高温预处理对力竭运动后人血液白细胞HSP70mRNA及自由基代谢的影响
2016-08-10张治远张月娟
陈 婷,张治远,张月娟
(西藏民族大学 体育学院,陕西 咸阳 712082)
高温预处理对力竭运动后人血液白细胞HSP70mRNA及自由基代谢的影响
陈婷,张治远,张月娟
(西藏民族大学 体育学院,陕西 咸阳712082)
摘要:目的:探讨一次及多次高温预处理对递增负荷至力竭运动后人血液白细胞 HSP70mRNA及自由基代谢的影响。方法:受试者随机分为运动对照组(C),一次高温预处理组(P1),多次高温预处理组(P2)。P1组进行一次性高温预处理1 h(桑拿,温度46±1 ℃,湿度85%-90%,体重下降2%)。P2组每隔三天进行一次(共八次)高温预处理。C组采用Bruce方案在跑台上进行递增负荷运动至力竭。P1和P2组高温预处理结束,在室温下恢复24 h后,在跑台上进行递增负荷运动至力竭。运动结束后记录最大心率、最大运动通气量、最大摄氧量、呼吸商等,同时记录通气阈时的通气量、最大摄氧量利用率等。C组、P1组及P2组于运动前、运动后即刻、运动后1 h分别采肘静脉血。采用RT-PCR法检测受试者血液白细胞HSP70mRNA表达,用硫代巴比妥(TBA)法测定血清丙二醛(MDA)含量,用黄嘌呤氧化酶法测定血清超氧化物歧化酶(SOD)活性,采用全血乳酸分析仪测定血乳酸值。结果:C组力竭运动后即刻,人血液白细胞HSP70mRNA表达和血清MDA含量较安静时显著增加(P<0.01,P<0.05)。C组运动后1 h,HSP70mRNA表达及MDA含量较运动后即刻显著减少(均P<0.05),而血清SOD活性较运动后即刻显著下降(P<0.05)。P1组在安静时、运动后即刻及运动后1 h,均较C组相同时间点的HSP70mRNA显著升高(均P<0.05)。而P1组在运动后1 h较C组同时间点的SOD活性显著升高(P<0.05)。P2组血液在运动后即刻和运动后1 h,均较C组相同时间点的HSP70mRNA表达及血清SOD活性显著升高(均P<0.05)。而P2组在安静、运动后即刻及运动后1 h较C组同时间点的MDA含量均显著减少(均P<0.05)。P2组在安静时、运动后即刻及运动后1 h,均较P1组相同时间点的HSP70mRNA显著下降(均P<0.05)。P1组、P2组力竭运动中通气阈时通气量VE较C组均显著增加(P<0.05),且受试者血液白细胞HSP70mRNA表达与血清SOD/MDA比例呈显著正相关(R=0.48,P<0.05)。结论:一次及多次高温预处理可上调运动后人血液白细胞HSP70mRNA表达,抑制力竭运动所造成的氧化应激损伤。且多次高温预处理后HSP70mRNA表达降低,可能是对高温重复应激的适应性反应。
关键词:高温预处理;HSP70mRNA;氧自由基;抗氧化酶;递增负荷运动;力竭
研究发现,在缺血神经元损伤[1],心肌梗死及心脏[2]、肝脏[3]、脊髓缺血再灌注损伤[4]等疾病模型中,高温预处理均可发挥保护作用。其有益作用与热休克蛋白70(HSP70)表达上调呈正相关[3]。HSP70在正常细胞中水平较低,而环境和生理应激(包括热、冷、缺血、缺氧、损伤、运动等)状态下HSP70可显著升高[5-6]。研究表明,小鼠骨骼肌HSP70过表达可保护肌肉免于萎缩及骨骼肌功能紊乱[7],心脏HSP70过表达可抑制缺血再灌注心肌细胞凋亡水平[8]。而敲除HSP70后的脑损伤小鼠,细胞死亡水平增加,活性氧ROS诱导的p53基因表达上调,导致外伤性脑部损伤进一步恶化[9]。可见HSP70在提高细胞存活率、减轻细胞受损伤程度、维持机体内稳态中发挥着重要的保护作用。有文献报道,一次力竭运动[10]和长期运动训练[11]均可造成机体HSP70合成增加,且高温预处理后24 h人白细胞中HSP70蛋白表达较安静时显著上调。但关于高温预处理对力竭运动后人血液白细胞HSP70mRNA表达的影响未见报道,HSP70mRNA是否会对高温重复应激产生适应亦未见报道。因此,本研究建立一次和多次两种高温预处理方式,采用一次递增负荷至力竭运动方案,探讨一次及多次高温预处理对力竭运动后人血液白细胞HSP70mRNA 及自由基代谢的影响,旨在为高温预处理方式防治运动损伤研究提供实验依据。
