普通小麦-黑麦草(Loliumperenne L.)易位系鉴定及其后代灌浆与产量性状分析
2016-12-15王掌军张文杰万佳颖白丽娟贾影影
王掌军,张文杰,万佳颖,白丽娟,贾影影,
刘 妍1,刘凤楼1,刘生祥1,张晓岗1
(1.宁夏大学农学院,宁夏银川 750021; 2.宁夏农林科学院作物研究所,宁夏银川 750001)
普通小麦-黑麦草(LoliumperenneL.)易位系鉴定及其后代灌浆与产量性状分析
王掌军1,张文杰2,万佳颖1,白丽娟1,贾影影1,
刘 妍1,刘凤楼1,刘生祥1,张晓岗1
(1.宁夏大学农学院,宁夏银川 750021; 2.宁夏农林科学院作物研究所,宁夏银川 750001)
为明确外源染色体或片段对小麦籽粒灌浆和产量性状的影响,以普通小麦宁春4号和黑麦草远缘杂交后代为材料,借助分子细胞学手段鉴定其BC1代染色体组成,并对其BC1F3代籽粒灌浆和产量性状进行了分析。结果表明,ND99株系出现两个顶端小片段易位类型的GISH杂交信号,为真杂种;ND99的BC1F3后代中分离出有芒(ND99A)和无芒(ND99B)两种类型,并且同一个分子标记3EST-185能够追踪这两种类型。宁春4号和易位系后代(ND99A、ND99B)籽粒的体积、鲜重、干重、水分均表现为先增后降的变化趋势;宁春4号和ND99A的灌浆速率呈双峰曲线变化,而ND99B 呈多峰曲线变化。宁春4号的灌浆时间早,快速增长期长,籽粒饱满指数和粒重最大;ND99A灌浆迟,持续期短,籽粒饱满指数最低,粒重仅次于宁春4号;ND99B灌浆的峰值出现较早、较大,灌浆持续期短,籽粒饱满指数居中,粒重最低。易位系后代的穗长、穗粒数和结实小穗数明显高于宁春4号,可能是影响其灌浆的关键因素之一。
普通小麦;黑麦草;易位系;GISH;分子标记;灌浆特性;产量性状
Ningxia Academy of Agricultural-Forestry Sciences,Yinchuan,Ningxia 750021,China)
由于长期人工选择和栽培,导致大量小麦优异基因丢失,栽培小麦的遗传基础日趋狭窄,遗传脆弱性逐渐增加[1]。而小麦近缘种蕴含丰富的优异基因资源,如抗病虫害、抗逆、高蛋白含量等,将其导入普通小麦,可以拓宽遗传变异范围[2]。但导入外源遗传物质后,通常会对某个性状产生有利影响,而对小麦的其他性状产生不利影响。因此,在育种实践中,不仅要注重外源基因是否能够改良某些性状,还应重视其对小麦其他性状的影响,特别是产量性状,这也是创制的很多携带有近缘物种染色体的小麦材料很少能应用于小麦育种实践中的原因[3-7]。
黑麦草(LoliumperenneL.,2n=2x=14)为禾本科黑麦草属下的一个种,有高产、营养价值高、抗寒性好、再生速度快等特点[8-9],在宁夏地区种植还表现为不易倒伏、发芽快的特性。已有研究通过远缘杂交将黑麦草优异基因导入其他物种中,温常龙等[10]总结了30多年黑麦草与羊茅属间杂种高产、优质、抗性强的优势;介绍了通过基因渗入、雄核发育和转基因手段改良牧草的方法;综述了黑麦草羊茅属间杂种基因组原位杂交、分子标记和遗传图谱建立、数量性状位点分析等分子遗传学的研究进展。王子平等[11]借助花粉管通道法将黑麦草 DNA 导入到水稻保持系中,发现了一份恢复系,并对该恢复系所含恢复基因独特的遗传行为进行了分析。