红彩真猎蝽对斜纹夜蛾幼虫捕食行为及室内扩散能力的研究
2016-11-16苏湘宁邓海滨朱丹荔蔡青年张茂新
苏湘宁,邓海滨,朱丹荔,蔡青年,张茂新
1 华南农业大学广东省生物农药创制与应用重点实验室,广东广州 510642;
2 广东省烟草南雄科学研究所,广东南雄 512400;
3 中国农业大学植物保护学院,北京海淀 100193
红彩真猎蝽对斜纹夜蛾幼虫捕食行为及室内扩散能力的研究
苏湘宁1,邓海滨2,朱丹荔1,蔡青年3,张茂新1
1 华南农业大学广东省生物农药创制与应用重点实验室,广东广州 510642;
2 广东省烟草南雄科学研究所,广东南雄 512400;
3 中国农业大学植物保护学院,北京海淀 100193
为掌握红彩真猎对斜纹夜蛾蝽捕食能力的强弱,采用培养皿和自制仪器装置目测观察,在室内条件下研究了红彩真猎蝽成虫和1~5龄若虫对斜纹夜蛾幼虫的捕食行为以及红彩真猎蝽1~5龄若虫及成虫的水平爬行能力和垂直攀爬能力。结果表明,成虫的取食时间随斜纹夜蛾幼虫龄期的增加而增加,4龄和5龄若虫的取食时间较其它龄期若虫的取食时间长。成虫的爬行时间随斜纹夜蛾幼虫龄期的增加而减少。随着红彩真猎蝽龄期的增加第一次刺入时间逐渐减少。随着红彩真猎蝽龄期的增加最长取食时间逐渐减少,取食量逐渐增多,红彩真猎蝽成虫和4-5龄高龄若虫的捕食能力最强。红彩真猎蝽5龄若虫及成虫的扩散能力很强,龄期越大室内扩散能力越强。这些研究结果对制定烟田释放红彩真猎蝽防治烟草害虫控制策略具有重要的理论意义。
红彩真猎蝽;斜纹夜蛾;捕食行为;水平爬行能力;垂直攀爬能力
红彩真猎蝽 (Harpactor fuscipes(Fabricius))属半翅目、猎蝽科,是重要的捕食性天敌,主要分布在广东、广西和福建等南方种烟区[1],是广东烟草种植区的优势天敌种类[2],国内已经有初步的研究报道。红彩真猎蝽的成虫和幼虫均可捕食斜纹夜蛾(Spodopteralitura(Fab.))[3],烟青虫(Helicoverpa assulta(Guenee))[4]和烟蚜(Myzus persicae(Sulzer))等重要烟草害虫[5]。这种猎蝽在广东省南雄市1年可繁殖2代,以成虫越冬,第1代的若虫和成虫的发育历期完全覆盖烟草大田生长期,能充分控制烟草害虫的危害[3]。
捕食性天敌的捕食行为和扩散能力是决定其捕食效果的主要因子之一,可直接影响它们的控制效应[6]。捕食性天敌捕食行为和扩散能力的研究是评价天敌的基础,是开展生物防治的依据。此前针对瓢虫捕食行为和扩散能力研究较多[6-8],而红彩真猎蝽对斜纹夜蛾捕食行为和室内扩散能力的研究国内外未见相关文献。为了充分利用红彩真猎蝽对烟草害虫进行有效控制,本研究以红彩真猎蝽成虫及1~5龄若虫为对象,研究其对各龄期斜纹夜蛾的捕食行为和扩散能力。
1 材料与方法
1.1 供试虫源与试验条件
供试红彩真猎蝽2014年采自广东省烟草公司南雄科学研究所试验烟田,2龄斜纹夜蛾幼虫 在(27±1) ℃, 湿 度 为(60±15)%, 光 照L∶D=16h∶8h的条件下饲养,扩繁约6代后,选择羽化8h内的健康成虫,和蜕皮8h内的健康若虫用于实验。斜纹夜蛾采自广东省烟草公司南雄科学研究所试验烟田,网室烟草上饲养至少2代作为实验种群。
1.2 试验方法
1.2.1 红彩真猎蝽捕食斜纹夜蛾幼虫的各行为阶段时间分配
