节节麦大穗大粒相关农艺性状的遗传分析
2016-12-15朱欣果郑建敏吕茂应杨武云万洪深
刘 磊,王 琴,朱欣果,郑建敏,吕茂应,杨武云,李 俊,万洪深
(1.四川省农业科学院作物研究所/农业部西南地区小麦生物学与遗传育种重点实验室,四川成都 610066;2.蓬安县农牧业局种子管理站, 四川蓬安 637800)
节节麦大穗大粒相关农艺性状的遗传分析
刘 磊1,王 琴1,朱欣果1,郑建敏1,吕茂应2,杨武云1,李 俊1,万洪深1
(1.四川省农业科学院作物研究所/农业部西南地区小麦生物学与遗传育种重点实验室,四川成都 610066;2.蓬安县农牧业局种子管理站, 四川蓬安 637800)
节节麦(Aegilopstauschii, DD) 是六倍体普通小麦D基因组的祖先,其自然类群中含有丰富的抗逆、高产基因,利用其与四倍体硬粒小麦合成的六倍体小麦在现代小麦育种中得到了愈来愈多的应用。本课题在野生节节麦类群中发现了大穗、大粒材料AT462,利用其作母本与节节麦材料AT18(强分蘖)杂交;构建了F2、F3群体,通过调查亲本和群体单株的穗长、小穗数、粒长、粒宽和粒重等表型,对这些穗部性状进行了相关性分析和遗传分析。结果表明:(1)在F2和F3群体中,粒重、粒长与穗长之间不存在显著相关性,而且穗长与粒宽之间在两个群体中的平均相关系数绝对值小于0.1,粒重与小穗数之间的相关系数绝对值小于0.2,表明节节麦大粒相关性状不受穗长的影响,受小穗数影响也较小;(2)采用F2单世代分离分析的方法对节节麦AT462×AT18的F2群体大穗、大粒相关性状进行遗传分析,其中穗长受2对具有加性效应的主效基因控制;粒重和小穗数均同时受2对基因的加性效应、显性效应以及互作效应控制,其中加性效应占主导地位;粒长、粒宽均受2对基因的加性效应、显性效应以及互作效应控制,且三种效应较为均衡。这说明控制节节麦粒重、穗长、小穗数等产量性状相关基因的加性效应在遗传中占主导地位,在育种中较易利用,且其主效基因的遗传力达0.9。
节节麦;大穗;大粒;遗传分析
穗是小麦产量形成的重要器官,大穗、大粒特性是形成超高产小麦的重要基础之一。在普通小麦穗部相关农艺性状中,小穗数、穗粒数与粒重往往呈显著负相关[1],穗长与小穗数、粒重呈正相关[1-2]。大穗品种表现为穗粒数与粒重之间的协调,往往具有较高的穗容量。随着我国小麦品种农艺性状的演化,品种的穗容量持续显著上升[1]。大穗型小麦品种在高肥条件下具有更高的高产潜力[3]。在西南麦区以川麦42为代表的骨干亲本成为超高产小麦品种选育的大穗骨架[4]。节节麦(Aegilopstauschii) 是六倍体普通小麦D基因组的祖先。现已证明,参与普通六倍体小麦自然进化的节节麦类群单一[5],较A、B基因组,普通六倍体小麦D基因组的遗传基础狭窄。自然界中现有的节节麦类群具有与抗逆[6]、高产相关的基因[7-8],利用其与四倍体硬粒小麦合成的人工六倍体小麦作为桥梁[9],来自节节麦的一些优良抗性基因被成功导入到六倍体小麦中[10-11],并在育种中得到了很好的利用[12-13]。前人对节节麦产量性状方面的直接研究较少,往往通过其与四倍体合成倍性稳定的六倍体后进行分子遗传相关的研究,而后发现在人工合成小麦的节节麦基因组上也具有与增产相关的基因[14],这些研究对节节麦中与增产潜力相关的基因的发掘没有针对性,制约了人工合成小麦在普通小麦遗传改良上的利用。