作为小麦三级基因库,20世纪90年代,我国小麦育种家沈天民将黑麦草(母本)和豫麦2号(父本)进行远缘杂交,培育了高产、优质、多抗的小麦新品种906,此后,再未见以黑麦草为亲本通过远缘杂交培育出小麦新品种或对小麦主要性状进行遗传改良方面的报道。鉴于此,本研究以本课题组前期得到的普通小麦宁春4号与黑麦草杂交后代为材料,借助分子细胞遗传学手段鉴定BC1代外源染色体或片段的来源、身份,并对BC1F3代籽粒灌浆和产量性状进行分析,以期明确外源染色体或片段对小麦籽粒灌浆和产量性状的影响,为把近缘种属的优良基因导入普通小麦、拓宽遗传变异范围提供参考信息和种质资源。
1 材料与方法
1.1 材 料
供试材料包括普通小麦宁春4号、黑麦草、宁春4号与黑麦草杂交后回交(宁春4号为轮回亲本)BC1代及其F1代回交、自交BC1F3代,均由宁夏大学小麦育种课题组提供。2015年3月7日将宁春4号及其与黑麦草杂交后代BC1F3种植于宁夏大学教学实验农场,每个材料种植1个小区,每小区20行,行长1.1 m,行宽0.2 m,生育期管理同大田。
1.2 方 法
1.2.1 基因组原位杂交(Genomicinsituhybridization,GISH)分析
根尖细胞染色体制片参照Gill等[12]的方法,并稍作改动。即剪取2个长约1.0~1.5 cm的种子根尖,0 ℃冰水中处理22~24 h,用卡诺氏固定液(95%乙醇∶冰醋酸=3∶1)固定,在45%的醋酸中压片,相差显微镜下镜检。
GISH分析参照Jiang和Gill[13]的程序。将45%醋酸压片-70 ℃冰冻揭片,经70%、95%和100%乙醇梯度脱水后备用。所用探针为经缺口平移法用Fluoresceint-12-dUTP标记的黑麦草基因组DNA。经杂交、洗脱、信号放大及PI复染后,在Olympus BX60型荧光显微镜下用450~490 nm激发光波长观察,用SPOT CCD (SPOT Color Digital Camera)摄取图像。
1.2.2 分子标记分析
基因组DNA的提取采用SDS法[14]。根据定位于普通小麦中国春7个部分同源群的不同区段的EST序列(http://wheat. pw. usda. gov/ cgi-bin/westsql/map_locus. cgi)或与小麦相应部分同源群具有共线性关系的水稻和短柄草的测序结果设计引物124对,并交由北京奥科生物技术有限责任公司合成。PCR反应体系(10 μL):10× Reaction Buffer 1 μL,MgCl2(25 mmol·L-1)0.8 μL,dNTPs(2.5 mmol·L-1)0.8 μL,上下游引物(10 μmol·L-1)各0.2 μL,模板DNA 25 ng,TaqDNA聚合酶0.5 U,加ddH2O至总体积10 μL。反应条件:94 ℃预变性3 min;94 ℃变性30 s,55 ℃退火45 s,72 ℃延伸1 min,35个循环;72 ℃ 延伸10 min,4 ℃保存。扩增产物采用8%聚丙烯酰胺凝胶电泳检测,电泳时总电压为150 V,电泳1.5 h左右,经硝酸银染色后观察并照相。
1.2.3 籽粒灌浆和产量性状分析
2015年在小麦开花盛期准确标记同一时期开花、穗型一致的穗,每个材料标记150穗左右。从开花当天开始,每5 d于上午9 :00前从5个单株上取10个单穗,直至成熟。每次取样后籽粒人工剥取并放入烧杯,称鲜重;运用液面差法测量籽粒体积;105 ℃杀青1 h后,75 ℃烘干至恒重测定籽粒干重。籽粒水分=籽粒鲜重-籽粒干重。平均灌浆速率=(前一时间测定的籽粒干重-下一时间测定的籽粒干重)/间隔天数。这两个指标均用1 000个籽粒计算。