试验方法参考姚松林等方法并略加改进[8]。在培养皿(直径5 cm,高1 cm)中放入新鲜烟草嫩叶片(ф=2 cm),叶片上接入大小一致的1~5龄斜纹夜蛾幼虫,1~2龄幼虫数量为10头,3~4龄幼虫为5头,5龄幼虫为3头,在培养皿内(ф=5 cm)接入饥饿24 h的红彩真猎蝽成虫或1~5龄若虫1头,目测观察并记录60 min内红彩真猎蝽爬行、取食、休息、清洁时间。每头猎蝽仅实验1次,每个处理10头虫,重复3次,试验在(25±1)℃、相对湿度(60±15)%、光线均匀条件下进行。
1.2.2 红彩真猎蝽捕食斜纹夜蛾幼虫时的第1次刺入斜纹夜蛾时间和最长取食时间
试验方法同上,记录60 min内,红彩真猎蝽接入培养皿到红彩真猎蝽第1次刺入斜纹夜蛾的时间;并记录红彩真猎蝽在多次取食中最长的取食时间。
1.2.3 红彩真猎蝽捕食斜纹夜蛾幼虫的刺探次数
试验方法同上,并记录60 min内红彩真猎蝽刺探猎物的次数,即口器试探猎物但没有取食猎物的次数。
1.2.4 红彩真猎蝽捕食斜纹夜蛾幼虫的取食量
试验方法同上,并记录60 min内红彩真猎蝽取食斜纹夜蛾的头数。
1.2.5 红彩真猎蝽室内水平爬行能力测定
室内扩散能力试验采用自制仪器装置[9]。水平扩散试验装置直径1 m,高2 cm透明塑料圆筒,圆筒顶部放一张透明塑料板,并将1 m长的红色细线粘在上部塑料板正中央一个点,圆筒底部放置画有正方形(5×5 cm)格子的纸,方格纸粘在透明玻璃柜子上,透明柜子下面用日光灯向上打光,周围环境保持黑暗。将1头1~5龄红彩真猎蝽若虫或成虫轻轻垂直放入粘细线点下面的圆筒中部,用细线代表若虫爬行轨迹,拐点处用透明胶粘在塑料板上,观察记录包括10 min内猎蝽爬行的总距离、10 min内的爬行时间和休息时间、轨迹。每个处理10头,重复3次。计算各龄期红彩真猎蝽水平爬行速度。
1.2.6 红彩真猎蝽室内垂直攀爬能力测定
垂直攀爬试验装置采用直径5 cm,高50 cm透明圆纸筒。将1头红彩真猎蝽1~5龄若虫或成虫轻轻放入透明圆纸筒底部,如果红彩真猎蝽爬到圆筒顶部,立即将圆筒倒转。每个龄期红彩真猎蝽的每个处理10头,重复3次。观察30 min内猎蝽爬行情况,观察记录包括红彩真猎蝽的垂直爬行的总高度、爬行时间和休息时间。每个处理10头,重复3次。计算各龄期红彩真猎蝽垂直攀爬速度。
1.3 数据处理
所有数据采用SPSS11.5软件统计分析。差异显著性比较采用one-way ANOVA和LSD法,P≤0.05和0.01分别表示显著和极显著差异,以a、b、c表示其差异性,数字后不含有相同字母表示差异有统计学意义。
2 结果与分析
2.1 红彩真猎蝽捕食斜纹夜蛾幼虫的各行为阶段时间分配
由表1可以看出,红彩真猎蝽捕食1龄斜纹夜蛾,红彩真猎蝽1龄若虫的爬行时间最短,为8.30 min。红彩真猎蝽3龄若虫取食时间最短,为39.03 min,红彩真猎蝽4龄和5龄若虫取食时间最长,在45.32~45.51 min,占总观测时间的75.53%~75.85%。红彩真猎蝽4龄和5龄若虫的休息时间最短,分别为4.19 min和4.94 min,两者没有差异(df=6,F=7.135,P4龄,5龄=0.353)。在所有行为中,清洁时间占总观测时间的0.22%~1.27%,在各行为时间中所占比例最少。可以得出红彩真猎蝽4~5龄若虫对1龄斜纹夜蛾幼虫的捕食能力最强。
红彩真猎蝽捕食2龄斜纹夜蛾,3龄若虫爬行时间最长,为11.45 min。5龄取食时间最长为52.49 min,3龄取食时间最短为40.04 min。5龄红彩真猎蝽若虫休息时间最短为2.00 min,3龄最长为8.08 min。1-5龄红彩真猎蝽若虫清洁时间的差异性不显著(df=6,F=13.338,P1龄-5龄=0.203),雌成虫和雄成虫差异性不显著(df=6,F=13.338,P1龄-5龄=0.120)。成虫的清洁时间比若虫多。可以得出红彩真猎蝽5龄若虫对2龄斜纹夜蛾幼虫的捕食能力最强。