本课题组从节节麦野生资源中筛选出了一个节节麦大穗、大粒品系AT462,其穗长、粒重显著高于其他节节麦材料。为了解其大穗、大粒性状的遗传特性,利用其与对照节节麦材料AT18构建了F2、F3分离群体,对其穗部相关性状进行了调查与相关性分析,并利用F2单世代分离分析的方法[15]对其进行遗传分析以及相关参数估计,以期为节节麦大穗相关基因在六倍体小麦遗传育种中的利用提供理论依据。
1 材料与方法
1.1 材 料
本研究以节节麦AT462(大穗、大粒)为母本、以节节麦AT18(强分蘖)为父本进行杂交、自交获得F2群体,群体大小为230单株,下一年在原有F2单株的基础上以单粒传的方式形成F3群体;其中亲本AT462、AT18由四川省农业科学院作物研究所杨武云研究员收集获得。
1.2 田间试验与农艺性状调查
田间试验分别于2013-2014、2014-2015年设立在四川省农业科学院郫县试验基地。行长2 m,行距0.5 m,每行5穴,每穴1苗。正常田间管理。2013-2014年试验材料为F2群体,2014-2015年为在上一年F2单株的基础上单粒传获得的F3群体。
灌浆期调查亲本及F2、F3群体各单株的穗长和小穗数,每个单株测量3~5个单穗并取平均值,测量后每个单穗用透明薄膜进行套袋,待成熟后进行收获、脱粒,最终每个单株大约收获100~200个籽粒。随机取50~100个籽粒测量,计算千粒重,重复测定3次并取平均值;同时利用数显游标卡尺随机测量20~50个籽粒的长和宽,取其平均值分别代表群体中每个单株粒长与粒宽。
1.3 统计与分析
采用IBM SPSS Statistics Version 22.0(运行平台为苹果OS X Version 10.9.4) 对表型数据进行t检验、相关性分析及次数分布作图。
遗传分析采用盖钧镒等[15]提出的主基因+多基因遗传模型进行F2群体单个分离世代的分析,软件包SEA-F2由章元明教授提供,计算时根据AIC(Akaike's Information Criterion) 值最小原则[16],具有最小AIC值且与其他模型AIC值差异较大的模型为相对最适模型,当具有多个较低AIC值的模型时,通过在均匀性U12、U22、U32检验、Sminov检验(nW2)和Kolmogorov检验(Dn)中所得到的显著性个数最少的模型为最适模型[17]。
2 结果与分析
2.1 亲本及群体的大穗相关性状的分布
物联网拓展了互联网应用发展的空间,推进了智能运输系统的发展,促进了物联网在现代物流领域的开发应用,尤其是在新鲜水果蔬菜等要求较高运输环境的物流运输系统的应用。采用物联网技术的运输系统,可促进交通运输和现代物流的安全,使驾驶员能在复杂多变的交通环境中操纵、行驶,以求最大限度地减少道路交通事故的发生,不断提高果蔬运输系统安全水平。
除小穗数外,大穗、大粒亲本AT462的表型值在两年中都显著大于亲本AT18,尤其是粒重、粒长和粒宽(表1)。亲本的表型值位于后代F2、F3群体表型最大值与最小值之间,群体呈现超亲分离,表明在两个亲本中具有多个增加效应的基因位点(表1)。对于粒重、粒长和粒宽来说,群体表型平均值位于两亲本表型值之间;而对于穗长来说,群体均值明显大于两个亲本的表型值(表1),说明两个亲本中可能都具有增加穗长的主效基因。在这些性状中,粒重、穗长及小穗数在群体中的变异系数相对较大,推测该群体中效应较大的基因之间可能发生了分离。