1材料与方法
1.1 实验受试者与分组
受试者为华南师范大学体育科学学院20名健康男性大学生。随机分为2组:一次高温预处理组(P1)、多次高温预处理组(P2),每组10人。另外运动对照组(C)与P2组受试者相同,以此做多次高温干预的前后对照。受试者在实验期间禁止饮酒,常规活动,禁止大强度运动。C组采用Bruce方法在跑台上进行递增负荷运动至力竭,作为力竭运动对照组。P1组干预方式为一次性高温预处理1 h,P2组高温预处理时间为一个月,每隔三天一次(共八次),P1和P2组室温恢复24 h后采用Bruce方法在跑台上进行递增负荷运动至力竭。
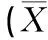
表1 受试者身体特征±SD)
1.2主要仪器和试剂
主要仪器为Quinton Q55活动跑台、2900心肺功能测试仪、Polar遥测心率表、YSI-1500 SPORT全血乳酸分析仪、PTC 200 PCR仪、Apollo水平电泳仪、BIO凝胶成像系统、722分光光度计、TU-1901紫外分光光度计、广州气体厂液氮等。主要试剂包括全血总RNA快速提取试剂盒、通用RT-PCR试剂盒、通用PCR优化试剂盒、HSPcDNA的引物、DNA Marker、EB、琼脂糖、南京建成SOD和MDA测试盒。
1.3高温预处理实验方案
高温预处理方案在文献[12-13]基础上改良[12-13]。P1组受试者高温预处理1 h(桑拿,温度46±1℃,湿度85%—90%,体重下降2%),受试者分两组进行桑拿,实验人员陪同,观察和询问受试者情况。于安静、高温预处理0.5 h、高温预处理后即刻记录直肠温度。P2组受试者每间隔三天进行一次(共八次)高温预处理,桑拿方式同P1组。
1.4受试者运动方案
C组采用Bruce方法在跑台上进行递增负荷运动至力竭[14-15](Bruce运动方案见表2)。P1组高温预处理1 h及P2组多次高温预处理结束,均在室温(温度25±0.5 ℃,湿度60%—65%)恢复24 h后,在跑台上进行递增负荷运动至力竭。运动方式同C组。
表2 Bruce运动方案
1.5最大摄氧量测定
采用Bruce运动方案,使用2 900气体代谢仪进行测试,参照文献[16]判断VO2max:①吸氧量不再增加而出现平台;②呼吸商大于1.1;③心率大于180/分;④血乳酸浓度在8 mmol/L以上。以上任何3种条件出现即可确定为最大摄氧量[16]。运动结束后记录最大心率(HRmax)、最大运动通气量(VEmax)、相对最大摄氧量(VO2max/kg)、最大摄氧量(VO2max)、呼吸商(R)、运动力竭时间(Time)。通气阈参照通气曲线进行判断[17],判断方法为:①通气量非线性上升的起点;②二氧化碳排出量非线性上升的起点;③二氧化碳当量非线性上升并不伴随氧当量的变化。记录通气阈时各指标,包括心率(HR)、通气量(VE)、最大摄氧量利用率(%VO2max)、呼吸商(R)、最大心率百分比(%HRmax)。
1.6受试者血液采集
C组、P1组及P2组于运动前安静、递增负荷运动后即刻、运动后1 h分别采肘静脉血,测试受试者血液白细胞HSP70mRNA、血清丙二醛(MDA)、超氧化物歧化酶(SOD)、血乳酸。
1.7RT-PCR
取1 ml的EDTA-K2抗凝血,倒入DEPC处理的离心管,9 000转离心2 min,去上清,加入1 ml的Trizol,4 ℃下12 000转离心5 min。离心后加入氯仿0.2 mL,4 ℃下12 000转离心15 min。加入异丙醇0.5 mL,4℃下12 000转离心20 min,加入75%乙醇1 mL,4℃下10 000转离心5 min。吸去剩余乙醇。取适量DEPC水溶解RNA。分装出5 μl用于RNA定量,余下RNA用于反转录。利用紫外分光光度计测定所提取样本总RNA浓度(μg/μl)。以A260/A280在1.8—2.0范围作为RNA纯度较好的标准。取4 μlRNA进行1%琼脂糖凝胶电泳,观察18S和28S的完整性,检测RNA有无降解。