成熟期在1.1 m双行区内,调查30个有效穗的总小穗数、总粒数和结实小穗数,并称量每穗粒重,然后计算每穗平均粒数、粒重、不孕小穗数和结实小穗数。不孕小穗数=总小穗数-结实小穗数。籽粒饱满指数=成熟籽粒晒干体积/最大鲜籽粒体积。经济系数=经济产量/生物产量,其中,生物产量为收获后未脱粒前的植株(中途未取样的植株)重量,经济产量为脱粒后的籽粒重量。
1.2.4 数据处理
采用Microsoft Excel 2010处理数据,并作图。
2 结果与分析
2.1 小麦-黑麦草远缘杂交后代的GISH分析和分子标记鉴定结果
在普通小麦宁春4号与黑麦草杂交后收获到的11粒F1代种子中,大多数干瘪、发芽力很低,或者发芽不结实,仅有1粒种子长出后代并结实,用宁春4号回交,BC1代经GISH鉴定,所有个体的体细胞染色体数目均为2n=42,但多数没有检测到杂交信号,唯有编号为ND99个体的染色体顶端有GISH信号(图1),说明该个体中小麦与黑麦草发生了易位,且属于自发易位类型。
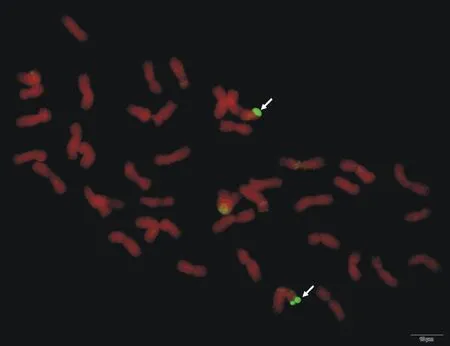
图中箭头所指绿色荧光信号为易位的黑麦草染色体;图示标尺为10 μm。
Chromosome fragments visualized with green fluorescence were fromLoliumperenne; Scale bar was 10 μm.
图1 ND99根尖细胞染色体的GISH分析
Fig.1 GISH analysis of ND99 root tip cell chromosome
以宁春4号和黑麦草的DNA为模板,利用已开发的124对 EST标记引物进行扩增,筛选到13对在宁春4号和黑麦草间有多态性的标记引物(表1),多态性比率为10.48%。利用这13对具有多态性的引物对宁春4号、黑麦草、ND99及ND99回交、自交后代中分离出的有芒和无芒株系(ND99A和ND99B)进行分析发现,定位于小麦染色体3AL、3BL、3DL上的标记3EST-185在黑麦草、ND99及ND99A和ND99B各15个随机取样单株中均扩增出相同的特异性条带,而宁春4号中并没有扩增到分子量相同的条带(图2)。进一步说明株系ND99为小麦与黑麦草之间发生了易位,而且标记3EST-185可以跟踪小麦背景中黑麦草染色体片段。
2.2 小麦-黑麦草易位系后代籽粒灌浆特性分析
2.2.1 籽粒形态指标的动态变化
宁夏灌区小麦基本上5月中下旬开花,5月底、6月初开始灌浆,7月中上旬小麦成熟,灌浆结束。本试验从花后第1天6月1日到7月15日,选取生育期一致的株系,每5d测量1次灌浆期籽粒体积、鲜重、干重和水分等指标。
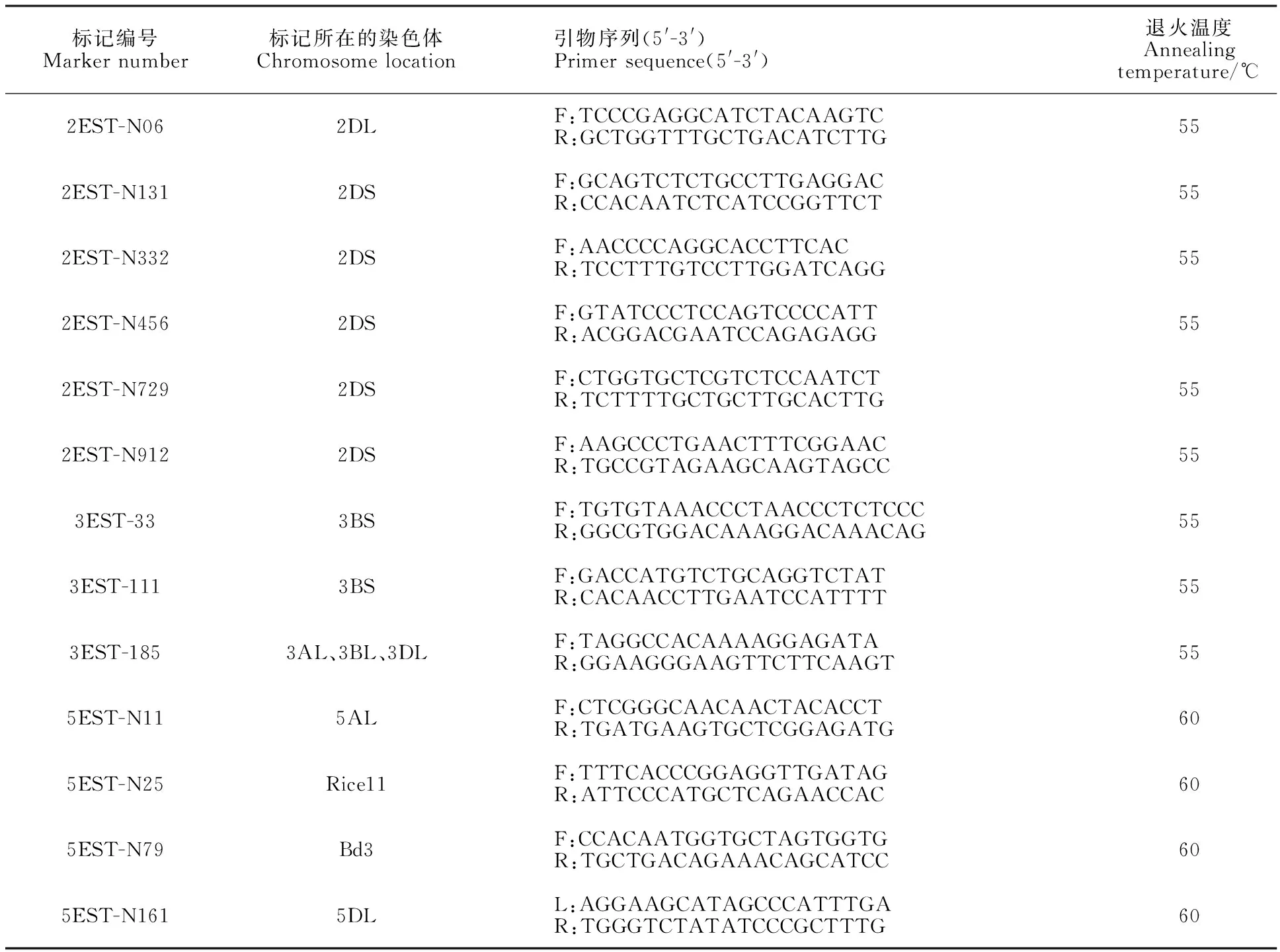
表1 本研究筛选到的黑麦草多态性引物
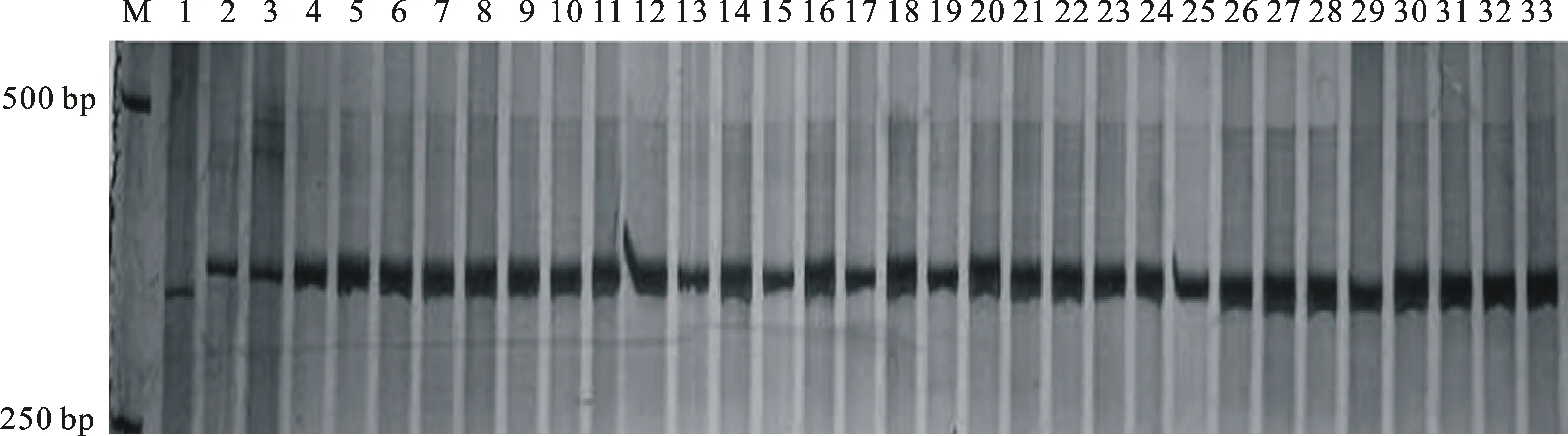
M:DL2000;1:宁春4号;2:黑麦草;3:ND99;4~18:ND99A的15个不同单株;19~33:ND99B的15个不同单株。
M:DL2000; 1:Ningchun 4; 2:Loliumperenne; 3:ND99; 4-18:Fifteen individual plants of ND99A; 19-33:Fifteen individual plants of ND99B.