利用单样本Kolmogorov-Smirnov拟合度检验,结果显示,除穗长、小穗数外,其他性状表型在F2、F3群体中都呈正态分布,平均P值为0.200。从次数分布(图1)可以看出,F2、F3群体中小穗数在12.0~12.4区间上的个体数目最多,分别为47个(占总数的21.4%)、49个(占22.4%),相对于其他性状来说,其分布比较集中,与正态分布偏离较大;而对于粒重、粒长以及粒宽来说,其个体数目最大值均小于36,占总数的15%,分布类型接近正态分布。
2.2 F2、F3群体中各性状之间的相关性
F3是从F2群体单株上通过单粒传获得的,是F2:3家系中的一个单株,两代群体之间所调查性状均显著相关,相关系数为0.308~0.484(表2)。在F2、F3群体中,粒重与粒长、粒宽呈显著正相关,相关系数平均值在0.60以上;粒重与小穗数呈负相关,相关系数平均为0.20;在两个群体中,粒重、粒长和粒宽与穗长相互独立,不存在显著相关或者相关系数很小;穗长与小穗数在两个群体中都呈显著正相关,相关性系数达0.782(表2)。这些结果表明,穗长、小穗数对粒重的影响较小。
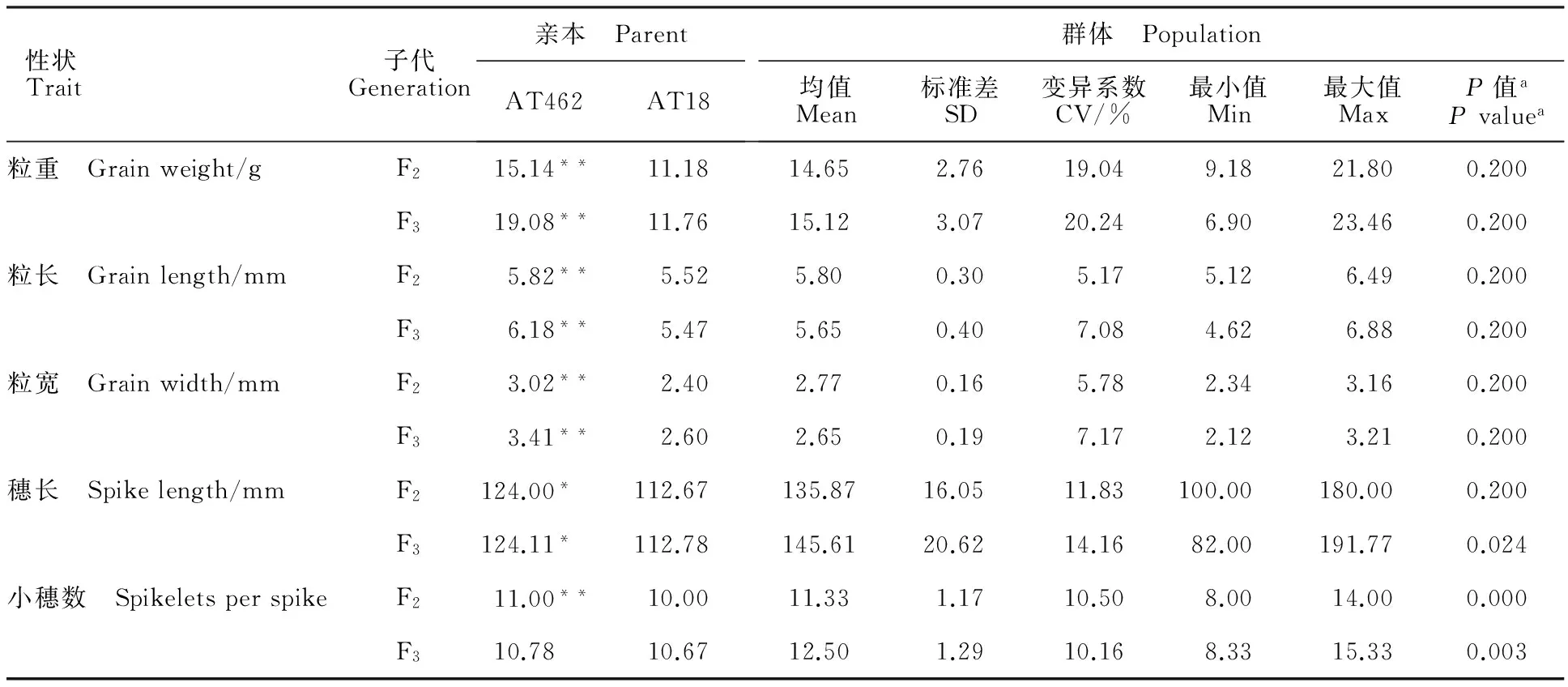
表1 亲本及其F2、F3群体大穗相关性状表型值的分布情况
a:单样本Kolmogorov-Smirnov拟合度检验,检验分布为正态分布(双侧),其结果经过Lilliefors显著水平修正;*和**分别表示亲本AT462与AT18在0.05和0.01水平上具有显著差异。
a:Corrected Lilliefors significance by one sample Kolmogorov-Smirnov test for normal distribution(two tailed test); * and ** indicate significance at 0.05 and 0.01 levels,respectively.
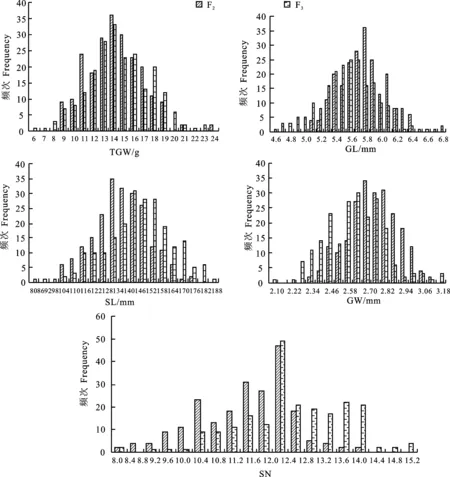
图1 F2、F3群体粒重(TGW)、粒长(GL)、粒宽(GW)、穗长(SL)和小穗数(SN)的次数分布
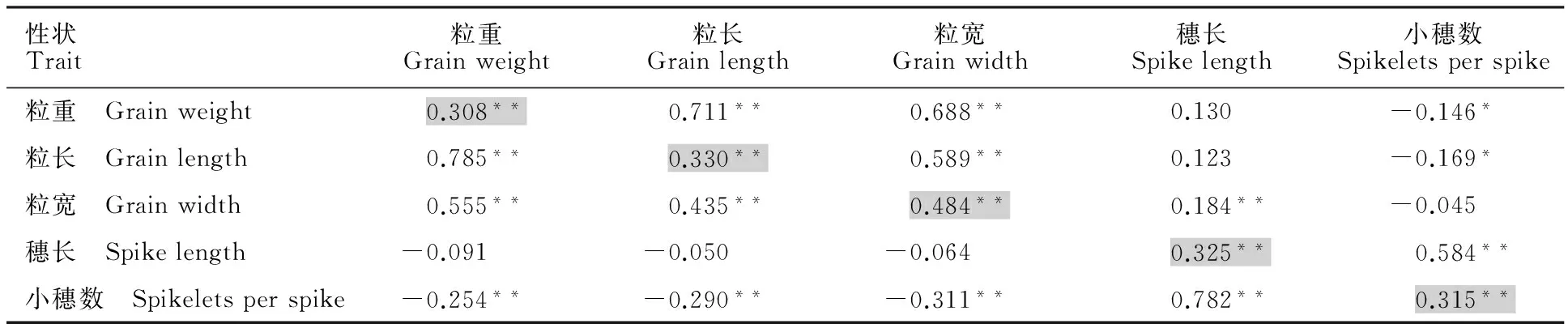
表2 F2、F3群体中粒重、粒长、粒宽、穗长与小穗数之间的关系
*:P<0.05;**:P<0.01;阴影单元格显示同一表型在F2与F3群体之间的相关性,其上部的数值表示F3群体的相关系数,下部的数值表示F2群体的相关系数。
*:P<0.05;**:P<0.01; Shadowed cells showed the correlation coefficients of the same traits between F2and F3populations, cells above the shadowed ones were correlation coefficients between different traits in F3population, and the below cells were for F2population.