红彩真猎蝽捕食3龄斜纹夜蛾,未发现1龄红彩真猎蝽若虫成功取食3龄斜纹夜蛾幼虫的行为。红彩真猎蝽2-5龄若虫爬行时间差异显著,雌成虫和雄成虫爬行时间差异不显著(df=5,F=44.370,P雌,雄=0.396),红彩真猎蝽5龄若虫的取食时间最长为52.78 min,3龄若虫的取食时间最短为46.37 min,红彩真猎蝽2龄若虫休息时间最短为1.98 min,红彩真猎蝽3龄若虫休息时间最长为5.46 min。随着红彩真猎蝽龄期的增加,其清洁时间有增多趋势。可以得出红彩真猎蝽5龄若虫对3龄斜纹夜蛾幼虫的捕食能力最强。
红彩真猎蝽捕食4龄斜纹夜蛾,未发现1龄和2龄红彩真猎蝽若虫成功取食4龄斜纹夜蛾幼虫的行为。3龄红彩真猎蝽爬行时间最长为13.92 min,占总行为时间的23.20%,雌雄成虫爬行时间较短,分别为3.10 min和3.46 min,红彩真猎蝽4龄、5龄、雌成虫和雄成虫的取食时间差异不显著(df=4,F=58.829,P=0.062)。红彩真猎蝽3龄若虫的休息时间最长为9.99 min,红彩真猎蝽4龄若虫休息时间最短为2.77 min。红彩真猎蝽3龄若虫的清洁时间最短为0.09 min,红彩真猎蝽雌雄成虫的清洁时间较长,分别为0.82 min和0.94 min。可以得出红彩真猎蝽成虫对4龄斜纹夜蛾幼虫的捕食能力最强。
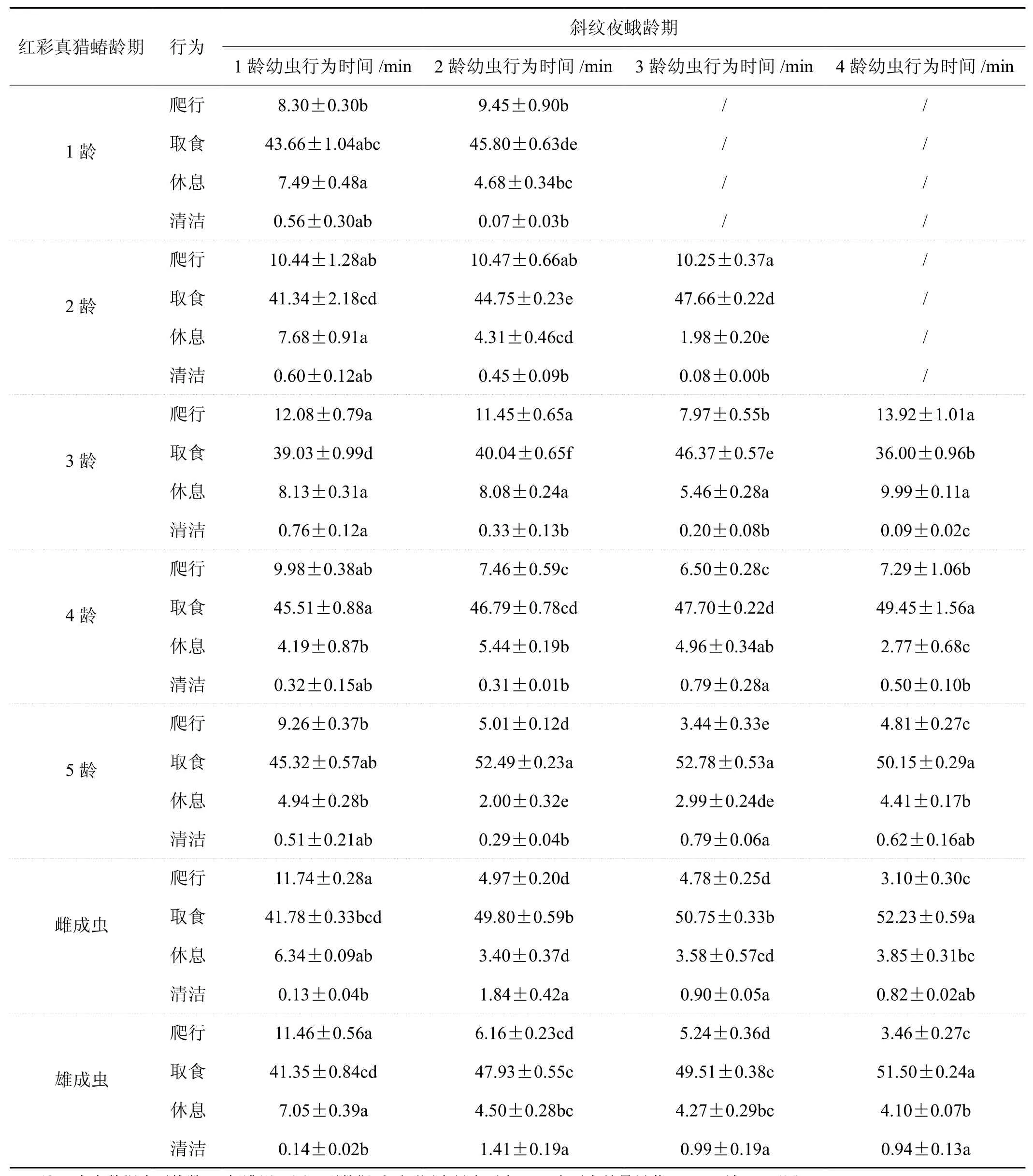
表1 不同龄期的红彩真猎蝽捕食1~4龄斜纹夜蛾幼虫的行为时间Tab.1 Time spent of H. fuscipes at different stages in four behavioral categories of feeding on 1st~4th instars larvae of S. litura
2.2 红彩真猎蝽捕食斜纹夜蛾的第1次刺入时间和最长取食时间
由表2可以看出,随着红彩真猎蝽龄期的增加,第1次刺入时间有减少的趋势。红彩真猎蝽成虫和1~5若虫捕食1龄斜纹夜蛾幼虫的第1次刺入时间没有规律性,其中3龄红彩真猎蝽若虫的第1次刺入时间最长为11.33 min,1龄红彩真猎蝽若虫的第1次刺入时间最短为6.80 min。红彩真猎蝽成虫和1~5若虫捕食2~4龄斜纹夜蛾幼虫时,随着红彩真猎蝽龄期的增加,其第1次刺入时间逐渐减少。在红彩真猎蝽捕食2龄斜纹夜蛾中,红彩真猎蝽2龄和3龄若虫的第1次刺入时间差异不显著(df=6,F=42.505,P=0.448),红彩真猎蝽5龄若虫、雌成虫和雄成虫第1次刺入时间差异性不显著(df=6,F=42.505,P=0.650);在红彩真猎蝽捕食3龄斜纹夜蛾中,红彩真猎蝽5龄若虫、雌成虫和雄成虫第1次刺入时间差异性不显著(df=5,F=9.12,P=0.454);在红彩真猎蝽捕食4龄斜纹夜蛾中,红彩真猎蝽4龄和5龄若虫第1次刺入时间差异不显著(df=4,F=18.474,P=0.123),5龄若虫、雌成虫和雄成虫第1次刺入时间差异不显著(df=4,F=18.474,P=0.209)。
由表3可知,红彩真猎蝽成虫和1~5龄若虫捕食1~3龄斜纹夜蛾低龄幼虫的最长取食时间,随着红彩真猎蝽龄期的增加,其最长时间逐渐减少;红彩真猎蝽成虫和1~5龄若虫捕食4龄斜纹夜蛾高龄幼虫的最长取食时间,随着红彩真猎蝽龄期的增加,其最长时间逐渐增加。在红彩真猎蝽捕食1龄斜纹夜蛾幼虫中,红彩真猎蝽4龄和5龄若虫最长取食时间差异不显著,雌成虫和雄成虫的最长取食时间差异不显著;在红彩真猎蝽捕食4龄斜纹夜蛾幼虫中,红彩真猎蝽3龄若虫最长取食时间最短为36.00 min,红彩真猎蝽4龄若虫、5龄若虫、雌成虫和雄成虫的最长取食时间差异不显著(df=4,F=57.708,P=0.064)。