图2 标记3EST-185在宁春4号、黑麦草及其二者远缘杂交后代中的扩增结果
Fig.2 Amplication of marker 3EST-185 in Ningchun 4,LoliumperenneL. and the two distant hybrid offsprings
就籽粒体积而言,宁春4号在花后1~5 d增长较ND99A和ND99B快,花后5~13 d快速增长,此后,增长较为缓慢,花后33 d达到最大(56.391 mL),花后33~41 d迅速降低,花后41~45 d又出现缓慢增长的趋势;ND99A在花后29~37 d,增长较宁春4号迅速,同在花后33 d达到最大(59.352 mL);ND99B在花后17~21 d和25~29 d增长最为迅速,花后29 d达到最大值(68.105 mL),花后29~45 d体积一直处于变小趋势(图3A)。
就籽粒鲜重而言,宁春4号、ND99A和ND99B三个材料在花后1~5 d,表现为稳中有升,之后,各自进入快速增长期;宁春4号和ND99A在花后33 d达到最大值(68.921 g和69.464 g),而ND99B花后29 d达到最大值(78.204 g),随后,鲜重下降,下降幅度大小顺序为:ND99A>ND99B>宁春4号(图3B)。
就籽粒干重而言,花后1~9 d,宁春4号、ND99A和ND99B均处于平稳增长期,增幅不大,此后迅速增加,宁春4号在花后9~29 d从6.973 g增加到38.062 g,表现为持续快速增加,持续时间最长,花后41 d达到最大值(47.267 g),随后下降,花后45 d降到43.787 g;ND99A在花后13 d进入持续快速增长期,花后37 d达到最大值(41.303 g);而ND99B分别在花后9~13、17~21、25~29 d有3个间断的快速增长期,在花后37 d达到最大值(44.822 g),从花后37 d开始,ND99A干重较ND99B大幅度下降(图3C)。
就籽粒水分而言,花后1~17 d,宁春4号远高于ND99A和ND99B,花后17 d达到最大值(30.939 g),之后宁春4号的明显低于其他两个材料;而ND99A和ND99B的最大值(33.442 g和43.063 g)分别出现在花后25 d和21 d,并且最大值均高于宁春4号;达到最大值后,均以不同幅度下降,宁春4号和ND99A花后41 d下降到最小值(4.715 g和3.747 g),而ND99B花后45 d下降到最小值(5.863 g)(图3D)。
2.2.2 籽粒灌浆速率的动态变化