反转录步骤:将提取的样本RNA以2 μg/μl的浓度确定所需体积(<11 μl),按照反转录试剂盒操作步骤加入Oligo(dT)1 μl,样本RNA,加DEPC水至总体积12 μl,65 ℃孵育5 min。按反转录试剂盒操作步骤分别加入5×Reaction Buffer 4 μl、RiboLockTM Rnase Inhibitor (20 μ/μl) 1 μl、10mM dNTP Mix 2 μl、RevertAidTM M-MμLV Reverse Transcriptase(200 μ/μl) 1 μl,混合均匀后离心。42 ℃孵育60 min,70 ℃加热5 min终止反应,获得RT产物。
PCR步骤:按照试剂盒操作步骤,配成25 μl反应体系。分别加入2×Taq MasterMix 12.5 μl、Forward Primer(10 μM)1 μl、Reverse Primer(10 μM)1 μl、Template DNA<1 μg,加Rnase-free Water至总体积25 μl。充分混匀后离心,进行PCR反应(PCR引物见表3)。
PCR产物分析:取PCR产物各7.5 μl,进行1%琼脂糖凝胶电泳。电压80V,电泳60 min,之后采用Syngene凝胶成像系统成像并定量分析。以HSP70与β-actin扩增条带的成像面积积分比值来评定HSP70mRNA表达水平。

表3 PCR引物
1.8血清MDA含量和SOD活性测定
采用硫代巴比妥(TBA)法测定血清丙二醛(MDA)含量,用黄嘌呤氧化酶法测定超氧化物歧化酶(SOD)活性,操作按照试剂盒说明书的规定程序进行。
1.9血乳酸值测试
用美国产YSI-1500 SPORT全血乳酸分析仪测定血乳酸值。
1.10数据采集与统计学处理
2实验结果
2.1高温预处理前中后受试者不同时刻直肠温度的比较
P1组受试者直肠温度在高温预处理0.5 h、1 h均较高温预处理前极显著升高(P<0.01),高温预处理1 h较0.5 h极显著升高(P<0.01)。P2组受试者进行8次高温预处理,直肠温度总平均值在高温预处理0.5 h、1 h均较高温前极显著升高(P<0.01)。高温预处理1 h较0.5 h极显著升高(P<0.01),见表4。
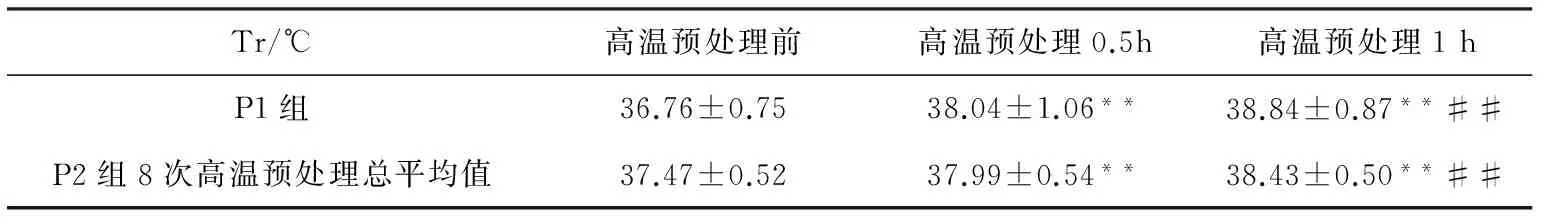
表4 高温预处理前中后不同时刻受试者直肠温度
注:*表示与高温预处理前相比,P<0.05;**表示与高温预处理前相比,P<0.01;#表示与高温预处理0.5 h相比,P<0.05;##表示与高温预处理0.5 h相比,P<0.01。
2.2高温预处理前后受试者裸重的比较
P1组受试者高温预处理后裸重极显著下降(P<0.01),下降百分比约为2%。P2组受试者进行8次高温预处理后,裸重总平均值极显著下降(P<0.01),下降百分比约为2%。见表5。
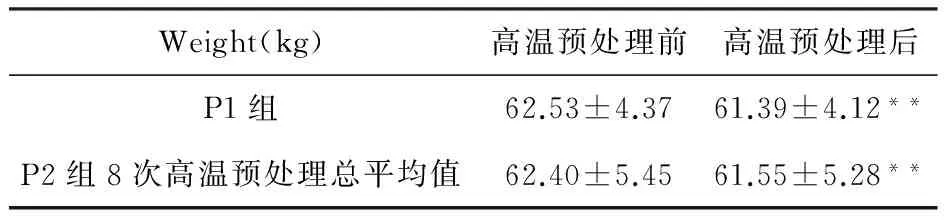
表5 高温预处理前后受试者裸重
注:**表示与高温预处理前相比,P<0.01。
2.3高温预处理前后血液白细胞HSP70mRNA的变化
C组受试者力竭运动后即刻,血液白细胞HSP70mRNA表达较安静值显著升高(P<0.01),升高了2.96倍。运动后1 h的HSP70mRNA表达较运动后即刻显著下降(P<0.05)。P1组HSP70mRNA在安静时、运动后即刻及运动后1 h,均较C组相同时间点的值显著升高(均P<0.05),且分别升高了10.29倍、2.34倍和10.97倍。P2组HSP70mRNA在运动后即刻和运动后1 h,均较C组相同时间点的值显著升高(P<0.05),且分别升高了0.93%和3.18倍。P2组HSP70mRNA在安静时、运动后即刻及运动后1 h,均较P1组相同时间点的值显著下降(P<0.05),且分别下降了72%、43%和65%。见图1、图2。
