重金属铅诱发机体氧化应激效应的研究进展
2016-12-01张浩刘汝涛魏云波
张浩+刘汝涛+魏云波
摘要:尽管各国政府已经采取多项措施限制和减少铅的使用,铅污染引发的健康问题依旧非常突出,仍是当前社会高度关注的环境污染与健康问题之一。研究表明,氧化应激是几乎所有疾病发生的诱因,而且铅类污染物对生命体的危害与氧化应激效应密切相关。铅暴露使机体产生过量的活性氧物质导致体内氧化还原平衡态被打破,造成蛋白质、核酸、脂类等生物大分子的氧化损伤,从而导致细胞凋亡或坏死和组织器官代谢紊乱,引发机体病变甚至癌症的发生。因此,有必要从生物大分子、细胞和实验动物三个层面系统综述铅暴露诱发机体氧化损伤的作用机理,阐述重金属铅诱发机体氧化应激效应的研究进展。
关键词:重金属铅;氧化应激;毒性效应
中图分类号:Q89 文献标识码:A
Abstract:Lead still possesses great threats to human health owing to its widespread distribution in the environment caused by human activities, although various actions have been taken to cut down the use and distribution of lead by the governments. Abundant evidence has indicated oxidative stress is a trigger of many varied diseases, which has still been one of the biggest concerns on environmental pollution and health. Multiple studies have shown that lead toxicity is related to oxidative stress because it generates reactive oxygen species (ROS), interferes with antioxidant enzyme activities, and breaks the balance of the pro-oxidant/antioxidant defense system, resulting in oxidative damage of proteins, nucleic acids and lipid compounds, cell apoptosis and necrosis, and metabolic disorders of tissues and organs of humans, and causes harmful diseases or cancers. Therefore, it is of great importance to review mechanisms of oxidative damage caused by lead combined at the molecular, cellular and organismal levels to understand harmful effects of lead exposure to human health.
Keywords: lead; oxidative stress; toxic effects
2015年11月下旬,国家环保部公布的《重金属污染综合防治“十二五”规划》2014年度考核结果显示,尽管国家重点监控的重金属污染物(铅、汞、镉、
铬和类金属砷)减排成效显著,但由于涉重金属产业的快速扩张造成重金属污染物排放总量仍处于高位水平,重金属环境风险隐患依然突出。根据美国地质调查局(United States Geological Survey)最新公布的2014年全球铅行业市场分析数据,中国依然是精铅产量和消费量最多的国家,均远远高于欧洲(第二位)和美国(第三位)之和。巨大的铅消耗量中仅有约1/4的铅被回收再利用,其余大部分以“三废”形式排入环境介质中,经食物链传递或呼吸道等途径进入人体并蓄积于肝脏、肾脏等靶器官中,进而侵入组织细胞诱发氧化应激、细胞凋亡等毒性作用损伤其生理功能。因此研究铅污染治理及铅污染引发的健康问题,已经成为环境污染与健康领域亟需解决的重要科学问题。本论文从动物水平、细胞水平和功能大分子水平系统综述铅暴露诱发氧化应激效应与机理的研究进展,为开展铅污染损伤人体健康的早期预警及防治技术提供借鉴。