表2 不同龄期红彩真猎蝽捕食斜纹夜蛾时的第1次刺入时间Tab.2 First pierce time of H. fuscipes at different stages of feeding on four instars larvae of S. litura

表3 不同龄期红彩真猎蝽捕食斜纹夜蛾时的最长取食时间Tab.3 The longest feeding time of H. fuscipes at different stages of feeding on four instars larvae of S. litura
2.3 红彩真猎蝽取食斜纹夜蛾的刺探次数
由表4可知,红彩真猎蝽取食斜纹夜蛾的刺探次数差别不大。在红彩真猎蝽成虫和1~5龄若虫捕食1龄斜纹夜蛾幼虫中,刺探次数均差异不显著(df=6,F=0.833,P=0.256);在红彩真猎蝽成虫和1~5龄若虫捕食2龄斜纹夜蛾幼虫中,1龄红彩真猎蝽若虫刺探次数最多为1.30次,红彩真猎蝽2~5龄若虫及成虫刺探次数均差异不显著(df=6,F=4.136,P=0.095);在红彩真猎蝽成虫和1~5龄若虫捕食3龄斜纹夜蛾幼虫中,2龄红彩真猎蝽若虫刺探次数最多为1.20次,红彩真猎蝽2龄若虫和3龄若虫刺探次数差异不显著(df=5,F=9.120,P=0.454),红彩真猎蝽4龄、5龄、雌成虫和雄成虫差异不显著(df=5,F=9.120,P=0.486);在红彩真猎蝽成虫和1~5龄若虫捕食4龄斜纹夜蛾幼虫中,红彩真猎蝽3龄若虫刺探次数最多为1.87次,红彩真猎蝽5龄若虫、雌成虫和雄成虫差异不显著(df=4,F=52.9,P=0.644)。
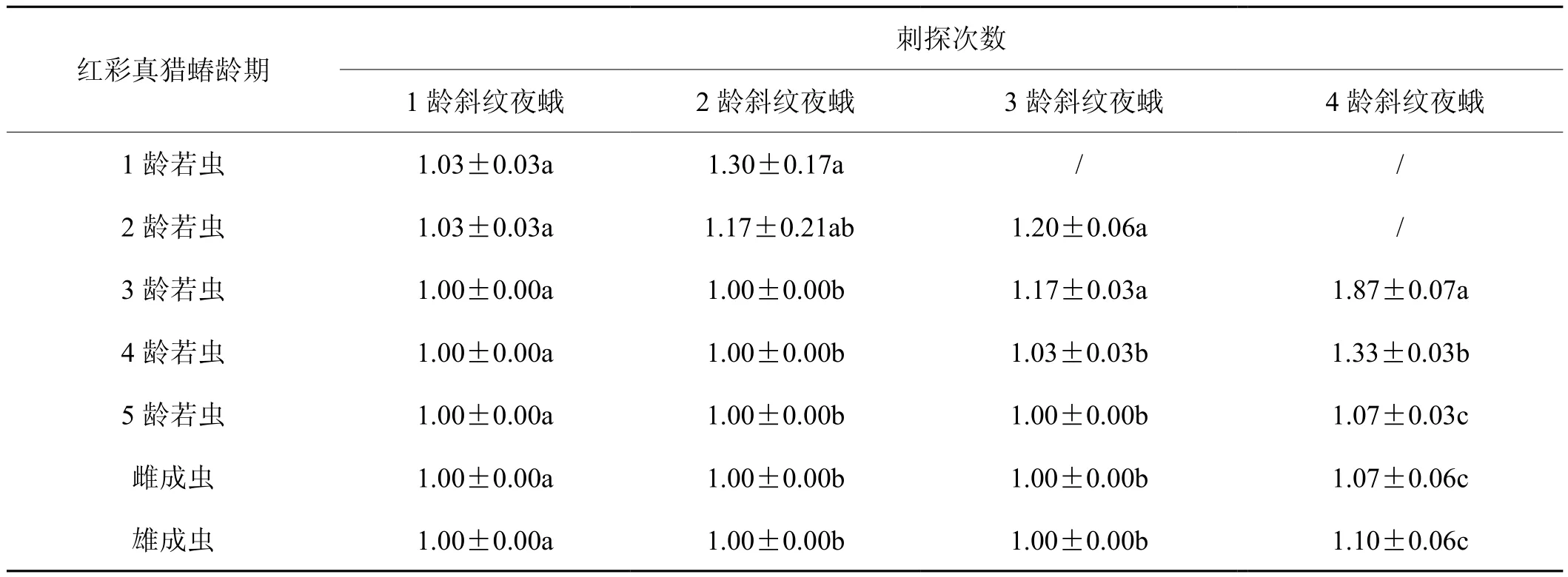
表4 不同龄期红彩真猎蝽捕食斜纹夜蛾时的刺探次数Tab.4 Piercetime of H. fuscipes at different stages of feeding on four instars larvae of S. litura
2.4 红彩真猎蝽捕食斜纹夜蛾的取食量
由表5可知,在红彩真猎蝽成虫和1~5龄若虫捕食1~3龄斜纹夜蛾低龄幼虫中,随着红彩真猎蝽龄期的增加,其取食量有增加的趋势。在红彩真猎蝽成虫和1~5龄若虫捕食4龄斜纹夜蛾高龄幼虫中,红彩真猎蝽均只取食1头;在红彩真猎蝽捕食1龄斜纹夜蛾幼虫中,2龄若虫和3龄若虫的取食头数为1.93,取食量差异不显著,雌成虫和雄成虫的取食量差异不显著;在红彩真猎蝽捕食2龄斜纹夜蛾幼虫中,1~3龄红彩真猎蝽若虫取食头数均为1头,雌成虫和雄成虫的取食量差异不显著(df=6,F=297.961,P=0.091);在红彩真猎蝽捕食3龄斜纹夜蛾幼虫中,2-3龄红彩真猎蝽若虫的取食头数均为1头,雌成虫和雄成虫取食头数差异不显著(df=5,F=84.533,P=0.712)。
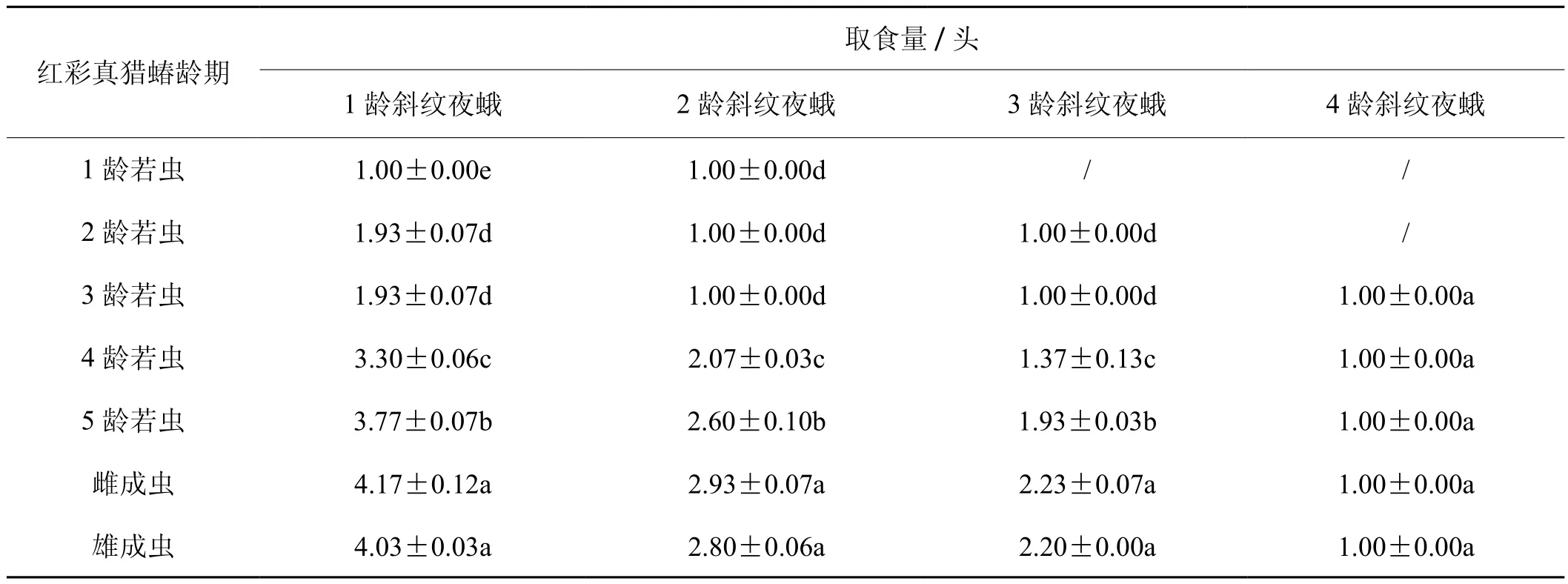
表5 不同龄期红彩真猎蝽捕食斜纹夜蛾时的取食量Tab.5 Feeding amount of H. fuscipes at different stages of feeding on four instars larvae of S. litura