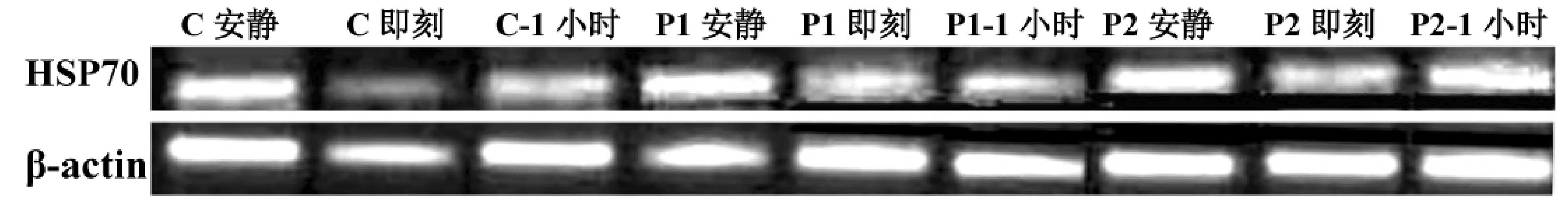
图1 受试者血液白细胞HSP70mRNA表达变化
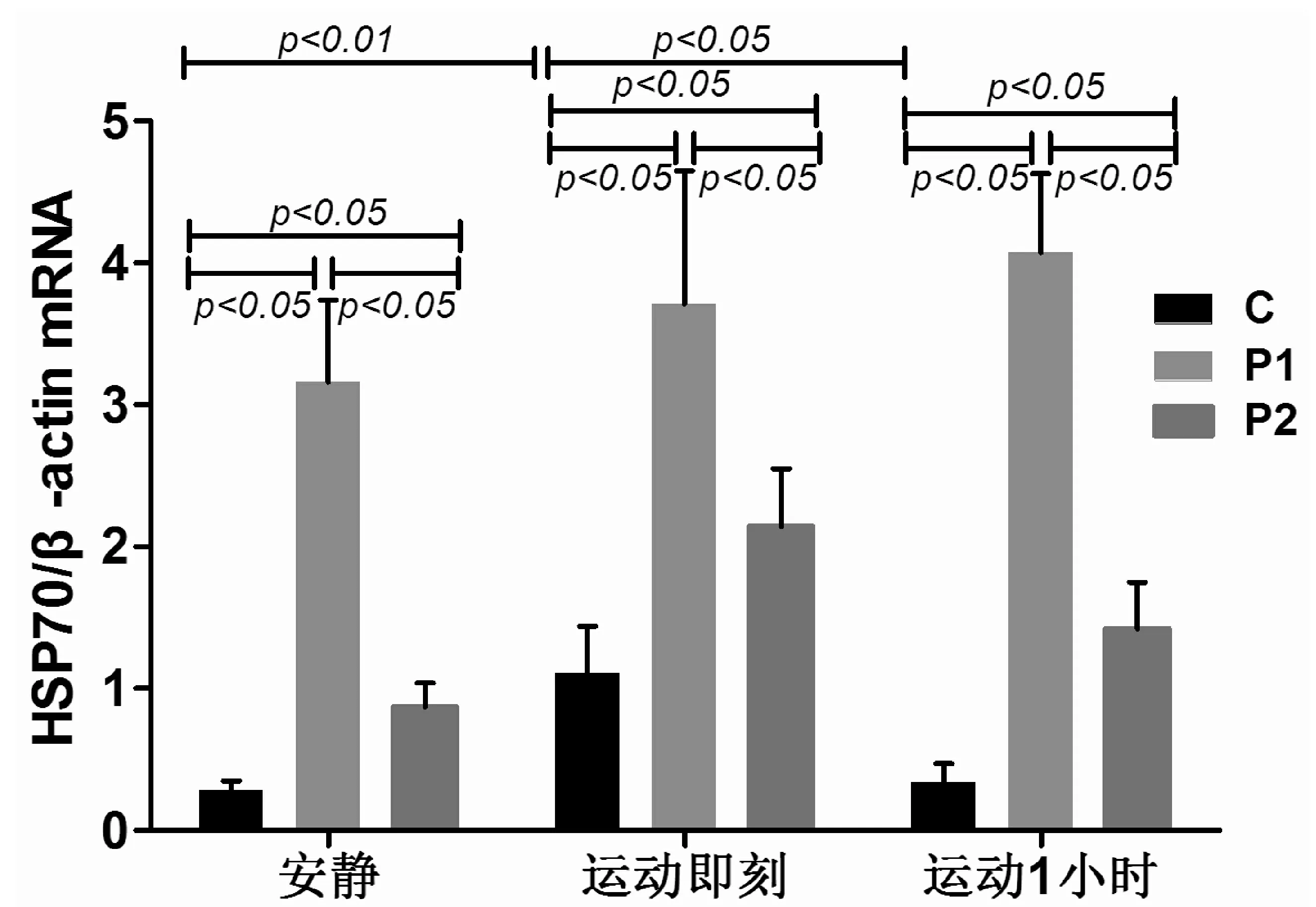
图2 受试者血液白细胞HSP70mRNA(HSP70/β-actin)面积积分比值
2.4高温预处理前后血清MDA含量、SOD活性的变化
C组力竭运动即刻血清MDA含量较安静时显著增加(P<0.05)。运动后1 h较即刻显著减少(P<0.05)。P2组MDA含量在安静、运动后即刻及运动后1 h较C组同时间点的值均显著减少(均P<0.05)。见图3。
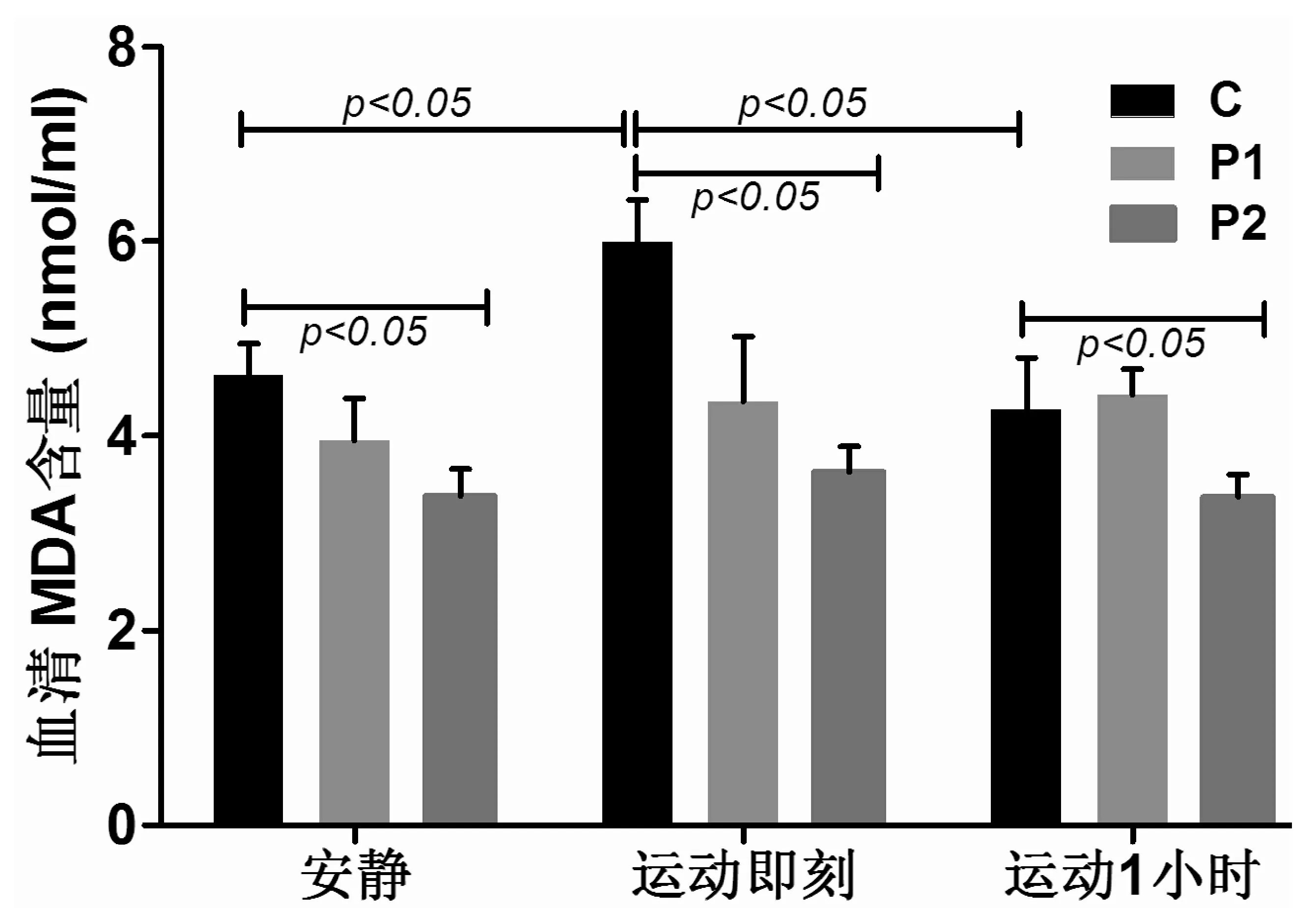
图3 受试者血清MDA含量(nmol/mL)变化
C组力竭运动后1 h血清SOD活性较运动后即刻显著下降(P<0.05)。P1组SOD活性在运动后1 h较C组同时间点的值显著升高(P<0.05)。P2组SOD活性在运动即刻和运动后1 h,均较C组同时间点的值显著升高(均P<0.05)。见图4。

图4 受试者血清SOD活性(μ/mL)变化
2.5受试者力竭运动时最大摄氧量、血乳酸等指标的变化
C组、P1组和P2组在力竭运动时,最大心率(HRmax)、最大运动通气量(VEmax)、相对最大摄氧量(VO2max/kg)、最大摄氧量(VO2max)、呼吸商(R)、运动力竭时间(Time)各指标均未出现显著性变化(P>0.05) 。见表6。
P1组、P2组力竭运动中通气阈时通气量(VE)较C组均显著增加(P<0.05),其他指标包括心率(HR)、最大摄氧量利用率(%VO2max)、呼吸商(R)、最大心率百分比(%HRmax)均未出现显著变化(P>0.05)。见表7。
C组、P1组和P2组受试者力竭运动后即刻血乳酸浓度较各组安静值均显著增加(均P<0.01)。C组、P1组和P2组受试者力竭运动后1 h血乳酸浓度较各组运动后即刻均显著降低(均P<0.01)。P2组运动后即刻血乳酸浓度较C组、P1组均显著增加(均P<0.01)。见图5。
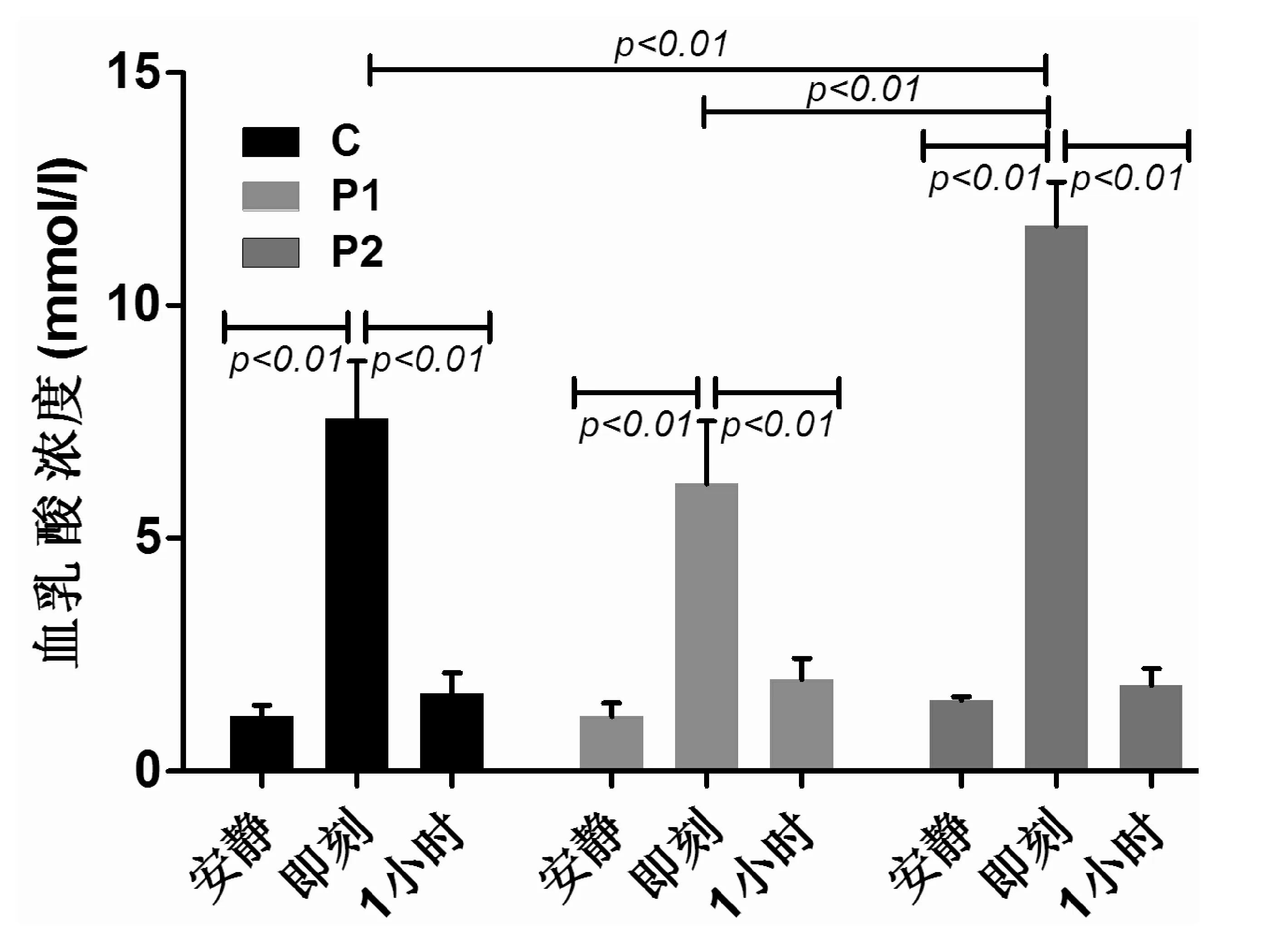
图5 受试者力竭运动时血乳酸浓度(mmol/l)的变化
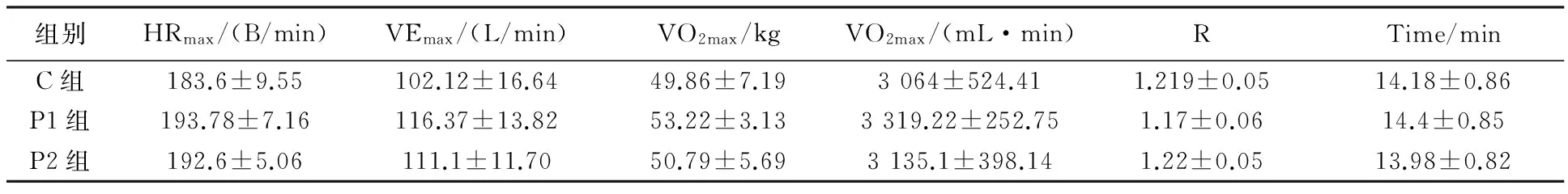
组别HRmax/(B/min)VEmax/(L/min)VO2max/kgVO2max/(mL·min)RTime/minC组183.6±9.55102.12±16.6449.86±7.193064±524.411.219±0.0514.18±0.86P1组193.78±7.16116.37±13.8253.22±3.133319.22±252.751.17±0.0614.4±0.85P2组192.6±5.06111.1±11.7050.79±5.693135.1±398.141.22±0.0513.98±0.82

表7 受试者力竭运动中通气阈时的各指标变化
注:*表示与C组相比,P<0.05。
2.6相关性分析
图6的相关性分析结果表明,受试者血液白细胞HSP70mRNA表达与血清SOD/MDA比例相关性密切,呈显著正相关(R=0.48,P<0.05)。表明一次及多次高温预处理后受试者血液白细胞HSP70mRNA表达增加,伴随血清SOD/MDA比例的增加。
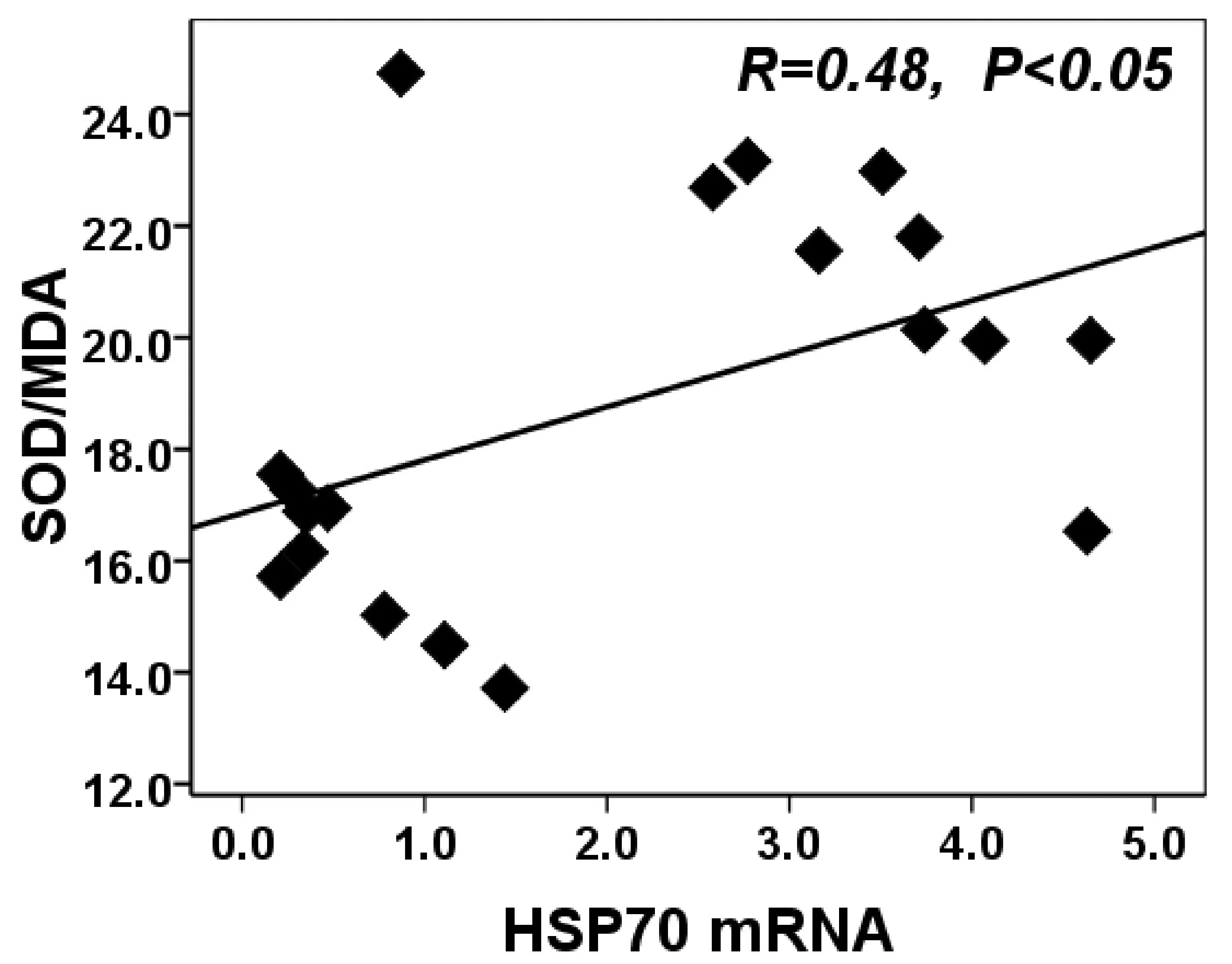
图6 受试者血液白细胞HSP70mRNA与血清SOD/MDA比例相关性示意图
3分析与讨论
