掉草莓[9]和桃[20]果实后,会导致其叶片光合速率大幅下降,原因可能是去除“库”组织后破坏了原本的“库-源”平衡,导致光合产物在叶片中积累,对叶片光合作用产生反馈抑制。本研究结果表明,妃子笑成花枝条叶片的光合速率极显著高于未成花枝条,说明荔枝花芽分化期间其花芽作为“库”组织存在对其“源”叶的净光合速率产生一定的促进作用。在研究植物库源关系时,许多学者发现叶片Pn与Gs间存在密切的正相关关系[23-25],“库强”降低首先会导致叶片气孔导度的下降,随后由于蒸腾速率的降低,造成叶片温度升高,从而调节光合系统的活性,最终使光合作用产生下调[6,24]。本试验中,除Pn外,妃子笑成花枝条的Gs、Ci、Tr和WUE均高于未成花枝条,其中花芽的存在显著提高了叶片的Gs和Tr,这可能是妃子笑成花枝条的Pn高于未成花枝条的原因之一。叶绿素为植物叶片进行光合作用的重要物质之一,其含量与光合速率有密切关系[26]。有研究表明在正常生长条件下,水稻叶片的叶绿素含量与其光合速率呈极显著正相关[27],而叶绿素指数可以用来表征叶片内的叶绿素含量[28]。本研究中,荔枝成花枝条叶片的叶绿素指数明显高于未成花枝条,表明妃子笑成花枝条潜在的光合潜能高于未成花枝条。
光合作用是果树生长发育的基础,果树积累的光合产物多则有利于树体健壮生长、根系发达、枝条充实、花芽饱满、产量高,且碳水化合物是光合作用的直接产物,对植物的成花结果也有十分重要的意义。研究表明,荔枝结果母枝的营养水平可以直接影响其成花和坐果[29-30]。Menzel等[31]也证明成花的荔枝树体内淀粉含量明显高于维持营养生长的植株。本研究中,妃子笑荔枝成花枝条和未成花枝条叶片中主要碳水化合物和总氮含量存在较大差异。出现白点后,妃子笑成花枝条叶片中的可溶性总糖含量明显高于未成花枝条,总体呈缓慢增加的趋势;而成花枝条叶片中的淀粉含量则逐渐降低,变化趋势与未成花枝条相反,这可能是叶片中储存的淀粉水解为可溶性糖导致的,而可溶性总糖含量的增加能提高细胞液浓度,有利于促进花芽分化;同一时期未成花枝条中的可溶性总糖含量要远低于成花枝条,这也反映了成花枝条的光合能力高于未成花枝条,以积累更多的光合产物用于花芽的发育。植物体内含氮化合物与同化糖类含量的比例是决定植物花芽分化的重要因素之一[32]。高的C/N比有利于荔枝的花芽分化,荔枝生产中常通过修枝或断根阻止氮素营养的吸收或通过环剥或环切树干提高C/N比,从而达到促进开花的目的。在妃子笑花芽分化期间,成花枝条叶片中淀粉、可溶性总糖的高含量和总氮的低水平,可能是花芽生理分化的重要营养基础。
[1]Chaumont M,Morot-Gaudry J F,Foyer C H. Seasonal and diurnal changes in photosynthesis and carbon partitioning in Vitis vinifera leaves in vines with and without fruit[J]. Journal of Experimental Botany,1994,45:1235-1243.
[2]李卫东,李绍华,吴本宏,等. 果实不同发育阶段去果对桃源叶光合作用的影响[J]. 中国农业科学,2005,38(3):565-570.
[3]Iqbal N,Masood A,Khan N. Analyzing the significance of defoliation in growth,photosynthetic compensation and source-sink relations[J]. Photosynthetica,2012,50(2):161-170.
[4]Nebauer S G,Renau-Morata B,Guardiola J L,et al. Photosynthesis down-regulation precedes carbohydrate accumulation under sink limitation in citrus[J]. Tree Physiology,2011,31(2):169-177.
[5]Proietti P,Nasini L,Famiani F. Effect of different leaf-to-fruit ratios on photosynthesis and fruit growth in olive(Olea europaea L. )[J]. Photosynthetica,2006,44(2):275-285.
[6]程杰山,王利军,蒋爱丽,等. 果树库源关系改变对源叶光合作用的影响机制研究进展[J].中国农学通报,2014,30(19):75-80.
[7]袁军,石斌,吴泽龙,等. 不同库源关系对油茶光合作用及果实品质的影响[J]. 植物生理学报,2015,51(8):1287-1292.
[8]朱振家,姜成英,史艳虎,等. 库源比改变对油橄榄产量及源叶光合作用的调节[J]. 中国农业科学,2015,48(3):546-554.
[9]付晓记,艾沙江. 买买提,等. 草莓去果降低库力对叶片光合特性和光破坏防御系统的影响[J]. 园艺学报,2011,38(7):1267-1274.
[10]Roper T R,Keller J D,Loescher W H,et al. Photosynthesis and carbohydrate partitioning in sweet cherry:fruiting effects[J]. Physiologia Plantarum,1988,72:42-47.
[11]Proietti P. Effect of fruiting on leaf exchange in olive(Olea europaea L. )[J]. Photosynthetica,2000,38:397-402.
[12]丰亚南,郑国生,王宗正,等. 牡丹开花前后碳水化合物的分配与光合速率的关系[J]. 园艺学报,2007,34(1):153-156.
[13]孔德良,严宁,胡虹. 开花对两种杓兰光合作用和同化产物分配的影响[J]. 云南植物研究,2006,28(6):639-644.