2.5 红彩真猎蝽水平爬行试验
红彩真猎蝽水平爬行距离随其龄期的增加而增加,红彩真猎蝽成虫水平爬行距离显著高于各龄期红彩真猎蝽若虫。红彩真猎蝽雄成虫和雌成虫水平爬行距离分别为416.65 cm 和421.58 cm,差异不显著(n=3,P=0.765),1龄和2龄红彩真猎蝽若虫水平爬行距离差异不显著(n=3,P=0.194)。红彩真猎蝽成虫与4龄和5龄若虫爬行时间差异不显著(n=3,P=0.069),3~5龄红彩真猎蝽若虫爬行时间差异不显著(n=3,P=0.062),1~3龄红彩真猎蝽若虫爬行时间差异不显著(n=3,P=0.192)红彩真猎蝽水平爬行速度随其龄期的增加而增加。红彩真猎蝽雌雄成虫水平爬行平均速度差异不显著(n=3,P=0.657),各龄期红彩真猎蝽若虫水平爬行平均速度差异显著。因此,在10 min的时间内,红彩真猎蝽龄期越大水平爬行距离越长,其中红彩真猎蝽成虫爬行距离最长,各龄期若虫中5龄若虫爬行距离最长,1龄最短。红彩真猎蝽龄期越大水平爬行速度越快,其中红彩真猎蝽成虫水平爬行平均速度最快,红彩真猎蝽各龄期若虫中5龄若虫水平爬行平均速度最快,1龄最慢。所以,红彩真猎蝽龄期越大,其水平扩散能力越强(表6)。
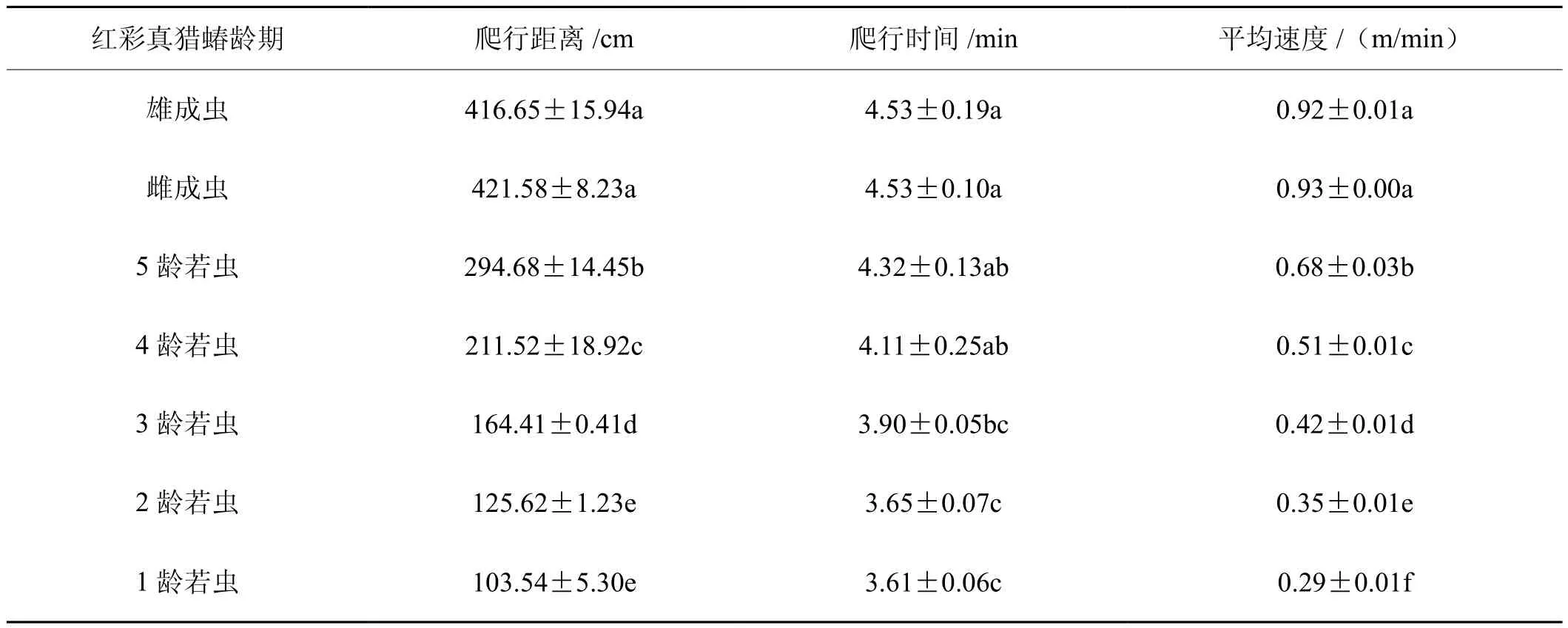
表6 不同龄期红彩真猎蝽的水平扩散能力Tab.6 Horizontal walking capacity of H. fuscipes at different stages
2.6 红彩真猎蝽垂直攀爬试验
红彩真猎蝽垂直攀爬高度随其龄期的增加而增加,红彩真猎蝽成虫垂直爬行高度显著高于各龄期红彩真猎蝽若虫。红彩真猎蝽雌雄成虫垂直爬行高度差异不显著(n=3,P=0.084),1龄和2龄红彩真猎蝽若虫垂直爬行高度差异不显著(n=3,P=0.061)2龄和3龄红彩真猎蝽若虫垂直爬行高度差异不显著(n=3,P=0.185),3龄和4龄红彩真猎蝽若虫垂直爬行高度差异不显著(n=3,P=0.097)。红彩真猎蝽雌雄成虫垂直爬行时间差异不显著(n=3,P=0.178),红彩真猎蝽雌成虫和5龄若虫垂直爬行时间差异不显著(n=3,P=0.140)。3~5龄红彩真猎蝽若虫垂直爬行时间差异不显著(n=3,P=0.074),1~4龄红彩真猎蝽若虫垂直爬行时间差异不显著(n=3,P=0.250)。红彩真猎蝽垂直攀爬平均速度随其龄期的增加而增加。红彩真猎蝽雌雄成虫垂直爬行平均速度差异不显著(n=3,P=0.370),4龄和5龄红彩真猎蝽若虫垂直爬行平均速度差异不显著(n=3,P=0.071)。因此,在30 min的时间内,红彩真猎蝽龄期越大垂直爬行高度越大,其中红彩真猎蝽成虫垂直爬行高度最大。各龄期若虫中5龄若虫垂直爬行高度最大,1龄最小。红彩真猎蝽龄期越大垂直爬行速度越快,其中红彩真猎蝽成虫垂直爬行平均速度最快,红彩真猎蝽各龄期若虫中5龄若虫垂直爬行平均速度最快,1龄最慢。所以,红彩真猎蝽龄期越大,其垂直扩散能力越强(表7)。
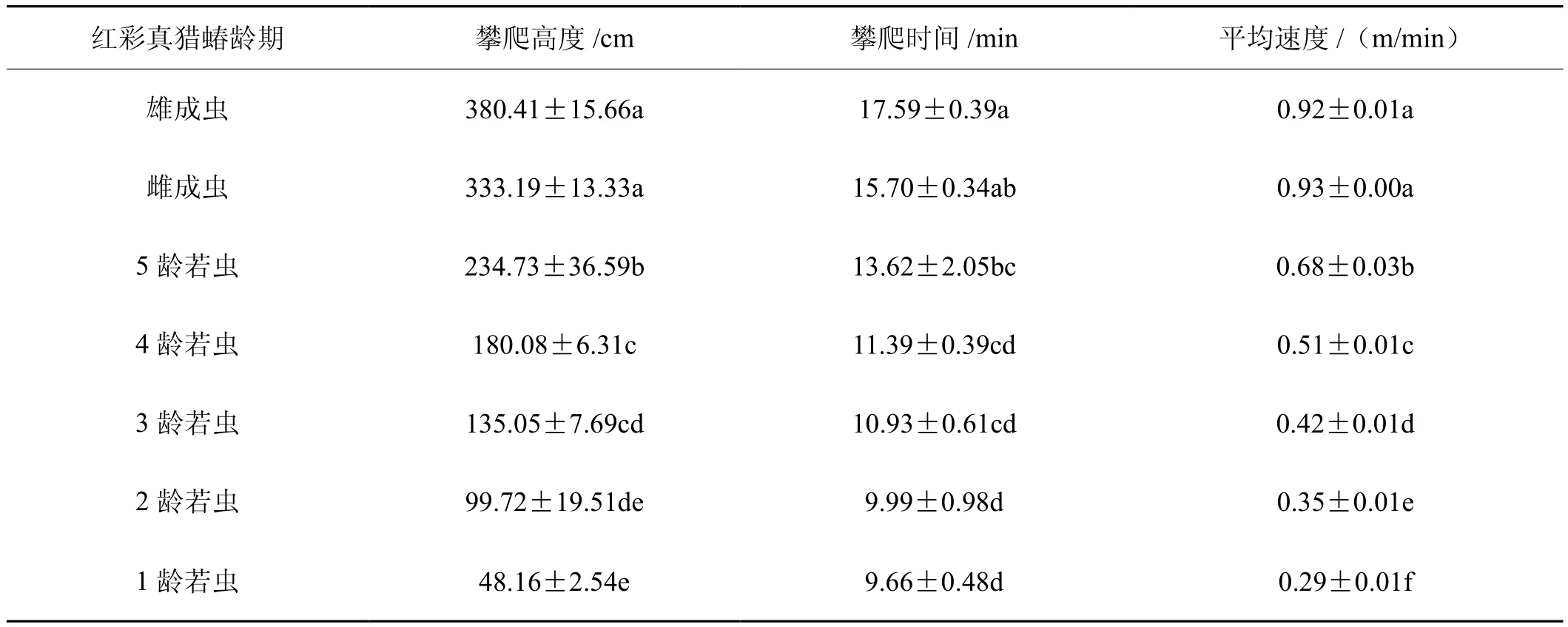
表7 不同龄期红彩真猎蝽的垂直扩散能力Tab. 7 Vertical walking capacity of H. fuscipes at different stages
3 讨论
3.1 红彩真猎蝽捕食斜纹夜蛾幼虫时的各行为阶段时间分配