有研究发现,一次力竭运动[10]和长期运动训练[11]均可造成机体HSP70合成增加。而两种运动方式的时相性表现为,一次力竭运动后30—60 min,骨骼肌HSP70mRNA表达达到峰值,之后逐渐下降,6 h后恢复安静水平[10]。而连续4周大强度训练,运动员肌肉HSP70蛋白水平(训练前为100%)从第一周到第四周分别增长181%、405%、456%和363%,呈下降趋势。HSP70蛋白峰值出现在训练第二周后[18]。同时研究发现运动员跑完半个马拉松后,白细胞胞浆HSP70在运动后即刻、3 h、24 h显著增加[19-20]。HSP70表达与运动时间和强度密切相关[21]。中等强度运动尚不足刺激HSP70表达增加,高强度训练对HSP70的诱导更为显著[22]。而关于运动诱导HSP70表达增加的机制,目前认为剧烈运动过程中ATP的减少、体温升高、pH值改变、氧化应激损伤、乳酸堆积、运动引起的肌肉收缩、血流改变等应激,都可能是诱导HSP70增加的刺激因素[23]。
而本实验证实,力竭运动后即刻人血液白细胞HSP70mRNA显著增加,运动后1 h逐渐降低。本实验还发现,一次及多次高温预处理可显著上调人血液白细胞HSP70mRNA表达,高温预处理与运动叠加后HSP70mRNA表达进一步增加。文献表明热应激可导致HSP70合成增多,其合成量与受热时间和程度有关。随着HSP70表达增加,细胞存活率增加,受损伤程度减轻[24]。通过高温预处理方式提前诱导HSP70产生,可增加机体抵抗损伤的能力,从而起到保护作用[2,12]。而HSP70表达的时相性研究发现,经冠状静脉窦逆行高温灌注稀释血液入心脏后大鼠心肌HSP70mRNA水平于15 min增加5倍,30 min增加10倍,60 min增加2倍[25]。而高温预处理后大鼠肝脏HSP70表达在8—24 h达到最高峰[25]。而本实验指标时相性结果发现,一次高温预处理白细胞HSP70mRNA在运动后即刻及运动后1 h,较运动对照组分别升高了2.34倍和10.97倍。而多次高温预处理HSP70mRNA在运动后即刻和运动后1 h,较运动对照组分别升高了0.93%和3.18倍。且多次高温预处理后HSP70mRNA表达较一次高温预处理降低,在安静时、运动后即刻及运动后1 h,分别下降了72%、43%和65%。表明可能是机体对高温重复应激的适应性反应。
本实验采用一次递增负荷至力竭的运动方案,发现运动对照组在运动后即刻,血清MDA含量较安静时显著增加,SOD活性有上升趋势,此时HSP70mRNA表达在运动后即刻较安静时显著升高。说明力竭运动使机体脂质过氧化水平增加,而抗氧化酶活性提高不足以对抗ROS的增加,引起机体氧化应激损伤。文献中降低氧化应激损伤的方式多样,本实验中采用了高温预处理方案。研究表明,高温预处理可增加大鼠腓肠肌和淋巴细胞HSP表达,提高肌纤维线粒体氧化酶活性,增加大鼠在高温环境中的运动时间和耐力水平[24]。且高温预处理的保护作用与HSP70表达上调呈正相关[3]。在本实验中,一次高温预处理后血清SOD活性在运动后即刻和运动后1 h,较运动对照组显著升高。多次高温预处理后MDA含量在运动后即刻和运动后1 h,较运动对照组显著降低。而SOD活性在运动后即刻和运动后1 h显著增加。此时一次及多次高温预处理组白细胞HSP70mRNA在运动后即刻和运动后1 h均显著升高,且HSP70mRNA表达与血清SOD/MDA比例呈显著正相关。提示高温预处理及运动双因素刺激后HSP70mRNA表达显著增高,可提高机体抗氧化酶活性,抑制脂质过氧化水平,减轻力竭运动造成的氧化应激损伤。同时本研究发现,一次及多次高温预处理组在运动后通气阈值VE均较运动对照组显著增加。通气阈值可反映人体有氧工作能力,提示两种高温预处理方式可增强受试者的有氧工作能力。