由图4可知,在花后1~5 d,宁春4号千粒重每天增加0.4 g左右,而ND99A和ND99B平均每天增加分别为0.13和0.16 g;花后9~13 d,ND99B灌浆速率最快(1.61 g·d-1),宁春4号次之(1.18 g·d-1),ND99A最低(0.65 g·d-1);花后29~33 d,ND99A首次超过其他两个材料;花后17~21 d,宁春4号、ND99A和ND99B同时迎来第一个灌浆高峰期,花后25~29 d,3个材料迎来第二个灌浆高峰期,灌浆速率分别为1.75、2.02和2.63 g·d-1,此后,所有材料的灌浆速率呈下降趋势,花后37~41 d,宁春4号千粒重每天增加0.8g,而ND99A和ND99B的千粒重呈负增长;花后41~45 d,3个材料的千粒重增长均呈负增长。
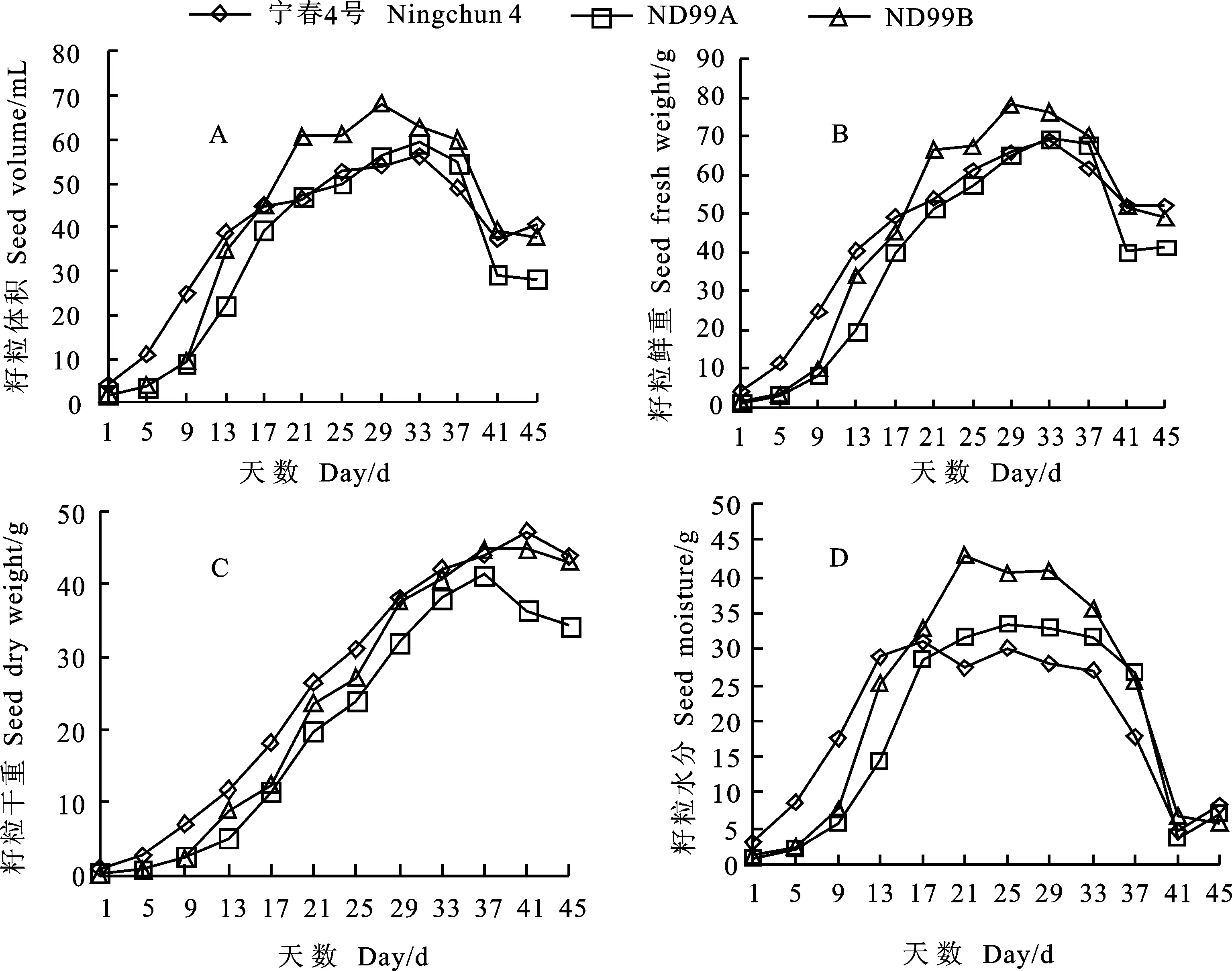
图3 亲本和小麦-黑麦草易位系后代灌浆期籽粒(1 000粒)不同形态指标的动态变化
2.3 小麦-黑麦草易位系后代产量性状分析
产量性状分析结果(表2)表明,ND99A和ND99B的穗长分别高出宁春4号32.33%和22.21%,穗粒数明显高于宁春4号,分别高出86.81%和88.70%,结实小穗数也分别高出42.86%和28.57%,因而易位系后代因为穗长、穗粒数较多和结实小穗数较高,形成较大库容,进而影响灌浆,导致其穗粒重和籽粒饱满指数较低;不孕小穗数为ND99A>宁春4号>ND99B;宁春4号的生物产量居中,而经济产量最高,所以,其经济系数为最高。总体上,易位系后代有效地提高了穗长、穗粒数、结实小穗数,但是籽粒饱满指数较低,穗粒重偏低,在生物产量相当的情况下,经济产量不高,导致其经济系数偏低。
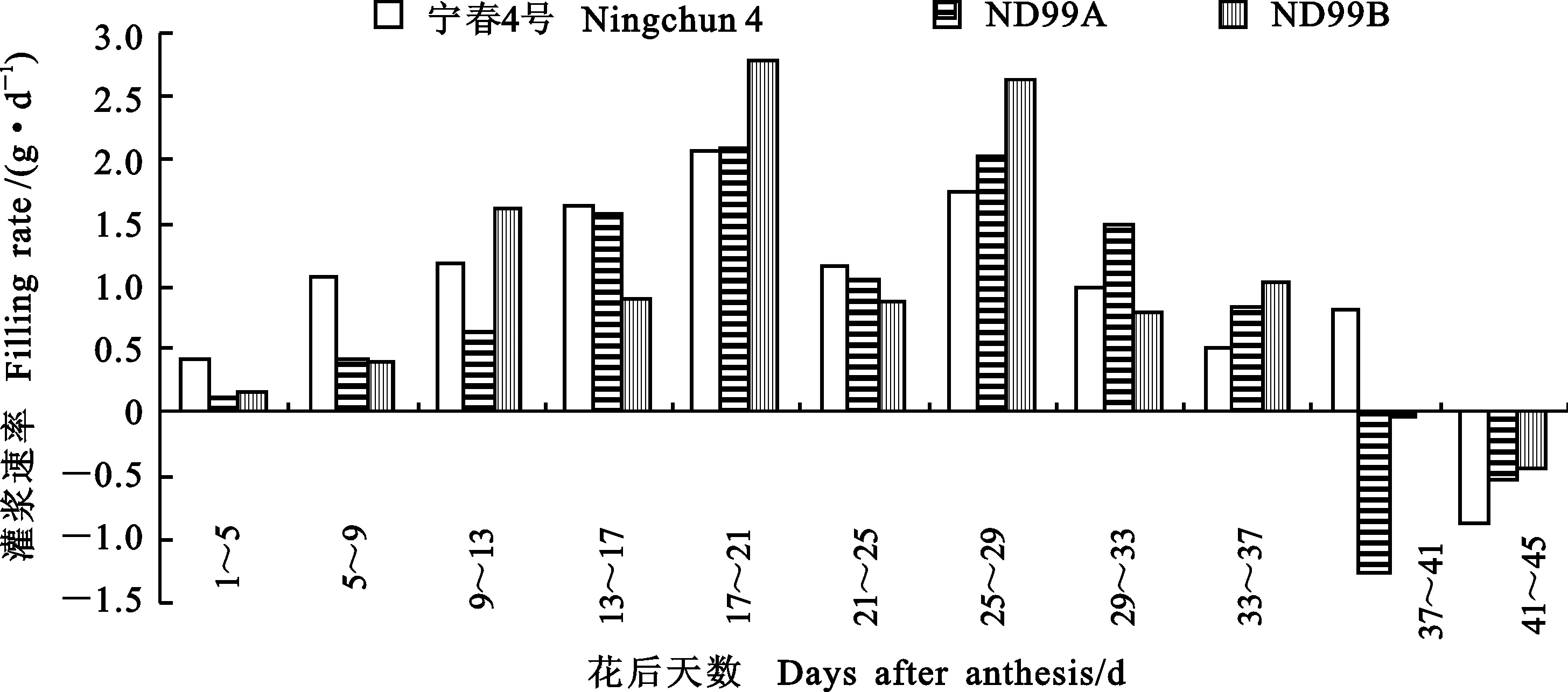
图4 亲本和小麦-黑麦草易位系后代籽粒灌浆速率的动态变化
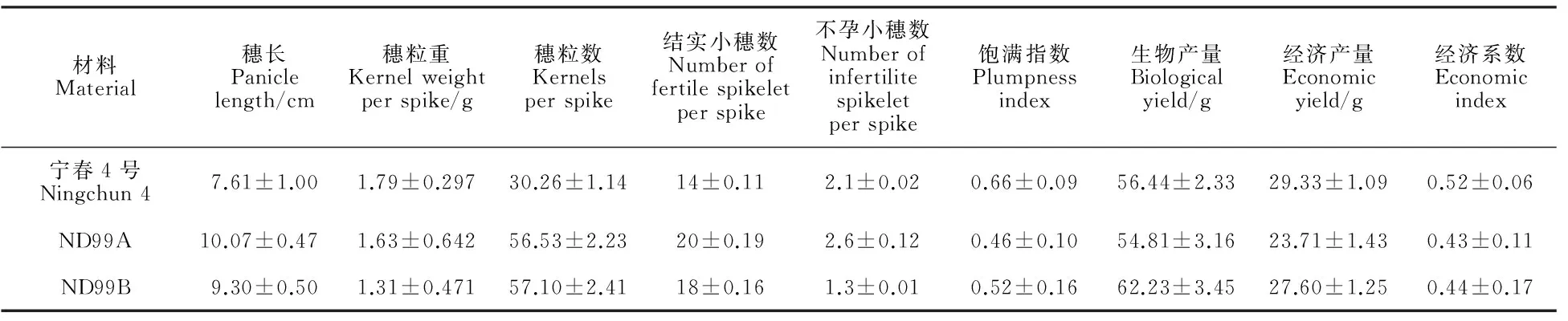
表2 亲本和小麦-黑麦草易位系后代产量性状测定结果
3 讨 论
黑麦草作为小麦遗传改良的三级基因库中的一个物种[15],因亲缘关系较远,与小麦杂交难度很大[16],而经一些科技工作者的不懈努力,已将外源物种一些优异基因导入小麦背景,在生长上发挥着巨大的作用[17-20];同时,利用GISH[13]和DNA分子标记技术[21-22],有效地鉴定小麦背景中外源染色体的来源、身份。本研究成功利用GISH鉴定了小麦-黑麦草易位系,并筛选到一个黑麦草的特异分子标记,可以看出,因外源染色体(片段)的导入,对小麦灌浆和产量性状有明显影响。该研究结果对于黑麦草基因组学研究和挖掘利用这类资源具有现实意义。