1铅引发的毒性效应及与氧化应激的关系
1.1铅的毒性作用简介
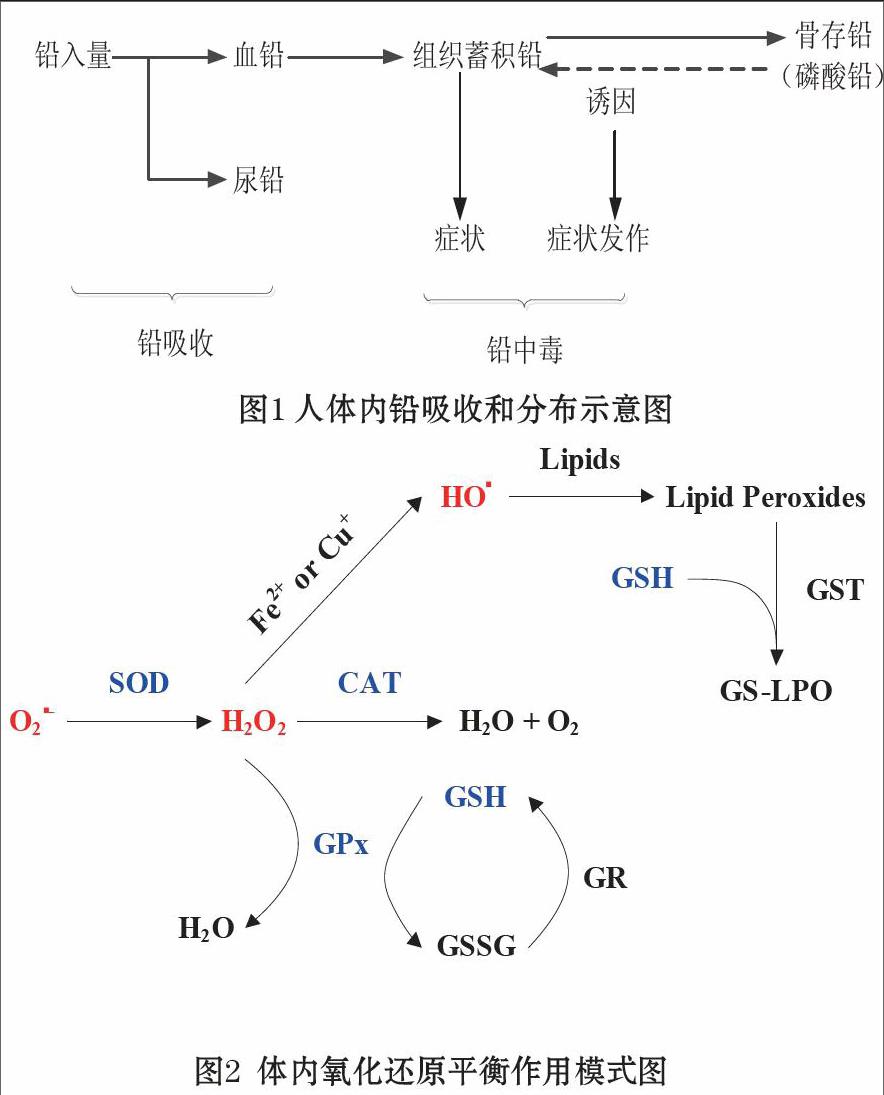
铅是地壳中含量最多的重金属元素。由于铅具有密度大,柔软性,耐腐蚀,延展性强等特点,在古代就有广泛应用。在古罗马时代就有铅制作的水管和酒器;我国在各个朝代都有使用铅制作的钱币。铅以工业规模生产和开发是19世纪才开始的,在19世纪中叶炼铅工业获得迅猛发展。据统计目前铅在有色金属生产中占第四位[2]。在工业开采、加工和制作过程中,铅会通过水、空气、食物链等途径与人类和动物体广泛接触,进而通过消化道、呼吸道和皮肤等吸收进入机体[3]。进入消化道的铅(吸收率仅为1%-2%,婴幼儿吸收率较高,钙缺乏可提高吸收率)主要在十二指肠吸收,经门静脉到达肝脏,一部分进入血液循环,一部分由胆汁分泌进入肠道,而后排出体外;进入呼吸道的铅25%-30%被吸收,粒径大于10 μm的含铅颗粒主要沉积于鼻腔和咽喉部,2.5μm以下者能够到达肺泡。铅吸收后进入血液,约96%迅速与红细胞结合,只有4%留在血浆中,然后经血液循环分布到肝、肾、脾、肺和脑等组织器官中[4],其中以肝脏和肾脏浓度最高[5],最终蓄积到骨骼、牙齿和毛发中,以磷酸铅的形式沉积下来[6],引发多种多系统性、多器官性的机体损伤[7]。铅的吸收和分布示意图见图1所示[2]。
1.2 氧化应激效应介绍
在正常的生理条件下,人体内存在着氧化活性物质和抗氧化防御体系的动态平衡,作用模式见图2所示。一旦体内氧化活性物质含量增加或者体内抗氧化防御体系被干扰和破坏,致使这种动态平衡产生紊乱和失调,引起一系列新陈代谢失常和免疫功能降低,形成氧自由基连锁反应而导致各组织发生氧化损伤,这就是氧化应激效应[8]。所说的氧化物质主要包括活性氧物质(Reactive Oxygen Species, ROS)和活性氮物质(Reactive Nitrogen Species, RNS),其中ROS是主要的氧化活性物质,主要包括过氧化氢(H2O2)、超氧阴离子(O2·-)和羟基自由基(HO·)等[9, 10]。正常条件下,人体内存在一定量的ROS,作为体内多种代谢和信号通路的信使,通过激活和调控各种转录因子参与体内多种基因的转录及相关功能蛋白的表达,参与细胞的增殖、分化及迁移及促进和维护细胞、组织和机体的新陈代谢[11, 12]。但是ROS具有较强的反应活性和氧化性,当ROS浓度较高时,会直接或间接对DNA、蛋白质及脂类物质发生氧化损伤,造成细胞凋亡(apoptosis)、衰老(senescence)和死亡(death)[13-16]。
大量体外和体内实验的研究表明,辐射和外源化学物(包括重金属类物质)是ROS介导的细胞损伤的主要环境诱导因子。电离辐射会通过氧化H2O产生HO·[17],皮肤受到紫外线(290-400 nm)照射会产生大量ROS,进而干扰了调控细胞增殖和分化的主要信号通路,影响了细胞的正常生理功能[18]。外源化学物如乙醇和苯巴比妥类物质通过改变细胞色素P450的途径,导致细胞色素P450的4A过氧化物酶体增殖,而导致了过氧化氢的产生[19]。重金属类污染物被认为是外源化学物中毒性较强的一类物质,它们发挥毒性效应的关键因素就是诱发机体产生氧化应激[20-23]。首先,重金属类物质可以参与类Fenton反应产生更多的ROS,是诱发氧化应激的直接因素[24];其次,重金属类物质易与体内抗氧化物质如还原型谷胱甘肽(GSH)发生共价结合而使其丧失消除ROS的能力[25, 26];另外,重金属类物质可以与抗氧化酶及谷胱甘肽相关酶发生相互作用,影响酶的活性和浓度,从而干扰体内的氧化还原反应[27-29]。后面这两种情况是重金属产生氧化损伤效应的非直接因素。
同时,体内存在着抗氧化防御系统,由抗氧化物质、抗氧化酶及非酶抗氧化蛋白组成[30]。抗氧化物质主要包括GSH、维生素C、维生素E、黄酮类物质、尿酸等;抗氧化酶主要包括超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、谷胱甘肽过氧化物酶(GPX)、谷胱甘肽还原酶(GR)、谷胱甘肽转硫酶(GST)及溶菌酶(Lysozyme)等;非酶抗氧化蛋白是指不直接参与分解代谢ROS物质,却能起到维持体内氧化还原平衡作用的蛋白,主要包括血红蛋白、肌红蛋白等含有铁/亚铁离子的传输蛋白、各种金属硫蛋白及人绒毛膜促性腺激素(HCG)等可以抑制氧化应激过程中细胞凋亡等损伤的功能蛋白[31-33]。体内抗氧化防御体系的存在,催化清除或分解代谢了过量产生的ROS,保障了各组织器官的正常生理功能。
1.3 铅引发的毒性效应与氧化应激的关系