研究表明,HSP70的分子保护机制包括抗氧化作用[26]、协同免疫作用[27]、分子伴侣作用、抗细胞凋亡作用[28-29]。HSP具有抗氧化的生物活性,HSP与其结合物可激活蛋白激酶C,增强蛋白酶活性,促进ATP水解,刺激内源性抗氧化剂的合成和释放,从而抵抗过氧化物损伤。研究报道小鼠骨骼肌过表达HSP72,可增加血清抗氧化酶SOD活性,抑制氧化应激损伤,降低力竭运动造成的骨骼肌损伤和疲劳[26]。同时发现心肌过表达HSP72,可通过提高心脏Mn-SOD含量及活性,减少心肌细胞凋亡,对缺血再灌注心脏起到保护作用[29]。在本实验中两种高温预处理方式抑制了力竭运动造成的氧化应激损伤,其机制主要体现在白细胞HSP70mRNA的抗氧化作用。
4结论
本研究发现一次及多次高温预处理可上调运动后人血液白细胞HSP70mRNA表达,抑制力竭运动造成的氧化应激损伤。且多次高温预处理后HSP70mRNA表达降低,可能是对高温重复应激的适应性反应。
参考文献:
[1]Du F, Qian Z M, Zhu L, et al. A synergistic role of hyperthermic and pharmacological preconditioning to protect astrocytes against ischemia/reperfusion injury [J]. Neurochem Res, 2011,36(2):312-318.
[2]Juggi J S, Hoteit L J, Babiker F A, et al. Protective role of normothermic, hyperthermic and estrogen preconditioning and pretreatment on tumour necrosis factor-alpha-induced damage [J]. Exp Clin Cardiol, 2011,16(2):5-10.
[3]Mizushima Y, Wang P, Jarrar D, et al. Preinduction of heat shock proteins protects cardiac and hepatic functions following trauma and hemorrhage [J]. Am J Physiol Regul Integr Comp Physiol, 2000,278(2): 352-359.
[4]Zhang P, Abraham V S, Kraft K R,et al. Hyperthermic preconditioning protects against spinal cord ischemic injury [J]. Ann Thorac Surg, 2000,70(5):1490-1495.
[5]Kim N, Kim J Y, Yenari M A.Pharmacological induction of the 70-kDa heat shock protein protects against brain injury [J]. Neuroscience, 2015,284C(10):912-919.
[6]Kourtis N, Nikoletopoulou V, Tavernarakis N. Small heat-shock proteins protect from heat-stroke-associated neurodegeneration [J]. Nature, 2012,490(7419):213-218.
[7]McArdle A, Dillmann W H, Mestril R,et al. Overexpression of HSP70 in mouse skeletal muscle protects against muscle damage and age-related muscle dysfunction [J]. FASEB J,2001, 18(2): 355-357.
[8]Suzuki K, Sawa Y, Kagisaki K, et al. Reduction in myocardial apoptosis associated with overexpression of heat shock protein 70 [J]. Basic Res Cardiol,2000, 95(5):397-403.