籽粒灌浆是产量形成的一个关键阶段,而小麦灌浆进程受基因型、生态因素(温度、干旱、地区性等)、生长调节物质、耕作栽培措施等综合因素的影响[23-28]。本试验中宁春4号和两个易位系后代(ND99A和ND99B)灌浆期籽粒的体积、鲜重、干重、水分均表现为先增后降的变化趋势。易位系后代的籽粒体积、鲜重、水分最大值均高于宁春4号,而最大籽粒干重却明显低于宁春4号。同时,宁春4号和ND99A的灌浆速率呈双峰曲线变化,而ND99B呈多峰曲线变化。其中,(1)籽粒体积增大是灌浆的一个重要标志,但是体积过大,后期籽粒能否饱满是一个问题。由于易位系后代的最大籽粒体积较大,可能影响到籽粒的饱满指数,造成籽粒皱缩加深,饱满程度下降。(2) 由于宁夏灌浆初期气温较低,影响到光合强度和物质运输,同一麦穗不同位置的小穗其发育通常不同步,中部小穗开花比两端小穗早;中部的籽粒为强势籽粒,生长迅速,进入灌浆快速期早,而两端的弱势籽粒灌浆开始生长缓慢,历经较长时间才进入灌浆快速期;中部成熟籽粒的粒重高于两端相应的成熟籽粒。易位系后代的穗长、穗粒数、结实小穗数均高于宁春4号,因为易位系后代属于强、弱势粒异步灌浆型[29],所以,粒重较低、不稳定,进而影响其产量。因此,在改良性状的同时还应加强栽培措施,促其稳产、高产。(3) 籽粒干重是小麦产量最直接的体现,灌浆速率为每天增加的籽粒干重。灌浆速率主要受遗传控制[30],与粒重显著正相关[31]。宁春4号籽粒干重比易位系后代持续快速增长时间长,而且干重达到最大值的时间较晚,而易位系后代的干物质积累量提前进入负增长,这主要受到当地温度等环境因子的调控,综合采用育种改良和栽培学措施,使其灌浆速度和灌浆时间尽可能协调。(4) 水是植物体完成生命活动所必须的,小麦籽粒不同时期含水量的多少对小麦灌浆影响很大,干物质运输需要水分,所以,灌浆前期干物质积累与含水量增加呈正相关,这有利于干物质的在小麦体内的积累与运输;而灌浆后期,易位系后代籽粒水分较高,种子晒干随着水分的大量蒸发,籽粒饱满指数较低,造成籽粒皱缩加深,饱满程度下降;而宁春4号籽粒水分较低,这正是其鲜重不高的情况下籽粒干重较高,饱满指数和穗粒重较高的原因所在。值得一提的是,易位系在穗长、穗粒数、结实小穗数等性状方面得到了改良,明显高于对照宁春4号,这可能是影响其灌浆的关键因素之一。今后工作中,对易位系后代的株型(比如矮秆株系选育)、合理的栽培密度、籽粒灌浆同步性等方面进行深入研究,以提高源库平衡程度,充分发挥其穗长粒多的优势,通过提高小麦单产保证总产;同时,加强外源基因对小麦品质、抗生物和非生物逆境等方面的潜能,以保证小麦可持续化生产。
[1]JIANG J M,GILL B S.Different species specific chromosome translocations inTriticumtimopheeviiandT.turgidumsupportthe diphyletic origin of polyploid wheats [J].ChromosomeResearch,1994,2(1): 59-64.