[14]李建国. 荔枝学[M]. 北京:中国农业出版社,2007.
[15]Hieke S,Menzel C M,Doogan V J,et al. The relationship between yield and assimilate supply in lychee(Litchi chinensis)[J]. Journal of Horticultural Science and Biotechnology,2002,77(3):326-332.
[16]邹琦. 植物生理学实验指导[M]. 北京:中国农业大学出版社,2000.
[17]徐昌杰,陈文峻,陈昆松,等. 淀粉含量测定的一种简便方法—— 碘显色法[J]. 生物技术,1998,8(2):41-43.
[18]母建华. 基于光谱分析的茶鲜叶全氮含量快速检测技术[D]. 镇江:江苏大学,2008.
[19]谢华,沈荣开,徐成剑,等. 水、氮效应与叶绿素关系研究[J]. 中国农村水利水电,2003(8):40-43.
[20]彭丽丽,姜卫兵,韩健,等. 去果(花)对不同成熟期桃叶片可溶性糖积累的影响[J]. 江苏农业学报,2013,29(4):857-863.
[21]朱亚静,李绍华,王红清,等. 果实的有无对桃叶片净光合效率及相关生理反应的影响[J].园艺学报,2005,32(1):11-14.
[22]黄永敬,唐小浪,马培恰,等. 库-源调节对沙糖桔源叶光合特性的影响[J]. 广东农业科学,2009,36(8):93-95.
[23]Cheng J S,Li W D,Fan P G,et al. Accumulation of end products in source leaves affects photosynthetic rate in peach via alteration of stomatal conductance and photosynthetic efficiency[J]. Journal of the American Society for Horticultural Science,2009,134(6):667-676.
[24]Li W D,Duan W,Fan P G,et al. Photosynthesis in response to sink--source activity and in relation to end products and activities of metabolic enzymes in peach trees[J]. Tree Physiology,2007,27(9):1307-1318.
[25]Cheng Y H,Arakawa O,Kasai M,et al. Analysis of reduced photosynthesis in the apple leaf under sink-limited conditions due to girdling[J]. Journal of the Japanese Society for Horticultural Science,2008,77(2):115-121.
[26]谭新星,许大全. 叶绿素缺乏的大麦突变体的光合作用和叶绿素荧光[J]. 植物生理学报,1996,22(1):51-57.
[27]胡颂平,王正功,张琳,等. 干旱胁迫下水稻叶片光合速率与叶绿素含量的相关性及其基因定位[J]. 中国生物化学与分子生物学报,2007,23(11):926-932.
[28]王树良,李银国. 测定柑桔叶片叶绿素含量的两种方法比较[J]. 中国南方果树,1996,25(2):24.
[29]邱燕萍,袁沛元,张展薇,等. 荔枝不同秋梢结果母技的营养及其对成花与坐果的影响[J].广东农业科学,1995(2):22-25
[30]彭坚,李永红,席嘉宾,等. 荔枝大小年叶片营养比较研究[J]. 中国果树,2004(3):31-34.[31]Menzel C M,Rasmussen Ts,Simpson D R. Carbohydrate reserves in lychee trees(Litchi chinensis Sonn. )[J]. Journal of Horticultural Science,1995,70(2):245-255.
[32]Priestley C. The annual turnover resources in young olive trees[J]. Horticultural Science,1977,52(1):105-112.
(责任编辑 白雪娜)
Effects of flower bud differentiation on photosynthesis and carbon,nitrogen metabolism in Litchi(Litchi chinensis Sonn.)
ZHANG Hong-na1,SU Zuan-xian2,CHEN Hou-bin2
(1.Key Laboratory of Ministry of Agriculture for Tropical Fruit Biology/South Subtropical Crops Research Institute,CATAS,Zhanjiang 524091,China;2.College of Horticulture,South China Agricultural University,Guangzhou 510642,China)
To investigate the effects of flower bud differentiation on photosynthesis and assimilation products,we analyzed the variation in the concentrations of leaf photosynthesis,carbon and nitrogen metabolism of ‘Feizixiao’litchi trees. The results showed that Photosynthesis (Pn,2.39 μmol/m2·s) was significantly higher in flowering shoot than that in vegetative shoot (1.60 μmol/m2·s). In addition,it was found that the stomatal conductance(Gs),internal CO2concentration (Ci),transpiration rate (Tr),and water use efficiency (WUE) of flowering shoot were higher than those of vegetative shoot. During the flower bud differentiation stage,the contents of soluble sugars and C/N ratio of flowering shoot were significantly higher than those of vegetative shoot,whereas total nitrogen had a contrary tendency. The results suggested that there was higher potential photosynthetic capacity in flowering shoot than that in vegetative shoot. Litchi flowering was able to promote assimilation production,more sugar gave out a rich basis for the development of flower buds and to keep floral formation moving along smoothly.
Litchi (Litchi chinensis Sonn.);flower bud differentiation;photosynthesis;carbon and nitrogen
S667.2
A
1004-874X(2016)11-0050-06
2016-08-01
国家荔枝龙眼现代农业产业技术体系建设专项资金(CARS-33);广东省现代农业(重要热带作物)产业技术研发中心建设项目
张红娜(1986-),女,博士,助理研究员,E-mail:zhanghongna6789@163.com
陈厚彬(1963-),男,博士,研究员,E-mail:hbchen@scau.edu.cn
张红娜,苏钻贤,陈厚彬.荔枝花芽分化对其源叶光合作用及碳氮物质代谢的影响[J].广东农业科学,2016,43(11):50-55.