[9]Eroglu B, Kimbler D E, Pang J, et al. Therapeutic inducers of the HSP70/HSP110 protect mice against traumatic brain injury [J]. J Neurochem, 2014,130(5):626-641.
[10]Thompson H S, Scordilis S P, Clarkson, P M, et al. A single bout of eccentric exercise increases HSP27 and HSC/HSP70 in human skeletal muscle [J]. Acta Physiol Scand, 2001,171(2):187-193.
[11]Milne K J, Noble E G. Exercise-induced elevation of HSP70 is intensity dependent [J]. J Appl Physiol, 2002,93(2):561-568.
[12]Du F, Zhu L, Qian Z M, et al. Hyperthermic preconditioning protects astrocytes from ischemia/reperfusion injury by up-regulation of HIF-1 alpha expression and binding activity [J]. Biochim Biophys Acta, 2010,1802(11):1048-1053.
[13]Ozveri E S, Bekraki A, Cingi A, et al. The effect of hyperthermic preconditioning on the immune system in rat peritonitis [J]. Intensive Care Med, 1999,25(10): 1155-1159.
[14]Fielding R A, Frontera W R, Hughes V A, et al. The reproducibility of the Bruce protocol exercise test for the determination of aerobic capacity in older women [J]. Med Sci Sports Exerc, 1997,29(8):1109-1113.
[15]van der Cammen-van Zijp M H M, Ijsselstijn H, Takken T, et al. Exercise testing of pre-school children using the Bruce treadmill protocol: new reference values [J]. Arbeitsphysiologie, 2009,108(2):393-399.
[16]Levine B D .VO2max: what do we know, and what do we still need to know?[J]. J Physiol,2008, 586(1):25-34.
[17]Gaskill S E, Ruby B C, Walker A J, et al. Validity and reliability of combining three methods to determine ventilatory threshold [J]. Med Sci Sports Exerc, 2001,33(11):1841-1848.