2.3 F2单世代“主基因+多基因”遗传模型分析及其参数估计
在利用SEA-F2软件包进行遗传模型分析时,剔除了F2个体表型中个别极端离群值。根据AIC值最小原则,选取AIC值最小或者较为接近的模型作为备选模型。在粒重、粒长、粒宽及小穗数的单世代分离分析结果中,以2MG-ADI模型(2对加性-显性-上位性主基因模型)的AIC值最小,并且与其他模型之间的差异较大,为最适遗传模型;而穗长的最适遗传模型是2MG-A(2对加性主基因模型)(表3)。
利用最小二乘法估算各调查性状在最适模型下的遗传参数,结果(表4)表明,控制粒重、小穗数的2对主基因的遗传力均达0.90,第1对主基因的加性效应远远大于其显性效应,两者的效应比值大于5,第2对主基因加性效应与显性效应相当,两者比值在1~2之间,两个主效基因之间的互作效应相对较小。控制粒长、粒宽的2对主基因的遗传力分别为0.78、0.74,第1对主基因的加性效应大于显性效应,两个效应的比值在2左右,第2对主基因加性效应与显性效应相当,并且2对基因之间存在明显的互作。控制穗长的2对基因以加性效应为主,而且基因之间不存在显性和互作效应,主基因的遗传力高达0.94。总的来说,除粒长和粒宽外,节节麦穗长、粒重及小穗数等大穗相关性状基因的加性效应要大于其显性、互作效应,其中至少有1对基因的加性效应占有绝对优势,其效应值最大,更利于小麦常规育种。
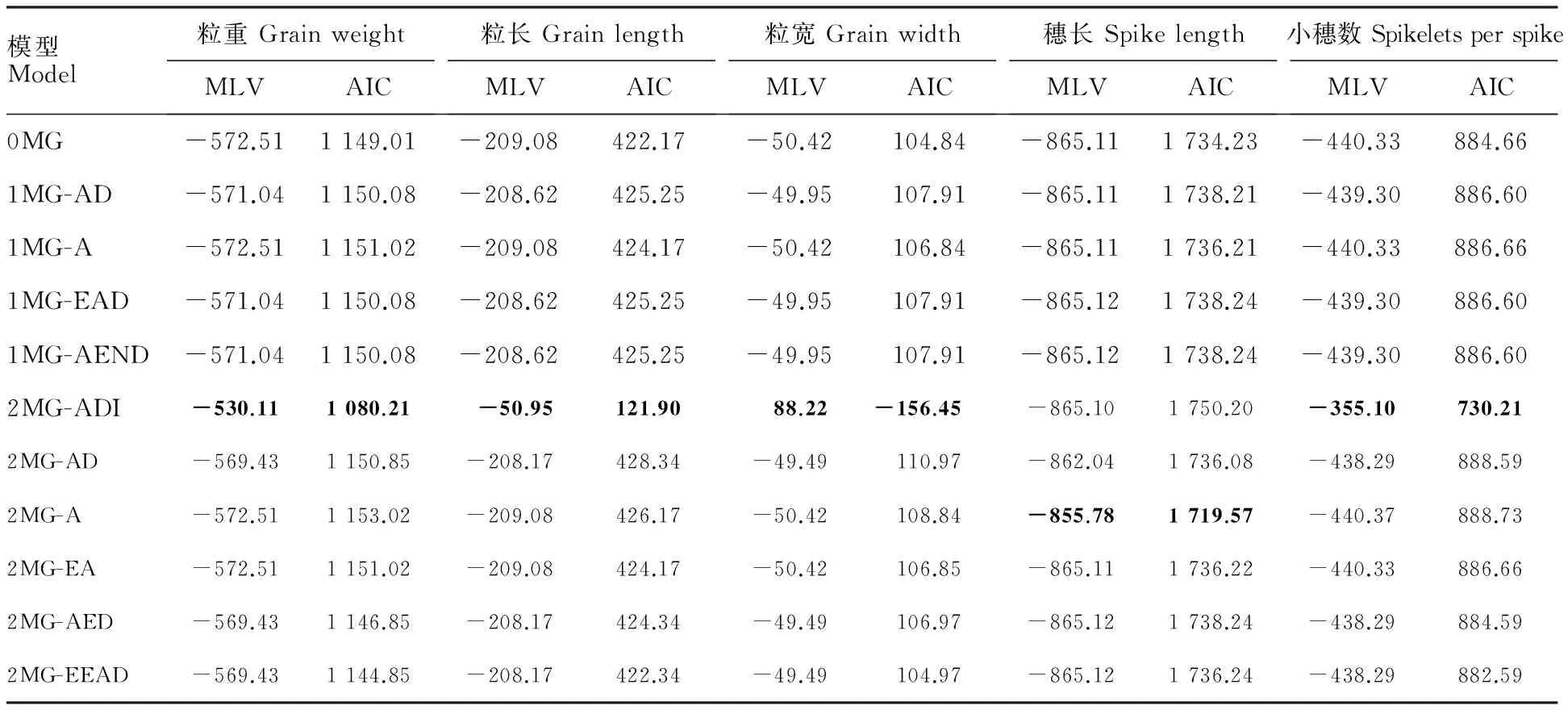
表3 F2群体粒重、粒长、粒宽、穗长和小穗数性状不同遗传模型的MLV和AIC值
MLV:极大对数似然函数值;粗体表示备选模型的MLV和AIC值。
MLV:log-maximum likelihood value; Bond latters were MLV and AIC for the selected optimal models, respectively.
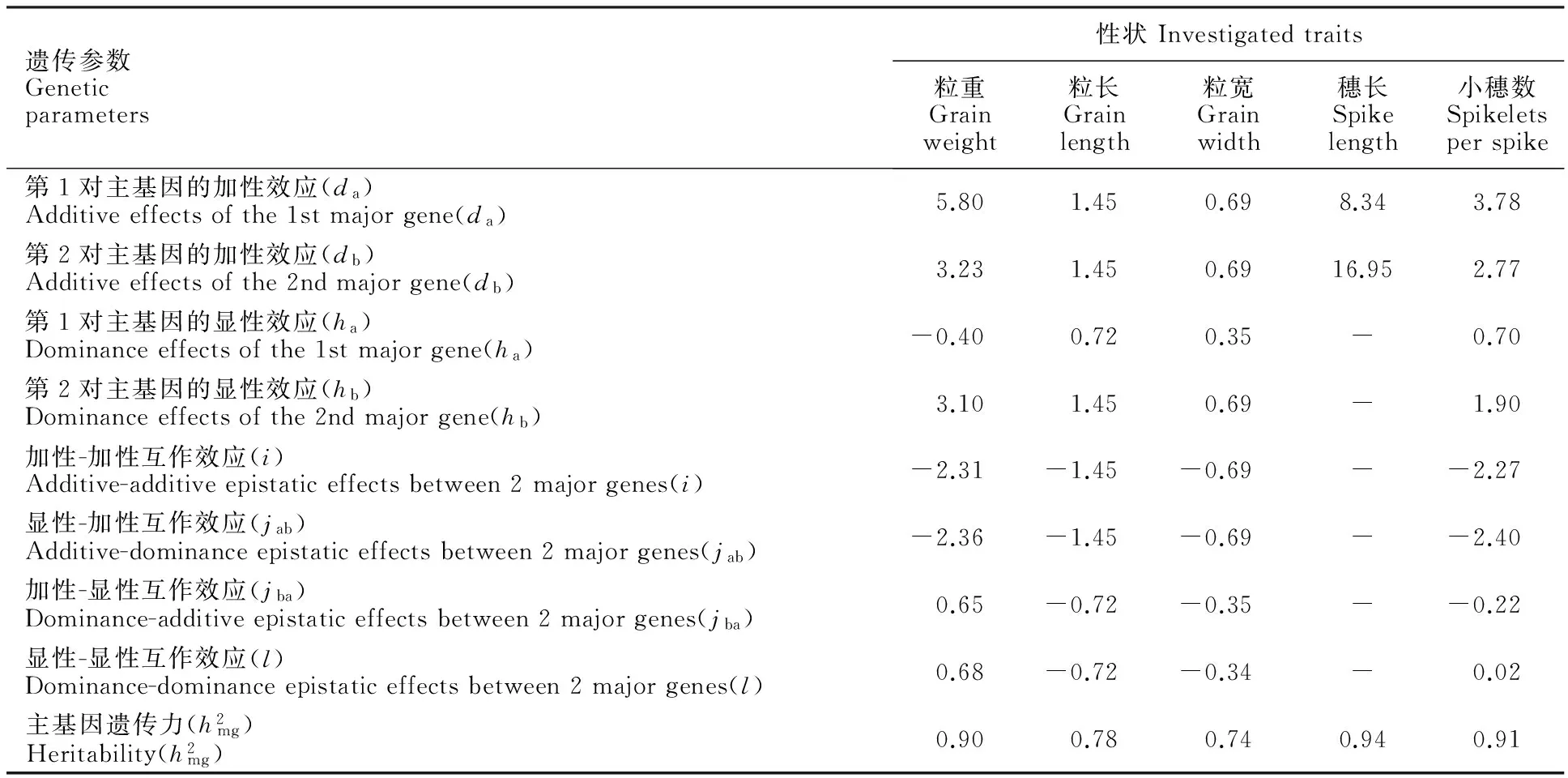
表4 F2群体粒重、粒长、粒宽、穗长和小穗数性状最适遗传模型的参数估计
3 讨 论