E. D. Willis于1965年最先发现铅毒性与氧化应激相关[34]。铅的致病机理是较为复杂的,它可以与抗氧化酶发生直接作用[35-38],竞争性抑制重要微量矿物质的吸收,还能与体内含巯基物质发生共价结合使其失去抗氧化作用[22]。铅诱发的氧化损伤作用主要通过两种独立而又具有相关性的机理。首先,铅可以直接诱导机体产生单线态氧、过氧化氢及超氧化物等活性氧物质;第二种机制为铅能够消耗细胞内的抗氧化物质,其中铅影响GSH代谢途径是最主要的机理[39]。GSH是由半胱甘酸(Cys)、谷氨酸(Glu)和甘氨酸(Gly)组成的三肽,在淋巴细胞间质产生,是人体中最主要、含量最高的巯基类抗氧化物质[40]。GSH除了可以清除体内的活性氧之外,还可以与毒性重金属发生结合,削弱其毒性效应[41]。在铅暴露动物实验的研究中,血液、肝脏和肾脏等器官中GSH的浓度明显低于未暴露组[42-44]。
铅能够直接与含有巯基的抗氧化酶发生共价结合,使这些酶失活,从而破坏了氧化还原平衡状态。Ahamed等人报道在印度勒克瑙市一个铅工业暴露的地区,血铅水平为11.39 μg/dL左右的儿童,血液中羟基乙酰丙酸脱氢酶(ALAD)和GSH的含量明显低于血铅水平在7.11 μg/dL左右的儿童[35]。ALAD能够催化亚铁血红素(heme)的生成,从而维持血红蛋白的携氧能力,因此铅对ALAD活性的抑制作用是铅发挥毒性很关键的一个方面[45]。Hunaiti等发现在约旦伊尔比德市的从事铅行业工人的血液中,GR、GPX和GST的含量均与血铅水平呈反比,表明铅暴露使上述三种谷胱甘肽相关酶含量均发生下降,从而干扰了GSH代谢途径[46]。
目前,铅的毒性效应与氧化应激的相关性研究已有大量报道,因此本文将从动物、细胞和分子三个水平上分别综述铅暴露诱发氧化应激的研究进展,并从中找到尚未搞清楚的科研问题,作为后续工作的研究重点。
2 动物、细胞和分子水平上铅暴露诱发氧化应激的研究进展
2.1动物水平上铅暴露诱发氧化应激的研究进展
研究表明,长期铅暴露可导致肝脏、肾脏、生殖系统及神经系统的病变。对于铅毒性效应的研究大部分集中在流行病学领域,通过研究人群血铅水平、与身体各器官相关病变的生物学效应之间的关系来评价铅的毒性,为提出有效的治疗方案提供数据支持[47-49]。但是流行病学研究并不能实现铅暴露剂量变化与氧化应激相关生物学效应关系的研究,因此不能揭示铅诱发氧化应激的毒性作用机理。因此很多学者从动物水平上研究了铅毒性与氧化应激的关系。
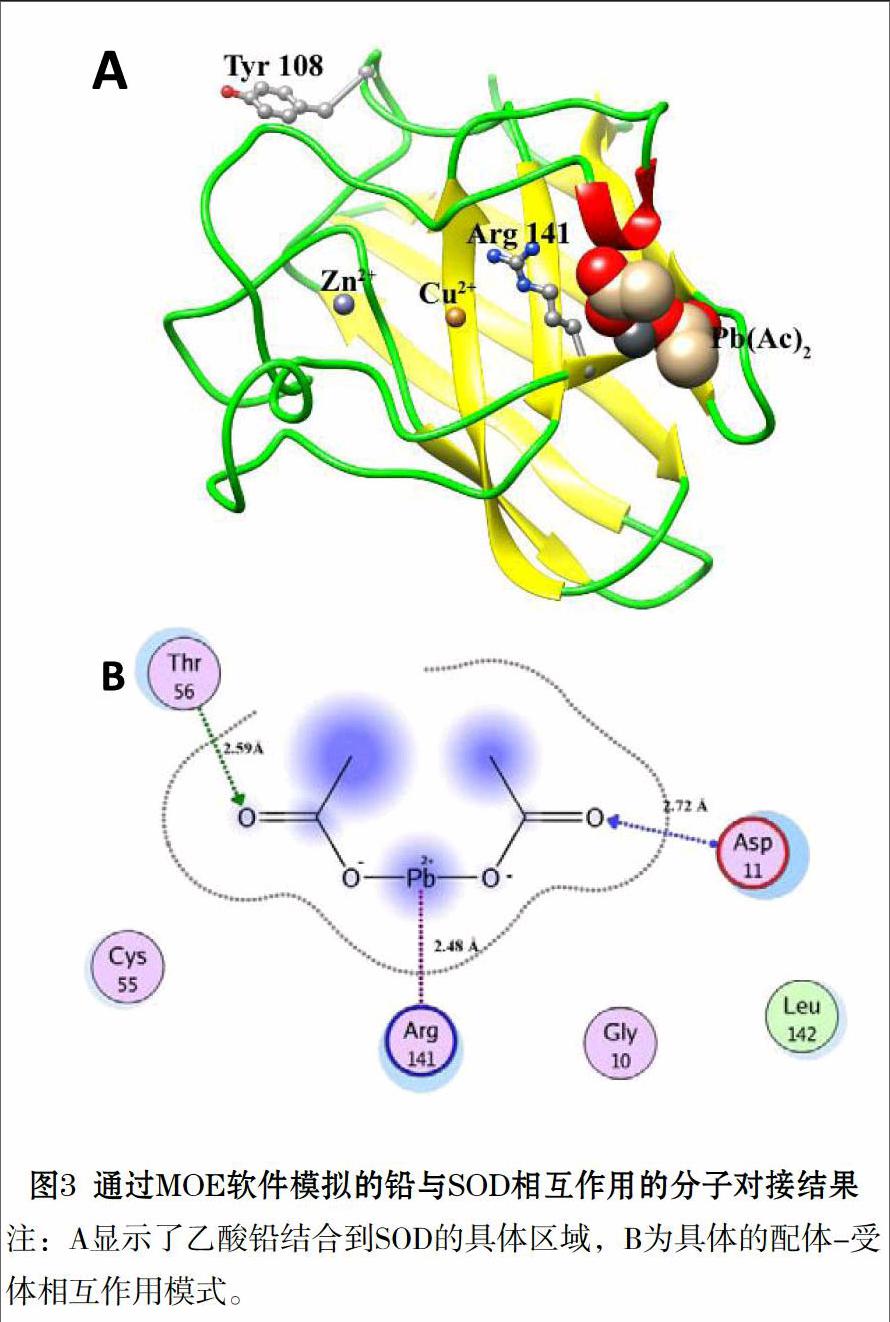
实验动物经铅暴露后,其抗氧化防御体系的变化情况是动物水平研究中主要关注的问题。Farmand等人研究了雄性Sprague-Dawley鼠经不同剂量铅染毒后主要抗氧化酶Cu/Zn SOD、CAT及GPx活性及含量的变化,结果显示铅暴露导致鼠胸动脉中Cu/Zn SOD活性升高,CAT和GPx活性没有变化,而且这三种酶的含量均未发生变化;在鼠肾脏和髓质中Cu/Zn SOD和CAT活性升高,而GPx活性没有变化。因此Farmand认为铅暴露后抗氧化酶活性升高是实验动物在氧化应激状态下的一种反馈抵抗机制[50]。但是Sivaprasad课题组的研究结果却与上述报道相反:他们在铅暴露后的小鼠血液红细胞中发现Cu/Zn SOD、CAT和GPx活性下降,因此解释为酶活性下降是铅诱发氧化应激的标志[51]。以上不同的实验结果表明,在不同的暴露剂量、暴露时间和靶器官等实验条件下,铅暴露过程中抗氧化酶活性变化规律不具有确定性,而且目前的研究并未阐明由于铅暴露导致酶活性的变化,引起体内氧化还原状态的失衡造成氧化损伤的发生;还是铅暴露诱发机体氧化损伤,导致了酶活性变化[52]。针对这一问题,我们研究了斑马鱼水平上铅暴露诱发氧化应激过程中SOD活性变化的机理:铅暴露打破了斑马鱼肝脏中的氧化还原稳态,导致SOD酶活性下降,谷胱甘肽相关酶GPx、GR和GST活性受到抑制, GSH/GSSG比率下降,并对细胞膜结构产生了氧化损伤,导致脂质过氧化物MDA含量增加;然后利用多种光谱学方法、ITC法及分子对接模拟等方法从生物大分子层面上深入探讨了铅暴露诱发斑马鱼氧化应激过程中CAT活性降低的机理。研究发现铅通过静电作用(ΔH<0, ΔS>0)与酶催化关键氨基酸残基Arg 141发生了直接作用,使铅结合到SOD的活性通道内,阻碍了底物(O2-·)进入酶活性中心的路径,破坏了SOD的骨架结构和二级结构,并使活性中心的Cu2+和Zn2+释放出来,从而导致了SOD活性的下降[53]。