[18]Liu Y, Mayr S, Opitz-Gress A, et al. Human skeletal muscle HSP70 response to training in highly trained rowers [J]. J Appl Physiol, 1999,86(1): 101-104.
[19]Fehrenbach E, Passek F, Niess A M, et al. HSP expression in human leukocytes is modulated by endurance exercise [J]. Med Sci Sports Exerc, 2000,32(3):592-600.
[20]Walsh R C, Koukoulas I, Garnham A, et al. Exercise increases serum Hsp72 in humans [J]. Cell Stress Chaperones,2001, 6(4): 386-393.
[21]Liu Y, Lormes W, Baur C, et al. Human skeletal muscle HSP70 response to physical training depends on exercise intensity [J]. Int J Sports Med, 2000,21(5):351-355.
[22]Liu Y, Lormes W, Wang L, et al. Different skeletal muscle HSP70 responses to high-intensity strength training and low-intensity endurance training [J]. Arbeitsphysiologie,2004, 91(2-3): 330-335.
[23]Pösö A R, Eklund-Uusitalo S, Hyyppä S, et al. Induction of heat shock protein 72 mRNA in skeletal muscle by exercise and training [J]. Equine Vet J Suppl,2002,(34):214-218.
[24]Chen H W, Chen S C, Tsai J L, et al. Previous hyperthermic treatment increases mitochondria oxidative enzyme activity and exercise capacity in rats [J]. Kaohsiung J Med Sci, 1999,15(10):572-580.
[25]McCully J D, Myrmel T, Lotz M M, et al. The rapid expression of myocardial hsp 70 mRNA and the heat shock 70 kDa protein can be achieved after only a brief period of retrograde hyperthermic perfusion [J]. J Mol Cell Cardiol, 1995,27(3):873-882.
[26]Liu C C, Lin C H, Lin C Y, et al. Transgenic overexpression of heat shock protein 72 in mouse muscle protects against exhaustive exercise-induced skeletal muscle damage [J]. J Formos Med Assoc, 2013,112(1): 24-30.
[27]Chang C C, Chen S D, Lin T K, et al. Heat shock protein 70 protects against seizure-induced neuronal cell death in the hippocampus following experimental status epilepticus via inhibition of nuclear factor-kappaB activation-induced nitric oxide synthase II expression [J]. Neurobiol Dis, 2013(62): 241-249.
[28]Daugaard M, Rohde M, Jaattela M. The heat shock protein 70 family: Highly homologous proteins with overlapping and distinct functions [J]. Febs Lett, 2010,581(19): 3702-3710.
[29]Suzuki K, Murtuza B, Sammut I A, et al. Heat shock protein 72 enhances manganese superoxide dismutase activity during myocardial ischemia-reperfusion injury, associated with mitochondrial protection and apoptosis reduction [J]. Circulation, 2002,106(1):270-276.
收稿日期:2016-02-27
作者简介:陈婷(1984-),女,陕西西安人,讲师,博士,研究方向为运动生理学。
中图分类号:G804.2
文献标志码:A
文章编号:1008-3596(2016)04-0074-09
Effects of Hyperthermic Preconditioning on Blood Leucocyte HSP70mRNA and Oxygen Free Radical after Exhaustive Exercise
CHEN Ting, ZHANG Zhi-yuan, ZHANG Yue-juan
(School of Physical Education, Tibet University for Nationalities, Xianyang 712082, China)
Abstract:Objectives: To discuss the effects of two kinds of hyperthermic preconditioning on blood leucocyte HSP70mRNA and oxygen free radical after incremental exercise. Methods: The subjects are randomly divided into 3 groups: exercise control group(C) ,exercise after one hyperthermic preconditioning group(P1), and exercise after more hyperthermic preconditioning group(P2). P1 Group: The hyperthermic preconditioning is performed for an hour(Sauna, 46±1℃; relative humidity, 85%-90%). P2 Group: one hyperthermic preconditioning every three days, total eight times are performed. Group C applies Bruce protocol on treadmill to do incremental exercise to exhaustion. P1 and P2 Group are at rest under the normal temperature for 24 hours after hyperthermic preconditioning, and then do incremental exercise to exhaustion on treadmill. The maximum heart rate, maximal exercise ventilation, maximal oxygen uptake, respiration quotient are recorded at the end of the exercise, and the ventilation rate, maximal oxygen uptake utilization ratio are recorded at the same time. The elbow vein blood of C Group, P1 and P2 are collected before exercise, immediately after exercise, and 1 hour after exercise. By RT-PCR method, the tested groups’ expression of blood white cells HSP70mRNA are detected, content of malondialdehyde (MDA) in the serum is determined by the method of TBA and superoxide dismutase (SOD) activity is measured by the method of xanthine oxidase, blood lactic acid values is examined by the blood lactate analyzer. Results: in C Group, in immediate testing, the expression of HSP70mRNA and the content of serum MDA are significantly increased (P<0.01, P<0.05) compared with the test at rest. C Group after 1 hour’s exercise, HSP70mRNA expression and MDA content are significantly reduced compared with immediate testing (both P<0.05), while the serum SOD activity is significantly decreased (P<0.05) compared with immediate testing. Compared with C Group, HSP70mRNA expression in P1 Group increases significantly at rest, immediately and 1h after exercise(both P<0.05). SOD activity of P1 Group elevates significantly immediately after exercise (P<0.05). Compared with C Group, HSP70mRNA expression and SOD activity in P2 Group increases significantly at immediately and 1hour after exercise (both P<0.05), MDA content in P2 Group reduces markedly at rest, immediately and1hour after exercise (both P<0.05). Compared with P1 Group, P2 Group HSP70mRNA expression decreases markedly at rest, immediately after exercise, 1 hour after exercise (P<0.05). And HSP70mRNA is positively correlated with the production of SOD/MDA(R=0.48, P<0.05). Conclusions: Two kinds of hyperthermic preconditioning could increase leukocyte HSP70mRNA expression after incremental exercise, and reduce exhaustive exercise-induced oxidative damage. Leukocyte HSP70mRNA expression is decreased after one month hyperthermic preconditioning, this may be the adaptive response to repeated heat stress.
Key words:hyperthermic preconditioning; HSP70mRNA; oxygen free radical; superoxide dismutase; incremental exercise; exhaustion