本研究利用大穗、大粒节节麦AT462与对照节节麦AT18杂交形成F2、F3群体,研究了节节麦的大穗、大粒性状在后代群体中的分离情况,并利用F2群体对这些产量相关性状进行了遗传分析,对于粒重、穗长以及小穗数而言,其基因的加性效应要大于非加性效应,这对节节麦的大穗、大粒相关基因的利用极为有利。
前人对节节麦大穗、大粒等产量相关农艺性状的研究较少,在对六倍体普通小麦穗部相关基因的研究中,与大穗相关的基因大多定位在A、B基因组,D组相对较少[1],这是由于参与普通小麦自然进化的节节麦类群较少,其多样性较低[5]。但随着普通小麦的自然进化与人工选择及人工合成小麦的应用,如今也有一些基因定位在D基因组上。如1D着丝粒附近的粒重QTL[18]、2D上的穗长基因[2, 19]、小穗数QTLs[19-20]、3D着丝粒附近千粒重QTL[21]、7DS上的千粒重QTL[14, 22]等,其中有一些大穗、大粒QTL位点是来自人工合成小麦,如人工合成小麦W7984的2DS位点具有增加穗长的效应[19],XX86的 Xgwm1220-7DS位点具有增加粒重的效应[22]。在本研究中,对F2群体的遗传分析显示,节节麦AT462后代群体的大穗、大粒特性至少受2对主效基因控制的,并且其遗传力都比较高。这些研究结果说明在节节麦的D基因组上具有大穗、大粒相关的基因位点。前人对人工合成小麦群体大穗、大粒等产量相关性状的QTL结果表明,节节麦大穗性状能够在六倍体小麦背景下正常表达[14, 19, 22]。
在节节麦大穗、大粒相关性状中,控制粒重和小穗数各自的2对主基因的平均加性效应值要显著大于其平均显性、互作效应值。前人对粒重配合力的研究也认为,千粒重的一般配合力方差要大于特殊配合力的方差,其主要受基因的加性效应和非加性效应控制,其中加性效应要大于非加性效应[23-25]。本研究中穗长受2对基因加性效应控制。庄巧生等[23]对多个农艺性状的配合力研究表明,穗长的一般配合力(加性效应)占比例最大。吴新义[26]对穗长近等基因系F1后代表型的分析发现,2D穗长基因表现为部分显性,具有基因剂量效应,其穗长接近两亲本的平均值,进一步说明2D穗长基因的加性效应远远大于其显性效应;粒长与粒宽在形态上与粒重显著相关。然而,本研究中粒长、粒宽性状主效基因的加性效应、显性效应及互作效应的分配情况与粒重遗传分析的结果有较大的区别,极有可能受不同基因控制。关于籽粒大小与形态的QTLs比较研究也表明粒重与籽粒形态在遗传上是独立的[27-28]。