当铅暴露导致机体各组织氧化损伤程度较深时,产生的ROS会进攻膜结构中的磷脂层,产生脂质过氧化反应[31]。脂质过氧化程度取决于膜结构中脂肪酸的饱和程度,不饱和程度越高(不饱和键越多),越容易发生脂质过氧化反应。脂质过氧化将使细胞膜的流动性和通透性改变,导致细胞结构和功能的异变,最终导致多器官损伤的发生[54]。Gerber和Rehman发现在铅暴露的小鼠大脑匀浆液中脂质过氧化物的产生量随着铅染毒浓度的升高而增多[55, 56]。而且Rehman还研究了小鼠大脑不同区域铅的含量,他发现铅富集量大的区域,脂质过氧化程度高[57]。Adanaylo和Oteiza 也证实铅暴露干扰了小鼠大脑中抗氧化防御体系,作为脂质过氧化的生物标志物MDA含量升高[58]。刘芳丽等人在铅暴露后的小鼠肝脏和肾脏中均发现了MDA含量的升高,且加入抗氧化物质白藜芦醇后减轻了铅的脂质过氧化水平[59]。
2.2细胞水平上研究铅诱发氧化应激的研究进展
动物水平的研究结果分析了铅暴露引发毒性效应与氧化应激的相关性,阐明了铅暴露导致组织器官损伤及引发各种疾病的重要机理是铅破坏了体内氧化还原状态,是机体发生氧化损伤所致。但是动物水平上的铅毒性评价并不能解释铅暴露诱发细胞毒性的微观机制,也就无法研究细胞内发生氧化应激作用机理。因此,很多研究者开始从细胞水平上研究铅诱发氧化应激的机理及其与细胞凋亡等细胞毒性的关系。
已有研究表明铅毒性引起细胞损伤与细胞内氧化还原状态异常有关。Yedjou和Tchounwou研究发现铅引起HepG2细胞活力下降与细胞内ROS含量升高有关,同时铅暴露引起细胞内MDA含量显著升高;当铅暴露同时加入抗氧化物质N-乙酰-L-半胱氨酸(NAC),会使细胞内ROS含量显著降低,而且显著提高了细胞成活率。表明NAC抑制了铅暴露引发细胞产生的氧化应激效应[60]。Stacchiotti等人选取小鼠肾脏近曲小管细胞(NRK-52E)为靶细胞进行铅暴露后,引起了细胞凋亡和坏死、线粒体损伤等细胞损伤作用,而且伴随着细胞内ROS和RNS含量的升高[61]。Navarro-Moreno等人同样在小鼠肾脏近曲小管细胞中发现了细胞损伤与氧化影响的相关性[62],评价了铅对PC 12细胞的细胞毒性。Jadhav等发现低浓度铅急性暴露(0.01 μM,24 h)可激活细胞内蛋白激酶C(PKC)的活性,PKC的活化调节了钙传输相关的NMDA受体通道,从而提高了细胞内Ca2+含量;而NMDA受体激活将促进兴奋型氨基酸谷氨酸盐的释放,进而引发细胞内ROS含量上升,产生氧化应激效应。他们进一步研究了铅与谷氨酸盐对PC12细胞的联合暴露效应,发现细胞内ROS含量大幅升高,细胞成活率显著降低,表明铅能够通过调节PCK活性进而引发细胞产生氧化损伤[63]。
在铅暴露诱发细胞氧化损伤的过程中,细胞内有着“动力车间”之称的线粒体起着至关重要的调节作用。线粒体的功能与ROS的产生及代谢密切相关,一方面线粒体在氧化代谢制造ATP过程中会伴随大量ROS的产生,在正常生理条件下其生成速率与线粒体膜电位水平(MMP)有直接关系[64]。Suski等人证实线粒体膜电位降低直接导致ROS的产生量增大[65];另一方面线粒体内的抗氧化防御体系能够有效消除过量的ROS,使自身免受氧化损伤而继续发挥其重要生理功能[66]。当细胞内氧化应激程度较严重时,过量蓄积的ROS可氧化渗透性转运通道上的敏感位点,会导致线粒体发生形态肿胀、膜电位降低等氧化损伤效应,产生严重的细胞毒性[67]。同时,线粒体在细胞凋亡过程中发挥重要作用。Oluwole等人于1998年在science杂质上详细分析了线粒体参与细胞凋亡的相关机理:(1)线粒体功能的紊乱导致细胞凋亡的产生,包括电子传递、氧化磷酸化和ATP产量的紊乱;(2)线粒体通过释放细胞色素C等信号因子激活Caspase家族,从而导致细胞凋亡的产生;(3)线粒体内氧化还原电位的改变,导致MMP值变化及调控Bcl-2家族的磷酸化水平,从而达到调控细胞凋亡的目的[68]。王林等人研究了铅暴露对大鼠肾小管上皮细胞MMP值与ROS含量的关系,发现铅染毒使肾细胞内ROS大量产生,且伴随着MMP数值下降。这是由于蓄积的ROS会直接靶性损伤线粒体膜结构,引起MMP值下降,进而导致了细胞凋亡[69]。徐进等人研究发现铅暴露可导致PC 12细胞DNA损伤及细胞凋亡,同时伴随着凋亡信号通路Bax表达量上升、Bcl-2表达量下降、P53表达量增加且Caspase-3被激活。表明铅诱发DNA损伤与P53表达量上调有关,导致Bax/Bcl-2状态失衡和线粒体损伤,从而激活了Caspase-3,进一步引起了细胞凋亡[70]。
2.3 分子水平上研究铅诱发氧化应激的研究进展