[2]SEARS E R.Transfer of Alien Genetic Material to Wheat [M].In:Evans L T,Peacock W J,eds.Wheat Science-Today and Tomorrow.Cambridge:Cambridge University Press,1981:75-89.
[3]陈佩度,周 波,齐莉莉,等.用分子原位杂交(GISH)鉴定小麦-簇毛麦双倍体、附加系、代换系和易位系 [J].遗传学报,1995,22(5):380-386.
CHEN P D,ZHOU B,QI L L,etal.Identification of wheat-Haynaldiaviilosaamphiploid,addition,substitution and translocation lines byinsituhybridization using biotin-labelled genomic DNA as a probe [J].ActaGeneticaSinica,1995,22(5):380-386.
[4]KISHII M,YAMADA T,SASAKUMA T,etal.Production of wheat-Leymusracemosuschromosome addition lines [J].TheoreticalandAppliedGenetics,2004,109:255-260.
[5]OLIVER R E,CAI X,XU S S,etal.Wheat alien species derivatives:a novel source of resistance to Fusarium head blight in wheat [J].CropScience,2005,45:1353-1360.
[6]SCHNEIDER A,MOLNR I,MOLNR-LNG M.Utilisation ofAegilops(goatgrass) species to widen the genetic diversity of cultivated wheat [J].Euphytica,2008,163:1-19.
[7]KANG H Y,ZHONG M Y,XIE Q,etal.Production and cytogenetics of trigeneric hybrid involvingTriticum,PsathyrostachysandSecale[J].GeneticResourcesandCropEvolution,2012,59:445-453.
[8]于玉红,王 亮,黄晓露,等.6个多年生黑麦草品种的交播性状比较 [J].草业科学,2009,26(8):172-176.
YU Y H,WANG L,HUANG X X,etal.Turf performance of six perennial ryegrass varieties used for overseeding [J].PrataculturalScience,2009,26(8):172-176.
[9]李杰勤,王丽华,詹秋文,等.2个黑麦草品种SPAD值和叶绿素及粗蛋白含量的相关性研究 [J].草业科学,2010,27(10):39-42.
LI J Q,WANG L H,ZHAN Q W,etal.The correlation study of SPAD value,chlorophyll content and crude protein content in two varieties ofLoliumperenne[J].PrataculturalScience,2010,27(10):39-42.
[10]温常龙,赵 冰,杜建材.黑麦草与羊茅属间杂种研究进展 [J].中国农业科学,2010,43(7):1346-1354.
WEN C L,ZHAO B,DU J C.Advances in research of festulolium intergeneric hybrids [J].ScientiaAgriculturaSinica,2010,43(7):1346-1354.
[11]王子平,阳标仁.黑麦草恢复基因在水稻中的遗传行为研究 [J].湖南农业科学,2013(13):7-9.
WANG Z P,YANG B R.Genetic action of ryegrass restoring gene in rice [J].HunanAgriculturalSciences,2011(13):7-9.[12]GILL B S,FRIEBE B,ENDO T R.Standard karyotype and nomenclature system for description of chromosome bands and structural aberration in wheat [J].Genome,1991,34:830-834.
[13]JIANG J M,GILL B S.Sequential chromosome banding andinsituhybridization analysis [J].Genome,1993,36:792-795.
[14]YANG Z J,LI G R,CHANG Z J,etal.Charaeterization of a partial amphiploid betweenTriticumaestivume cv.Chinese Spring andThinopyrumintermidumssp.Trichophorum[J].Euphytica,2006,149:11-17.
[15]DONG F G,SONG J,NAESS S K,etal.Development and applications of a set of chromosome-specific cytogenetic DNA markers in potato [J].TheoreticalandAppliedGenetics,2000,101:1001-1007.
[16]MULLAN D J,MIRZAGHADERI G,WALKER E,etal.Development of wheat-Lophopyrumelongatumrecombinant lines for enhanced sodium 'exclusion' during salinity stress [J].TheoreticalandAppliedGenetics,2009,119:1313-1323.
[17]郭光艳,李瑞芬,张敬原,等.小麦微卫星引物对多枝赖草基因组DNA扩增的研究 [J].华北农学报,2004(1):1-4.
GUO G Y,LI R F,ZHANG J Y,etal.Amplifying products fromLeymusmulticaulisusing wheat microsatellite prime pairs [J].ActaAgriculturaeBoreali-Sinica,2004(1):1-4.
[18]QI L L,PUMPHREY M O,FRIEBE B,etal.A novel Robertsonian translocation event leads to transfer of a stem rust resistance gene( Sr52) effective against race Ug99 fromDasypyrumvillosuminto bread wheat [J].TheoreticalandAppliedGenetics,2011,123:159-167.
[19]KLINDWORTH D L,NIU Z X,CHAO S M,etal.Introgression and characterization of a goatgrass gene for a high level of resistance to Ug99 stem rust in tetraploid wheat [J].GenesGenomesGenetics,2012,2:665-673.
[20]CHEN P D,YOU C F,HU Y,etal.Radition-induced translocations with reducedHaynaldiavillosachromatin at the Pm21 locus for powdery mildew resistance in wheat [J].MolecularBreeding,2013,31:477-484.
[21]ZHANG W,ZHANG R Q,FENG Y G,etal.Distribution of highly repeated DNA sequences inHaynaldiavillosumand its application in the identification of alien chromatin [J].ChineseScienceBulletin,2013,58:890-897.
[22]ZHAO R H,WANG H Y,JIA Q,etal.Development of EST-PCR markers for the chromosome 4V ofHaynaldiavillosaand their application in identification of 4V chromosome structural aberrants [J].JournalofIntegrativeAgriculture,2014,13(2):282-289.