综上,本研究利用F2、F3群体研究了节节麦AT462大穗、大粒相关性状的群体分布情况并对其进行了遗传分析,明确了与大穗、大粒紧密相关性状主要由基因的加性效应控制,可以通过合成六倍体小麦的途径发挥其在普通小麦育种中潜在的应用价值。
[1] 万洪深.小麦骨干亲本南大2419产量相关基因组区段的定位及其等位变异的效应[D].南京:南京农业大学,2013:50-56,108-120.
WAN H S.Yield-related genomic regions of founder wheat parent Nanda 2419 and the effects of their allelic variations [D].Nanjing:Nanjing Agricultural University,2013:50-56,108-120.
[2]WU X Y,CHENG R R,XUE S L,etal.Precise mapping of a quantitative trait locus interval for spike length and grain weight in bread wheat(TriticumaestivumL.) [J].MolecularBreeding,2014,33:129-138.
[3] 田纪春,邓志英,胡瑞波,等.不同类型超级小麦产量构成因素及籽粒产量的通径分析[J].作物学报,2006,32(11):1699-1705.
TIAN J C,DENG Z Y,HU R B,etal.Yield components of super wheat cultivars with different types and the path coefficient analysis on grain yield [J].ActaAgronomicaSinica,2006,32(11):1699-1705.
[4] 万洪深,李 俊,杨武云.西南麦区小麦骨干亲本川麦42的遗传构成与育种利用[C].第7届全国小麦遗传育种学术研讨会论文摘要集,中国郑州,2015:93.