研究表明,重金属引发生物体毒性效应是从生物大分子开始的,比如各种酶、非酶功能性蛋白、DNA和RNA等,然后逐步在细胞层面、组织器官层面和个体层面反映出来[71],因此研究重金属与生物大分子的相互作用,能从分子层面上解释重金属引发毒性效应的机理,为与重金属相关疾病的早期诊断及治疗提供依据。但目前从分子层面上研究铅与生物大分子的相互作用研究很少。Belatik等人利用FTIR、紫外可见吸收光谱、圆二色谱、荧光光谱仪和X射线光电子能谱等技术从分子水平上对比研究了铅(II)与HSA与BSA的结合模式。Vega等人通过研究铅与人鱼精蛋白(HP2)的直接作用来解释铅的生殖毒性。他们通过多种光谱学手段研究了铅对HP2结构的影响,结果显示铅结合到HP2的两个结合位点处,从而破坏了HP2的生理结构,并认为由于HP2对铅和锌的亲和力类似,因此铅暴露会竞争性抑制锌与HP2的结合从而引发生殖毒性效应。
但分子水平的研究中存在一些方法学问题。首先上述研究中都未考虑荧光内滤效应对体系荧光变化的影响。荧光内滤效应是指溶液在激发波长和发射波长处的光发生吸收或色散而导致荧光信号减弱的现象。针对这一问题,我们课题组已经提出利用内滤校正公式和外滤校正装置来消除内滤效应影响的方法[72, 73],但是内滤校正公式的适用性有待进一步研究。因此最好的校正内滤的方法应该是避免内滤效应的产生[74]。一般认为,让溶液在激发波长到扫描的发射波长范围内的吸光度小于0.1时,可以忽略内滤效应的影响。由于没有校正内滤效应,利用Stern-Volmer方程判别荧光猝灭机理的类型就存在问题。事实上,最准确的荧光猝灭类型判别方法是考察反应体系荧光寿命的变化[75]。另一个问题是利用双对数Stern-Volmer方程 计算结合位点数和结合常数,并进一步通过热力学常数的计算确定结合作用力类型。但这个方程当且仅当配体与受体的结合属于无限协同反应(infinite cooperativity),受体的结合位点全空或者全满)时才能用来计算结合常数及结合位点数[75]。近几年等温滴定微量热技术(ITC)的发展为研究配体与受体相互作用过程的结合常数、结合位点数及作用力类型成为可能[76, 77]。针对上述问题,我们利用荧光分析法、紫外可见吸收光谱、圆二色谱、等温滴定微量热技术、分子对接模拟等方法,并考虑内滤效应对反应体系的干扰,分别研究了铅与SOD、溶菌酶、人绒毛膜促性腺激素以及DNA的相互作用机理,纠正了铅与生物大分子相互作用研究中的方法学问题[53, 78-80]。
分子水平上研究铅的氧化应激效应,就是要通过研究铅与氧化应激相关酶的直接作用,阐明酶活性变化的分子机理,揭示氧化应激过程中细胞内活性氧水平发生变化的原因,从而更加准确解释铅暴露诱发氧化应激引发的相关病变,为开发有效拮抗铅毒性、保障健康的治疗方案提供实验数据支持。目前从分子水平上研究铅与氧化应激相关酶的直接作用机理研究较少。前文已经提及,我们从斑马鱼水平研究了铅诱发氧化应激过程中SOD活性降低的分子机理,并对比分析了SOD在体内和体外铅暴露实验后酶活性变化的相关性。该研究为从分子水平上准确评价重金属铅对氧化应激相关酶的毒性效应机理提供了方法学参考。
3. 展望
根据世界卫生组织(WHO)2002年的世界健康报告,铅暴露是威胁人类健康的全球前20位疾病风险因素之一。目前很多科研工作者关注新型环境污染物(如纳米材料、阻燃剂等)的毒理学研究,但包括重金属铅在内的毒性更强的传统持久性有毒环境污染物,其引发的健康问题更需值得关注,而且在铅的毒理学研究中仍然存在很多尚未阐明的基础科学问题,需要进行深入的研究。
重金属铅通过各种途径进入生物体后,会使生物体内产生过量的活性氧物质,导致机体发生氧化应激效应,造成蛋白质、核酸、脂类等生物大分子的氧化损伤,引发细胞凋亡、各功能脏器代谢紊乱,最终导致疾病的发生,严重时将引发肿瘤性病变。还会与生物大分子(各种酶、非酶蛋白及核酸)发生直接相互作用,造成其功能的衰退或丧失,并最终逐步在细胞水平和机体水平反映出来。同时,不同暴露途径下铅的毒性效应是有较大差别的,即使控制铅的剂量相同,对同一生物学效应的评价结果仍可能不一致。目前并没有在不同暴露途径下对同一生物学效应的相关性研究。因此在明确动物水平、细胞水平和分子水平上铅诱发氧化应激效应机理以后,建立对同一生物大分子(蛋白质、DNA)在动物、细胞和分子水平上的结构和功能变化的相关性研究和评价方法,对于系统研究铅的氧化应激效应至关重要。
参考文献
[1] Salazar, K. Mineral Commodity Summaries 2013: US Geological Survey (USGS)[J]. US Geological Survey, 2013.