[23]吕厚波,冯安荣,董 瑞,等.不同基因型小麦灌浆和抗倒伏性能与籽粒产量的关系 [J].西北农业学报,2015,24(6):27-33.
LÜ H B,FENG A R,DONG R,etal.Grain-filling characteristics and stem lodging resistance and their relationships with grain yields of different genotypic wheat [J].ActaAgriculturaeBoreali-occidentalisSinica,2015,24(6):27-33.
[24]孔治有,唐李军,张玉荣,等.Wx基因对灌浆期小麦叶片POD活性的影响 [J].湖北农业科学,2015,54(18):4401-4404.
KONG Z Y,TANG L J,ZHANG Y R,etal.Effect ofWxgenes on POD activity of wheat leaves at filling stage [J].HubeiAgriculturalSciences,2015,54(18):4401-4404.
[25]文廷刚,陈昱利,杜小凤,等.不同植物生长调节剂对小麦籽粒灌浆特性及粒重的影响 [J].麦类作物学报,2014,34(1):84-90.
WEN T G,CHEN Y L,DU X F,etal.Effects of different plant growth regulators on the grain filling characteristics and grain weight in wheat [J].JournalofTriticeaeCrops,2014,34(1):84-90.
[26]张卓亚,王晓琳,许晓明,等.腐植酸对小麦扬花期水分利用效率及灌浆进程的影响 [J].江苏农业学报,2015,31(4):725-731.
ZHANG Z Y,WANG X L,XU X M,etal.Effect of humic acid on water use efficiency and grouting process of wheat at flowering [J].JiangsuJournalofAgriculturalScience,2015,31(4):725-731.
[27]梁海燕,刘迪迪,张 娜,等.干旱胁迫对不同基因型小麦籽粒灌浆及内源激素的影响 [J].西北农业学报,2015,24(5):41-47.
LIANG H Y,LIU D D,ZHANG N,etal.Effect of water stress on grain filling and endogenous hormone changes in wheat grain of different genotypes [J].ActaAgriculturaeBoreali-occidentalisSinica,2015,24(5):41-47.
[28]张英华,杨佑明,曹 莲,等.灌浆期高温对小麦旗叶与非叶器官光合和抗氧化酶活性的影响 [J].作物学报,2015,41(1):136-144.
ZHANG Y H,YANG Y M,CAO L,etal.Effect of high temperature on photosynthetic capability and antioxidant enzyme activity of flag leaf and non-leaf organs in wheat [J].ActaAgronomicaSinica,2015,41(1):136-144.
[29]任红松,朱家辉,艾比布拉,等.小麦籽粒灌浆特性分析 [J].西北农林科技大学学报(自然科学版),2006,34(3):55-60.
REN H S,ZHU J H,AIBI B L,etal.Analysis on grain filling charaeteristics of wheat variety [J].JournalofNorthwestA&FUniversity(NaturalScienceEdition),2006,34(3):55-60.
[30]WIEGAND C I,CUELLAR J A.Duration of grain filling and kernel weight of wheat as effeeted by temperature [J].CropScience,1981,21(l): 95-101.
[31]冯素伟,胡铁柱,李 淦,等.不同小麦品种籽粒灌浆特性分析 [J].麦类作物学报,2009,29(4):643-646.
FENG S W,HU T Z,LI G,etal.Analysis on grain filling characteristics of different wheat varieties [J].JournalofTriticeaeCrops,2009,29(4):643-646.
Identification of Translocation Line ofTriticumaestivum-LoliumperenneL. and Analysis on Grain Filling and Yield Traits of Its Offsprings
WANG Zhangjun1,ZHANG Wenjie2,WAN Jiaying1,BAI Lijuan1,JIA Yingying1,LIU Yan1,LIU Fenglou1,LIU Shengxiang1,ZHANG Xiaogang1
(1.Agricultural College of Ningxia University,Yinchuan,Ningxia 750021,China; 2.Institute of Crop Sciences,
In order to explore the effect of exogenous chromosome or fragment on grain filling and yield of wheat,the offsprings of “Ningchun 4 ×Loliumperenne” distant hybridization were used as materials in this study. The chromosome composition of BC1hybrid offsprings was identified based on genomicinsituhybridization(GISH) usingLoliumperenneL. DNA as probe and molecular markers,and characteristics of grain filling and yield traits in BC1F3offsprings were analyzed. The results showed that,No. ND99 had two GISH signals of small fragment translocation types in top,and was true hybrid; The BC1F3offsprings of ND99 were divided into awned type(ND99A) and awnless type(ND99B),and a molecular marker 3EST-185 could identify the two types. The changes of grain volume,fresh weight,dry weight,and water at filling stage in variety Ningchun 4 and translocation offsprings(ND99A,ND99B) showed a trend of first increasing and then decreasing;The filling rate of Ningchun 4 and ND99A showed a double-peak curve,but ND99B showed many peaks.The filling stage of Ningchun 4 was early in time,with long rapid growth period and biggest plumpness index of grain and kernel weight per spike. The filling time of ND99A showed later,with short extending period and lowest plumpness index,and kernel weight per spike was lower than that of Ningchun 4. The filling peak of ND99B appeared earlier and bigger,with short extending period and lowest kernel weight per spike. In the yield traits,panicle length,kernel number and fertile spikelet number per spike of translocation lines were apparently higher than those of Ningchun 4,which could be one of the key factors affecting grain filling.
Triticumaestivum;LoliumperenneL.; Translocation line; GISH; Molecular marker; Filling characteristics; Yield trait
时间:2016-07-07
2016-01-31
2016-04-13
宁夏回族自治区农业育种专项(2013NYYZ0202)
E-mail:wangzj-gs@126.com
S512.1;S330
A
1009-1041(2016)07-0841-08
网络出版地址:http://www.cnki.net/kcms/detail/61.1359.S.20160707.1529.006.html