红彩真猎蝽捕食斜纹夜蛾幼虫时的各行为阶段时间分配及时间所占比例根据捕食者和猎物龄期的不同而异。其中3龄红彩真猎蝽若虫的爬行和休息时间较长,取食时间较短。可能的原因是3龄红彩真猎蝽若虫的捕食能力较其它龄期红彩真猎蝽弱。5龄红彩真猎蝽若虫和红彩真猎蝽雌雄成虫的取食时间比较长,所占行为时间比例较大。说明红彩真猎蝽5龄若虫和雌雄成虫的捕食能力强。这与大多数瓢虫幼虫的典型行为相似[10-12]。这些结论与烟粉虱天敌日本刀角瓢虫(Serangium japonicum)捕食烟粉虱的行为类似[8]。
3.2 红彩真猎蝽捕食斜纹夜蛾幼虫时的第一次刺入时间和最长取食时间
红彩真猎蝽捕食斜纹夜蛾幼虫时的第一次刺入时间和最长取食时间随捕食者和猎物龄期的不同而异。第一次刺入时间的长短一定程度上反映出红彩真猎蝽的捕食能力强弱。本文可以得出龄期越大的红彩真猎蝽捕食能力越强的结论。最长取食时间的长短一定程度上反映出捕食者对猎物的取食效率高低。本文的研究结果证明了红彩真猎蝽龄期越大取食效率越高。
3.3 红彩真猎蝽取食斜纹夜蛾的刺探次数
对斜纹夜蛾的刺探次数在一定程度上能反应出红彩真猎蝽对不同龄期猎物取食的难易程度。只有在较低龄的红彩真猎蝽捕食高龄的斜纹夜蛾幼虫时才出现刺探多次的现象,可能原因是斜纹夜蛾高龄幼虫虫体大,低龄红彩真猎蝽在捕食过程中比较难刺入斜纹夜蛾高龄幼虫导致的。
3.4 红彩真猎蝽捕食斜纹夜蛾的取食量
本研究发现,由于红彩真猎蝽龄期越大需补充的营养越多,取食量就越大。
目前有关捕食效能和捕食行为的研究有明显进步,已经从室内利用培养皿中放置叶片,向利用透明观察罩内放置寄主植株进行模拟田间的观察,并从目测观察向利用3台摄像机自动记录的方向发展[7]。在本论文的试验设置上,采用培养皿(直径5 cm,高1 cm)中放置叶片、目测观察1 h的方法进行了捕食行为观测。因此,试验结果的可靠性与实际情况可能存在差距。但是,仍然可以在一定程度上反映出红彩真猎蝽捕食斜纹夜蛾的行为特征。
3.5 红彩真猎蝽扩散能力
捕食性天敌的扩散能力的强弱直接影响捕食者对害虫的控制效果[13]。比如,根据龟纹瓢虫的扩散飞行能力强弱可以来制定特定区域内的害虫控制策略[13]。关于捕食性天敌扩散能力强弱研究报道并不多,对红彩真猎蝽室内扩散能力的研究国内外未见报道。
随着红彩真猎蝽龄期的增加其扩散能力逐渐增加,红彩真猎蝽成虫的扩散能力最强,室外条件下成虫有一定的飞行能力,甚至在种植大棚里也会飞到棚外,并不能保持长效的灭虫效果。如果红彩真猎蝽成虫飞行扩散能力极强则不利于对特定区域内害虫的防治。比如,采用飞行模的方法测定瓢虫本地种和外来种的扩散能力[14],瓢虫成虫飞行能力很强,增加了防治害虫的难度。在吴迪的研究中说明飞行能力低的法国种群异色瓢虫4龄幼虫、2龄幼虫及成虫对豆蚜有较强的控制能力[15]。目前日本有研究报道,有些瓢虫飞行能力较低,通过30多代能力较低的瓢虫个体交配,培育出不能飞行的异色瓢虫,取得了满意的灭蚜效果[16]。因此,扩散能力极强的红彩真猎蝽成虫不利于对特定区域内害虫的防治,不能飞行的4~5龄红彩真猎蝽若虫是适合防治应用的红彩真猎蝽龄期。
4 结论
本文通过研究红彩真猎蝽的捕食行为和扩散能力,从而确定出其捕食能力的强弱。红彩真猎蝽4~5龄若虫及成虫的捕食能力很强,另外红彩真猎蝽4~5龄若虫及成虫的扩散能力很强,但成虫能飞行不是适合防治应用的红彩真猎蝽龄期。因此,红彩真猎蝽4~5龄若虫是应用防治斜纹夜蛾的合适龄期。这些结论可为有效应用红彩真猎蝽防治斜纹夜蛾等烟草重要害虫奠定基础。本实验红彩真猎蝽的捕食行为是在培养皿中观察研究,扩散能力是在透明圆筒中研究,还需要进一步进行田间试验验证。
[1]吴钜文,彩万志,侯陶谦. 中国烟草昆虫种类及害虫综合治理[M].北京:中国农业科学技术出版社, 2003∶129.