WAN H S,LI J,YANG W Y.Genetic components of founder parent Chuanmai 42 and its utilization in wheat breeding of program of Southwest China [C].Collection of paper abstracts for 7th Chinese Wheat Genetics and Breeding Symposium,Zhengzhou,China,2015:93.
[5] WANG J R,LUO M C,CHEN Z X,etal.Aegilopstauschiisingle nucleotide polymorphisms shed light on the origins of wheat D-genome genetic diversity and pinpoint the geographic origin of hexaploid wheat [J].NewPhytologist,2013,198(3):925-937.
[6] 杨武云,余 毅,胡晓蓉,等.节节麦及其在小麦生物技术育种中的研究与应用[J].西南农业学报,1999,12(高新专辑):19-25.
YANG W Y,YU Y,HU X R,etal.Exploring useful genes inAegilopstauschiifor modern commercial wheat improvement by biotechnology [J].SouthwestChinaJournalofAgriculturalSciences,1999,12(AHTS):19-25.
[7]TER STEEGE M W,DEN OUDEN F M,LAMBERS H,etal.Genetic and physiological architecture of early vigor inAegilopstauschii,the D-genome donor of hexaploid wheat: A quantitative trait loci analysis [J].PlantPhysiology,2005,139:1078-1094.
[8]LI J,WEI H T,HU X R,etal.Identification of a high-yield introgression locus in Chuanmai 42 inherited from synthetic hexaploid wheat [J].ActaAgronomicaSinica,2011,37(2):255-261.
[9]YANG W Y,LIU D C,LI J,ZHANG L Q,etal.Synthetic hexaploid wheat and its utilization for wheat genetic improvement in China [J].JournalofGeneticsandGenomics,2009,36:539-546.
[10]杨武云,余 毅,胡晓蓉,等.节节麦抗白粉病基因直接转移及遗传表达[J].西南农业学报,2000,13(4):15-20.
YANG W Y,YU Y,HU X R,etal.Direct transfer of powdery mildew resistant gene fromAegilopstaushiiinto bread wheat and its genetic expression [J].SouthwestChinaJournalofAgriculturalSciences,2000,13(4):15-20.
[11]OLSON E L,ROUSE M N,PUMPHREY M O,etal.Introgression of stem rust resistance genes SrTA10187 and SrTA10171 fromAegilopstauschiito wheat [J].TheoreticalandAppliedGenetics,2013,126:2477-2484.
[12]LI J,WAN H S,YANG W Y.Synthetic hexaploid wheat enhances variation and adaptive evolution of bread wheat in breeding processes [J].JournalofSystematicsandEvolution,2014,52:735-742.
[13]WAN H S,YANG Y M,LI J,etal.Mapping a major QTL for hairy leaf sheath introgressed fromAegilopstauschiiand its association with enhanced grain yield in bread wheat [J].Euphytica,2015,205:275-285.
[14]HUANG X Q,CSTER H,GANAL M W,etal.Advanced backcross QTL analysis for the identification of quantitative trait loci alleles from wild relatives of wheat(TriticumaestivumL.) [J].TheoreticalandAppliedGenetics,2003,106:1379-1389.
[15]盖钧镒,章元明,王健康.植物数量性状遗传体系[M].北京: 科学出版社,2003: 96-101.
GAI J Y,ZHANG Y M,WANG J K.Genetic System of Quantitative Traits in Plants [M].Beijing: Science Press,2003:96-101.
[16]AKAIKE H.On Entropy Maximum Principle.In:Krishnaiah P R.(ed) Applications of Statistics[M].Amsterdam: North Holland Publishing,1977:27-41.
[17]曹锡文,刘 兵,章元明.植物数量性状分离分析Windows软件包SEA的研制[J].南京农业大学学报,2013,36(6):1-6.
CAO X W,LIU B,ZHANG Y M.SEA: a software package of segregation analysis of quantitative traits in plants [J].JournalofNanjingagriculturaluniversity,2013,36(6):1-6.
[18]MCINTYRE C L,MATHEWS K L,RATTEY A,etal.Molecular detection of genomic regions associated with grain yield and yield-related components in an elite bread wheat cross evaluated under irrigated and rainfed conditions [J].TheoreticalandAppliedGenetics,2010,120:527-541.
[19]KUMAR N,KULWAL P L,BALYAN H S,etal.QTL mapping for yield and yield contributing traits in two mapping populations of bread wheat [J].MolecularBreeding,2007,19:163-177.
[20]JIA H Y,WAN H S,YANG S H,etal.Genetic dissection of yield-related traits in a recombinant inbred line population created using a key breeding parent in China’s wheat breeding [J].TheoreticalandAppliedGenetics,2013,126:2123-2139.
[21]MCCARTNEY C A,SOMERS D J,HUMPHREYS D G,etal.Mapping quantitative trait loci controlling agronomic traits in the spring wheat cross RL4452בAC Domain’ [J].Genome,2005,48: 870-883.