[2] 贾振邦. 环境与健康[M]. 北京: 北京大学出版社, 2008.
[3] Patrick, L. Lead toxicity, a review of the literature. Part 1: Exposure, evaluation, and treatment[J]. Alternative medicine review : a journal of clinical therapeutic, 2006, 11 (1): 2-22.
[4] Gillis, B. S.,Arbieva, Z.,Gavin, I. M. Analysis of lead toxicity in human cells[J]. BMC genomics, 2012, 13(1): 1-12.
[5] Oktem, F.,Arslan, M. K.,Dundar, B., etc. Renal effects and erythrocyte oxidative stress in long-term low-level lead-exposed adolescent workers in auto repair workshops[J]. Archives of toxicology, 2004, 78 (12): 681-687.
[6] Fontanellas, A., Navarro, S., Morán-Jiménez, M.-J., etc. Erythrocyte aminolevulinate dehydratase activity as a lead marker in patients with chronic renal failure[J]. American Journal of Kidney Diseases, 2002, 40 (1): 43-50.
[7] Patrick, L. Lead toxicity part II: the role of free radical damage and the use of antioxidants in the pathology and treatment of lead toxicity[J]. Alternative medicine review : a journal of clinical therapeutic, 2006, 11 (2): 114-127.
[8] Environmental stressors in health and disease[M]. New York: CRC Press, 2001.
[9] Balaban, R. S.,Nemoto, S.,Finkel, T. Mitochondria, oxidants, and aging[J]. Cell, 2005, 120 (4): 483-495.
[10] Lushchak, V. I. Free radicals, reactive oxygen species, oxidative stress and its classification[J]. Chemico-biological interactions, 2014, 224C 164-175.
[11] Reuter, S.,Gupta, S. C.,Chaturvedi, M. M., etc. Oxidative stress, inflammation, and cancer: how are they linked?[J]. Free radical biology & medicine, 2010, 49 (11): 1603-1616.
[12] Suzuki, Y. J.,Forman, H. J.,Sevanian, A. Oxidants as Stimulators of Signal Transduction[J]. Free Radical Biology and Medicine, 1997, 22 269–285.
[13] Alvarez-Gonzalez, R. Free radicals, oxidative stress, and DNA metabolism in human cancer[J]. Cancer investigation, 1999, 17 (5): 376-377.
[14] Folkes, L. K.,Christlieb, M.,Madej, E., etc. Oxidative metabolism of combretastatin A-1 produces quinone intermediates with the potential to bind to nucleophiles and to enhance oxidative stress via free radicals[J]. Chemical research in toxicology, 2007, 20 (12): 1885-1894.
[15] Valko, M.,Rhodes, C. J.,Moncol, J., etc. Free radicals, metals and antioxidants in oxidative stress-induced cancer[J]. Chemico-biological interactions, 2006, 160 (1): 1-40.
[16] Upadhyay, D.,Panduri, V.,Ghio, A., etc. Particulate matter induces alveolar epithelial cell DNA damage and apoptosis: role of free radicals and the mitochondria[J]. American journal of respiratory cell and molecular biology, 2003, 29 (2): 180-187.
[17] Yamaguchi, H.,Uchihori, Y.,Yasuda, N., etc. Estimation of yields of OH radicals in water irradiated by ionizing radiation[J]. Journal of radiation research, 2005, 46 (3): 333-341.
[18] Sakurai, H.,Yasui, H.,Yamada, Y., etc. Detection of reactive oxygen species in the skin of live mice and rats exposed to UVA light: a research review on chemiluminescence and trials for UVA protection[J]. Photochemical & photobiological sciences : Official journal of the European Photochemistry Association and the European Society for Photobiology, 2005, 4 (9): 715-720.
[19] Ziech, D.,Franco, R.,Georgakilas, A. G., etc. The role of reactive oxygen species and oxidative stress in environmental carcinogenesis and biomarker development[J]. Chemico-biological interactions, 2010, 188 (2): 334-339.
[20] Zhang, W.,Tan, N. G.,Fu, B., etc. Metallomics and NMR-based metabolomics of Chlorella sp. reveal the synergistic role of copper and cadmium in multi-metal toxicity and oxidative stress[J]. Metallomics : integrated biometal science, 2015, 7 (3): 426-438.
[21] Lee, J. C.,Son, Y. O.,Pratheeshkumar, P., etc. Oxidative stress and metal carcinogenesis[J]. Free radical biology & medicine, 2012, 53 (4): 742-757.
[22] Jomova, K.,Valko, M. Advances in metal-induced oxidative stress and human disease[J]. Toxicology, 2011, 283 (2-3): 65-87.
[23] Galanis, A.,Karapetsas, A.,Sandaltzopoulos, R. Metal-induced carcinogenesis, oxidative stress and hypoxia signalling[J]. Mutation research, 2009, 674 (1-2): 31-35.
[24] Liochev, S. I. The mechanism of "Fenton-like" reactions and their importance for biological systems. A biologists view[J]. Met Ions Biol Syst, 1999, 36 (4): 1-39.
[25] Jozefczak, M.,Remans, T.,Vangronsveld, J., etc. Glutathione is a key player in metal-induced oxidative stress defenses[J]. International journal of molecular sciences, 2012, 13 (3): 3145-3175.
[26] Potter, A. J.,Trappetti, C.,Paton, J. C. Streptococcus pneumoniae uses glutathione to defend against oxidative stress and metal ion toxicity[J]. Journal of bacteriology, 2012, 194 (22): 6248-6254.