WU Juwen, CAI Wanzhi, HOU Taoqian. China tobacco insect species and integrated pest management[G].China Agricultural Science and Technology Publishing House,2003∶129.
[2]周忠实,陈泽鹏,邓海滨,等.不同干扰因素对斜纹猫蛛(Oxyopes sertatus)和红彩真猎蝽(Harpactor fuscipes)捕食作用的影响(英文)[J].生态学报.2007(08)∶ 3341-3347.
ZHOU Zhongshi, CHEN Zepeng, DENG Haibin, et al.Effect of interference factors on predations ofOxyopes sertatus(L.)Koch(Araneae∶Oxyopidae)andHarpactor fuscipes(F.)(Hemiptera∶Reduviidae)onSpodoptera litura(F.)[J]. ActaEcologicaSinica, 2007(08)∶ 3341-3347.
[3]邓海滨,吕永华,邱妙文,等. 捕食性天敌红彩真猎蝽的生物学特性研究[J]. 中国烟草科学. 2014(02)∶ 109-112.
DENG Haibin, LV Yonghua, QIU Miaowen, et al.Studies on the biological characteristics of predatory bugHarpactorfuscipes[J].Chinese Tobacco Science,2014(02)∶109-112.
[4]邓海滨,王珍,陈永明,等. 红彩真猎蝽对斜纹夜蛾和烟青虫的捕食功能反应[J]. 广东农业科学. 2012(13)∶107-109.
DENG Haibin, WANG Zhen, CHEN Yongming, et al.Predation ofHarpactor fuscipesonHelicoverpa assulta Spodoptera litura[J]. Guangdong Agricultural Sciences,2012(13)∶ 107-109.
[5]邓海滨,吕永华,田明义,等. 红彩真猎蝽对烟蚜的捕食功能反应及寻找效应[J]. 中国烟草学报. 2015, 21(5)∶74-78.
DENG Haibin, LV Yonghua, TIAN Mingyi,et al.Functional responses and searching efficiency of Harpactor fuscipes on predation of Myzuspersicae[J]. Acta Tabacaria Sinica, 2015,21(5)∶ 74-78.
[6]魏开炬,詹祖仁,林滨,等. 大突肩瓢虫对居竹伪角蚜捕食行为的观察[J]. 中国森林病虫. 2015, 34(4)∶ 26-29.
WEI Kaiju, ZHAN Zuren, LIN Bin, et al. Observation on the predatory behavior ofsynonycha grandisonPseudoregma bambusicola[J]. Forest Pest Disease, 2015,34(4)∶ 26-29.