[22]HUANG X Q,KEMPF H,GANAL M W,etal.Advanced backcross QTL analysis in progenies derived from a cross between a German elite winter wheat variety and a synthetic wheat(TriticumaestivumL.) [J].TheoreticalandAppliedGenetics,2004,109:933-943.
[23]庄巧生,王恒立,曾啓明,等.冬小麦亲本选配的研究Ⅰ.杂种第一代优势和配合力的分析[J].作物学报,1963,2(2):117-130.
ZHUANG Q S,WANG H L,ZENG Q M,etal.Studies on the choice of parents in wheat crosses.I.Hererosis and combining ability in F1hybrids [J].ActaAgronomicaSinica,1963,2(2):117-130.
[24]赵献林,康明辉,任明全,等.几个小麦品种(系)产量性状的配合力分析[J].华北农学报,1995,10(增刊):38-41.
ZHAO X L,KANG M H,REN M Q,etal.Analyses on combining ability of yield characters in common wheat [J].ActaAgriculturaeBoreali-Sinica,1995,10(S):38-41.
[25]HASSAN G,MOHAMMAD F,AFRIDI S S,etal.Combining ability in the F1generations of diallel cross for yield and yield components in wheat [J].SarhadJournalofAgriculture,2007,23:937-942.
[26]吴新义.小麦产量相关性状QTL近等基因系的选育及一个穗长QTL的精确定位[D].南京:南京农业大学,2013:60-69.
WU X Y.Development and evaluation of wheat yield-related QTL near-isogenic lines and precise mapping of a QTL for spike length [D].Nanjing: Nanjing Agricultural University,2013:60-69.
[27]RAMYA P,CHAUBAL A,KULKARNI K,etal.QTL mapping of 1000-kernel weight,kernel length,and kernel width in bread wheat(TriticumaestivumL.) [J].JournalofAppliedGenetics,2010,51:421-429.
[28]GEGAS V C,NAZARI A,GRIFFITHS S,etal.A genetic framework for grain size and shape variation in wheat [J].PlantCell,2010,22:1046-1056.
Genetic Analysis for Long-Head and Big-Kernel Related Traits fromAegilopstauschii
LIU Lei1, WANG Qin1, ZHU Xinguo1, ZHENG Jianmin1, LÜ Maoying2,YANG Wuyun1, LI Jun1, WAN Hongshen1
(1.Crop Research Institute, Sichuan Academy of Agricultural Sciences/Key Laboratory of Wheat Biology and Genetic Improvement on Southwestern China, Ministry of Agriculture, Chengdu, Sichuan 610066, China; 2.Corps Introduction and Reproduce Center, The Farming and Animal Husbandry Bureau of Peng’an County, Peng’an, Sichuan 637800, China)
Aegilopstauschii, the diploid D genome progenitor of the hexaploid wheat, has been used to improve modern common wheat through creating synthetic hexaploid wheat by crossing with durum wheat, as a bridging mechanism for wheat breeding and gene pool for environmental stress resistance and high yield. In this study, we obtained an accession AT462 with long-head and big-kernel in the wild tauschii gene pool and created the populations of F2and F3by crossing with the otherAegilopstauschiiaccession AT18. A total of 5 yield-related traits were investigated, including spike length, spikelets per spike, kernel length, width and weight. The results of correlation and genetic analysis are as follows:(1) In the F2and F3populations, there was no significant relationship between kernel-related traits including grain weight, kernel length and spike length. The absolute value of average correlation coefficient between spike length and kernel width was less than 0.1, and the absolute value between kernel weight and spikelet number per spike was less than 0.2. These results indicated that the expression of the big kernel related traits was not significantly influenced by spike length and less affected by spikelets per spike.(2) Kernel length was controlled by two pair of major genes with additive effects only with F2-generation separation genetic analysis. Both kernel weight and spikelets per spike were controlled by two major genes, and their additive effects were the strongest among the three kinds of genetic effects. The segregations of kernel length and kernel width were controlled by both additive, dominance and interaction effects of genes in F2population, where three types of genetic effects were distributed in a relatively equitable manner. These results suggested that the long-head and big-kernel related traits such kernel weight, spike length and spikelets per spike were mostly determined by the additive effects of genes with their heritability more than 0.9, which was easily utilized in breeding.
Aegilopstauschii; Long-head; Big-kernel; Genetic analysis
时间:2016-07-07
2016-01-08
2016-03-04
四川省青年科技基金项目(2015JQ0022);四川省财政创新能力提升工程项目(2013GXJS-004);国家自然科学基金项目(31401382);四川省青年科技创新研究团队项目(2014TD0014)
E-mail:469311148@qq.com
万洪深(E-mail:wanhongshen@126.com);李 俊(E-mail:lijunchd@126.com)
S512.9;S330
A
1009-1041(2016)07-0849-07
网络出版地址:http://www.cnki.net/kcms/detail/61.1359.S.20160707.1529.008.html