[27] Barata, C.,Lekumberri, I.,Vila-Escale, M., etc. Trace metal concentration, antioxidant enzyme activities and susceptibility to oxidative stress in the tricoptera larvae Hydropsyche exocellata from the Llobregat river basin (NE Spain)[J]. Aquatic toxicology, 2005, 74 (1): 3-19.
[28] Vigneshkumar, B.,Pandian, S. K.,Balamurugan, K. Catalase activity and innate immune response of Caenorhabditis elegans against the heavy metal toxin lead[J]. Environmental toxicology, 2013, 28 (6): 313-321.
[29] Choudhary, M.,Jetley, U. K.,Khan, M. A., etc. Effect of heavy metal stress on proline, malondialdehyde, and superoxide dismutase activity in the cyanobacterium Spirulina platensis-S5[J]. Ecotoxicology and environmental safety, 2007, 66 (2): 204-209.
[30] Halliwell, B.,Gutteridge, J. M. C.,Halliwell, B., etc. Free radicals in biology and medicine[J]. Oxford University Press, 1999.
[31] 海春旭. 自由基医学[M]. 第四军医大学出版社, 2006.
[32] Liu, H.,Zheng, F.,Cao, Q., etc. Amelioration of oxidant stress by the defensin lysozyme[J]. American journal of physiology. Endocrinology and metabolism, 2006, 290 (5): 824-832.
[33] Kajihara, T.,Uchino, S.,Suzuki, M., etc. Human chorionic gonadotropin confers resistance to oxidative stress-induced apoptosis in decidualizing human endometrial stromal cells[J]. Fertility and sterility, 2011, 95 (4): 1302-1307.
[34] Wills, E. Mechanisms of lipid peroxide formation in tissues role of metals and haematin proteins in the catalysis of the oxidation of unsaturated fatty acids[J]. Biochimica et Biophysica Acta (BBA)-Lipids and Lipid Metabolism, 1965, 98 (2): 238-251.
[35] Ahamed, M.,Verma, S.,Kumar, A., etc. Environmental exposure to lead and its correlation with biochemical indices in children[J]. The Science of the total environment, 2005, 346 (1-3): 48-55.
[36] Gurer-Orhan, H.,Sab?r, H. U.,?zgüne?, H. Correlation between clinical indicators of lead poisoning and oxidative stress parameters in controls and lead-exposed workers[J]. Toxicology, 2004, 195 (2-3): 147–154.
[37] Lachant, N. A.,Tomoda, A.,Tanaka, K. R. Inhibition of the pentose phosphate shunt by lead: a potential mechanism for hemolysis in lead poisoning[J]. Blood, 1984, 63 (3): 518-524.
[38] Sandhir, R.,Julka, D.,Gill, K. D. Lipoperoxidative damage on lead exposure in rat brain and its implications on membrane bound enzymes[J]. Pharmacology & toxicology, 1994, 74 (2): 66-71.
[39] Ercal, N.,Gurer-Orhan, H.,Aykin-Burns, N. Toxic metals and oxidative stress part I: mechanisms involved in metal-induced oxidative damage[J]. Current topics in medicinal chemistry, 2001, 1 (6): 529-539.
[40] Sies, H. Glutathione and its role in cellular functions[J]. Free radical biology & medicine, 1999, 27 (9-10): 916-921.
[41] Jozefczak, M.,Remans, T.,Vangronsveld, J., etc. Glutathione is a key player in metal-induced oxidative stress defenses[J]. International journal of molecular sciences, 2012, 13 (3): 3145-3175.
[42] Garcon, G.,Leleu, B.,Zerimech, F., etc. Biologic markers of oxidative stress and nephrotoxicity as studied in biomonitoring of adverse effects of occupational exposure to lead and cadmium[J]. Journal of occupational and environmental medicine / American College of Occupational and Environmental Medicine, 2004, 46 (11): 1180-1186.
[43] CM, L.,GH, Z.,QL, M., etc. Protective effect of quercetin on lead-induced oxidative stress and endoplasmic reticulum stress in rat liver via the IRE1/JNK and PI3K/Akt pathway[J]. Free radical research, 2013, 47 (3): 192-201.
[44] ND, V.,CY, L.,F, F., etc. Superoxide dismutase, catalase, glutathione peroxidase and NADPH oxidase in lead-induced hypertension[J]. Kidney International, 2003, 63 (1): 186–194.
[45] Bergdahl, I. A.,Grubb, A.,Schütz, A., etc. Lead binding to delta-aminolevulinic acid dehydratase (ALAD) in human erythrocytes[J]. Pharmacology & Toxicology, 1997, 81 (4): 153-158.
[46] AA, H.,M, S. Effect of lead concentration on the level of glutathione, glutathione S-transferase, reductase and peroxidase in human blood[J]. Science of The Total Environment, 2000, 248 (1): 45–50.
[47] Toscano, C. D.,Guilarte, T. R. Lead neurotoxicity: from exposure to molecular effects[J]. Brain research. Brain research reviews, 2005, 49 (3): 529-554.
[48] Hu, H.,Shih, R.,Rothenberg, S., etc. The Epidemiology of Lead Toxicity in Adults: Measuring Dose and Consideration of Other Methodologic Issues[J]. Environ Health Perspect., 2007, 115 (3): 455-462.
[49] Koller, K.,Brown, T.,Spurgeon, A., etc. Recent developments in low-level lead exposure and intellectual impairment in children[J]. Environmental health perspectives, 2004, 112 (9): 987-94.
[50] F, F.,A, E.,CK, R., etc. Lead-induced dysregulation of superoxide dismutases, catalase, glutathione peroxidase, and guanylate cyclase[J]. Environmental research, 2005, 98 (1): 33–39.
[51] R, S.,M, N.,P, V. Combined efficacies of lipoic acid and meso-2,3-dimercaptosuccinic acid on lead-induced erythrocyte membrane lipid peroxidation and antioxidant status in rats[J]. Human & Experimental Toxicology, 2003, 22 (4): 183-192.