[7]苗静,李绍建,邱宝利,等. 烟粉虱天敌沙巴拟刀角瓢虫与越南斧瓢虫的捕食行为比较[J]. 昆虫知识.2010(04)∶ 700-704.
MIAO Jing, LI Shaojian, QIU Baoli, et al. The predatory behaviors ofSerangiella sababensisandAxinoscymnus apioides.[J]. Chinese Bulletin of Entomology.2010(04)∶700-704.
[8]姚松林,任顺祥,黄振. 烟粉虱天敌日本刀角瓢虫的捕食行为[J]. 应用生态学报. 2005(03)∶ 509-513.
YAO Songlin, REN Shunxiang, HUANG Zhen.Feeding behavior ofSerangium japonicum(Coleoptera∶Coccinellida e),a predator ofBemisia tabaci(Homoptera∶Aleyrodidae)[J].Chinese Journal of Applied Ecology.2005(03)∶ 509-513.
[9]Doo-Hyung Lee, Anne L. Nielsen, Tracy C. Leskey.Dispersal Capacity and Behavior of Nymphal Stages of Halyomorphahalys(Hemiptera∶Pentatomidae)Evaluated Under Laboratory and Field Conditions[J]. Journal of Insect Behavior. 2014, 27(5)∶ 639-651.
[10] Dixon A F C. Factors limiting the effectiveness of the coccinellid beetle,Adalia bipunctata(L.), as a predator of Sycamore aphid,Drepanosiphum platanoides(scher)[J].Anim Ecol. 1970, 39∶ 739-751.
[11]Hoelmer K A, Osborne L S, Yokomi P K. Reproduction and feeding behavior ofDelphastus pusillus(Coleoptera∶Coccinellidae),Predator of Bemisia argentifolii (Homoptera∶ Aleyrodidae)[J].Econ Ent, 1993(86)∶ 322-329.
[12] Dixon A F C. An Experimental study of the searching behavior of the predatory coccinellidbeetleAdalia decempunctata(L.)[J]. AnimEcol, 1959, 28(2)∶ 259-281.
[13] 赵秀芝,刘玉升,张帆.龟纹瓢虫不同飞行力种群的扩散及控害能力研究[C]//公共植保与绿色防控——中国植物保护学会2010年学术年会论文集.鹤壁∶中国植物保护学会,2010∶842.
ZHAOXiuzhi, LIU Yusheng, ZHANG Fan.Dispersal and pests control capacity ofPropylaea japonica(Coleoptera∶Coccinellidae)in different flight capability populations[C]//Public plant protection and green prevention and control-The academic conference proceedings of China Society of Plant Protection 2010. Hebi∶ China Society of Plant Protection,2010∶842.
[14] Sara Maes, Xavier Massart, Jean-Claude Gre´goire, et al.Dispersal potential of native and exotic predatory ladybirds as measured by a computer-monitored flight mill[J].BioControl. 2014, 59(4)∶ 415-425.
[15] 吴迪. 异色瓢虫法国种群飞行能力及捕食效应研究[D].甘肃农业大学, 2008.
WU Di. Study on Flying Ability and Predatory Function Response ofHarmonia axyridis(France population)[D].Gansu Agricultural University,2008.
[16] 杨光.日本将瓢虫改良成飞不走的“生物农药”[N/OL].世界农药国际,2004.17.044. http∶∥www.pesticide.com.cn/
YANG Guang. Ladybird wereimproved not flying“biopesticide” in Japan [N/OL]. World agrochemical international,2004.17.044.http∶//www.pesticide.com.cn/
Study on indoor dispersive capacity and predatory behavior of Harpactor fuscipes (Fabricius)to larvae of Spodopteralitura
SU Xiangning1, DENG Haibin2, ZHU Danli1, CAI Qingnian3, ZHANG Maoxin1
1 Key Laboratory of Bio-Pesticide Innovation and Application of Guangdong Province, South China Agricultural University,Guangzhou 510642, China;
2 Guangdong Nanxiong Tobacco Research Institute, Nanxiong, Guangdong 512400, China;
3 College of Plant Protection, China Agricultural University, Beijing 100193, China
In order to learn the ability of predation of H. fuscipes to Spodopteralitura, the Spodopteralitura-attacking behavior of H.fuscipesadults and 1st-5thinstar larvae, horizontal and vertical walking capacity of H. fuscipes larvae (i.e.,1st through 5th)and adults were examined by culture dish and self-apparatus in laboratory. Results showed that time for starved adults to feed S. lituranymphs increased as host nymphs developed. 4th-instar and 5th-larvae of H. fuscipesspent had more time in feeding than other larvae instar. With the number of larval instars increasing, the time spent on walking decreased for H. fuscipesadults. With the number of instars of H. fuscipes increasing,first pierce time and the longest feeding time of H. fuscipes were decreased. With the number of instars of H. fuscipes increasing, feeding amount of H. fuscipes was increased. Predatory capacity of H. fuscipes adults and 4th-5thinstar larvae were the strongest of all instars. H.fuscipes adults and 5thinstar larvae had strong dispersal capacity. In the lab, the dispersal capacity of H. fuscipes increased as assassin bug instar developed. These research results had important theoretical significance for formulating strategy of releasing assassin bug to control these important pests.
Harpactor fuscipes(Fabricius);Spodopteralitura;feeding behavior;horizontal walking capacity;vertical walking capacity
苏湘宁,邓海滨,朱丹荔,等. 红彩真猎蝽对斜纹夜蛾幼虫捕食行为及室内扩散能力的研究[J]. 中国烟草学报,2016,22(5)
广东省烟草专卖局(公司)科技项目“红彩真猎蝽控害机理及人工饲养技术研究和应用”(No.201304)
苏湘宁(1990—),硕士,研究方向为昆虫生态学,Tel:020-85280203, Email:851513723@qq.com
张茂新(1957—),教授,主要从事昆虫生态学研究,Tel:020-85280203,Email:mxzhang@scau.edu.cn
2016-04-22
:SU Xiangning, DENG Haibin, ZHU Danli, et al. Study on indoor dispersive capacity and predatory behavior ofHarpactor fuscipes(Fabricius)to larvae ofSpodopteralitura[J]. Acta Tabacaria Sinica, 2016, 22(5)