[52] Gurer, H.,Ercal, N. Can antioxidants be beneficial in the treatment of lead poisoning?[J]. Free Radical Biology and Medicine, 2000, 29 (10): 927–945.
[53] Gardner, H. W. Oxygen radical chemistry of polyunsaturated fatty acids[J]. Free Radical Biology and Medicine, 1989, 7 (1): 65-86.
[54] Zhang H, Liu Y, Liu R, et al. Molecular Mechanism of Lead-Induced Superoxide Dismutase Inactivation in Zebrafish Livers.[J]. Journal of Physical Chemistry B, 2014, 118 (51): 18420-18426.
[55] Gerber, G. B.,Maes, J.,Gilliavod, N., etc. Brain biochemistry of infant mice and rats exposed to lead [J]. Toxicology letters, 1978, 2 (1): 51–63.
[56] Shafiq-ur-Rehman,S, R.,O, C., etc. Evaluation of malondialdehyde as an index of lead damage in rat brain homogenates[J]. Biometals : an international journal on the role of metal ions in biology, biochemistry, and medicine, 1995, 8 (4): 275-279.
[57] Shafiq Ur, R. Lead-induced regional lipid peroxidation in brain[J]. Toxicology letters, 1984, 21 (3): 333-337.
[58] Adonaylo, V. N.,Oteiza, P. I. Lead intoxication: antioxidant defenses and oxidative damage in rat brain[J]. Toxicology, 1999, 135 (2-3): 77-85.
[59] 刘芳丽,薛振菲,张楠, etc. 白藜芦醇对铅诱导小鼠脂质过氧化的拮抗作用[J]. 卫生研究, 2012, 41 (6).
[60] Yedjou, C. G.,Tchounwou, P. B. N-acetyl-l-cysteine affords protection against lead-induced cytotoxicity and oxidative stress in human liver carcinoma (HepG2) cells[J]. International journal of environmental research and public health, 2007, 4 (2): 132-137.
[61] A, S.,F, M.,F, B., etc. Stress proteins and oxidative damage in a renal derived cell line exposed to inorganic mercury and lead[J]. Toxicology, 2009, 264 (3): 215-224.
[62] LG, N.-M.,MA, Q.-E.,S, G., etc. Effects of lead intoxication on intercellular junctions and biochemical alterations of the renal proximal tubule cells[J]. Toxicology in Vitro, 2009, 23 (7): 1298–1304.
[63] AL, J.,GT, R.,PG., G. Contribution of protein kinase C and glutamate in Pb(2+)-induced cytotoxicity[J]. Toxicology letters, 2000, 115 (2): 89–98.
[64] Chan, D. C. Mitochondria: Dynamic Organelles in Disease, Aging, and Development[J]. Cell, 2006, 125 (7): 1241–1252.
[65] JM, S.,M, L.,M, B., etc. Relation between mitochondrial membrane potential and ROS formation[J]. Methods in Molecular Biology (Clifton, N.J.), 2012.
[66] Scherz-Shouval, R.,Elazar, Z. ROS, mitochondria and the regulation of autophagy[J]. Trends in Cell Biology, 2007, 17 (9): 422-427.
[67] KA, F.,F, G.,FJ, G., etc. Optical And Pharmacological Tools To Investigate The Role Of Mitochondria During Oxidative Stress And Neurodegeneration[J]. Progress in Neurobiology, 2006, 79 (3): 136–171.
[68] Green, D. R.,Reed, J. C. Mitochondria and apoptosis[J]. Science (New York, NY), 1998, 281 (5381): 1309-1312.
[69] Wang, L.,Wang, H.,Li, J., etc. Simultaneous effects of lead and cadmium on primary cultures of rat proximal tubular cells: interaction of apoptosis and oxidative stress[J]. Archives of environmental contamination and toxicology, 2011, 61 (3): 500-511.
[70] J, X.,LD, J.,LH., X. Lead-induced apoptosis in PC 12 cells: involvement of p53, Bcl-2 family and caspase-3[J]. Toxicology letters, 2006, 166 (2): 160–167.
[71] 孔繁翔,尹大强,严囯安. 环境生物学[M]. 高等教育出版社, 2000.
[72] Z, C.,R, L.,H., Z. Noncovalent interaction of oxytetracycline with the enzyme trypsin[J]. Biomacromolecules, 2010, 11 (9): 2454-2459.
[73] W, Z.,R, L.,F, S., etc. A new strategy to identify and eliminate the inner filter effects by outer filter technique[J]. Journal of fluorescence, 2011, 21 (3): 1249-1254.
[74] Luciani, X.,Mounier, S.,Redon, R., etc. A simple correction method of inner filter effects affecting FEEM and its application to the PARAFAC decomposition[J]. Chemometrics and Intelligent Laboratory Systems, 2009, 96 (2): 227–238.
[75] Weert, M. v. d.,Stella, L. Fluorescence quenching and ligand binding: A critical discussion of a popular methodology[J]. Journal of Molecular Structure, 2011, 998 144–150.
[76] Keswani, N.,Choudhary, S.,Kishore, N. Quantitative aspects of recognition of the antibiotic drug oxytetracycline by bovine serum albumin: Calorimetric and spectroscopic studies[J]. The Journal of Chemical Thermodynamics, 2013, 58 (3): 196–205.
[77] JX, H.,MA, C.,MA, B., etc. Drug-binding energetics of human α-1-acid glycoprotein assessed by isothermal titration calorimetry and molecular docking simulations [J]. Journal of Molecular Recognition, 2012, 25 (12): 642–656.
[78] Zhang H, Hao F, Liu R. Interactions of lead (II) acetate with the enzyme lysozyme: A spectroscopic investigation [J]. Journal of Luminescence, 2013, 142(142):144-149.
[79] Zhang H, Liu Y, Zhang R, et al. Binding mode investigations on the interaction of lead(II) acetate with human chorionic gonadotropin [J]. Journal of Physical Chemistry B, 2014, 118(32):9644-9650.
[80] Hao Z, Kai W, Zhang M, et al. Assessing the mechanism of DNA damage induced by lead through direct and indirect interactions[J]. Journal of Photochemistry & Photobiology B Biology, 2014, 136 (7):46-53.
