基于形态学特征和ITS序列分析秦巴山区天麻萌发菌的亲缘关系
2016-07-23虞小燕邓百万陈文强
虞小燕++邓百万++陈文强
摘要:利用拮抗试验,萌发试验和ITS序列分析技术对秦巴山区12个天麻萌发菌菌株进行了亲缘关系分析。结果表明:12个萌发菌菌株分为两大类,其中8103-4、8103-6、8103-7、8103-8这4个菌株与其他菌株拮抗反应明显,并且ITS序列分析显示在同一个分支上,亲缘关系相近;萌-3、萌-7、萌MD-2、萌发菌、石斛-1、石斛-2、萌-云南此8个菌株拮抗反应不明显,ITS序列分析显示在同一个分支上,8103-3、萌发菌与标准菌种Mycena citrinomarginata亲缘关系相近,萌MD-2、石斛小菇-1、萌-3、萌-7、石斛小菇-2、萌-云南与Mycena purpureofusca亲缘关系相近。
关键词:天麻萌发菌;拮抗;rDNA序列;亲缘关系
中图分类号:S182文献标志码:A文章编号:1002-1302(2016)05-0245-04
萌发菌隶属于担子菌纲(Basidiomyeetes)伞菌目(Agarieales)口蘑科(Trieholomataeeae)小菇属(Myeena)[1],主要包括紫萁小菇(M . mosmundicola)、石斛小菇(M . mdendrobii)和开唇小菇(M . manoectochila)三大类属。天麻萌发菌主要分布于我国陕西、云南、甘肃、河南、河北、湖南、湖北、贵州、新疆、四川、安徽、黑龙江、吉林、广西、海南、福建、内蒙古、西藏及台湾等省区[2],为天麻(Gastrodia elata)有性繁殖时的必要共生菌。天麻种子细小如粉尘,无胚乳及其营养贮备,无外源营养供给,种子不能发芽。它的萌发靠小菇属(Mycena)一类真菌菌丝侵染种胚提供营养,促使其萌发[3]。随着天麻药用需求不断增加,野生资源日益匮乏,且人工栽培中以无性繁殖来连续种植,天麻球茎明显退化,侧芽发芽率低等原因,经过有性繁殖培育0代天麻或1代天麻,再通过无性繁殖扩大种植已成为天麻稳产和高产的重要途径[4-5]。目前,国内外对天麻萌发菌种质资源研究报道较少,且主要集中于地区优良栽培品种的筛选和生物学特性的研究,如冉孝琴等对贵州天麻萌发菌优良菌株进行了筛选[6];李方安等对紫萁小菇生物学特性进行了初步研究[7]。
由于天麻萌发菌菌株市场上命名混杂,且物种间形态差异不大,通过形态鉴定较为困难。目前,秦巴山区天麻萌发菌菌种的亲缘关系尚无相关研究报道,萌发菌菌种的种源关系不明确,缺少适宜地域栽培生长的天麻萌发菌菌种,对于培育出适合秦巴山区栽培的优良高产菌种的难度较大。利用rDNA ITS序列分析技术[8],可以直接提取蕴含于基因组内的遗传信息,检测和比较传统形态学无法区分的相似菌株种间或种内差别,具有快速、灵敏、准确的优点。笔者选取秦巴山区8103-3、8103-4、8103-6、8103-7、8103-8、萌-3、萌-7、萌MD-2、萌发菌、石斛-1、石斛-2、萌-云南共12个天麻萌发菌主要栽培菌株,对其形态特征及核糖体转录间隔区(ITS)进行序列分析,并进一步对ITS序列进行核酸序列数据库GenBank同源性检索比对,构建系统发育树,以期为基于传统形态特征对秦巴山区天麻萌发菌菌株之间的亲缘关系提供分子依据,为天麻萌发菌建立种质资源库及优良菌种的选育提供依据。
1材料与方法
1.1材料
1.1.1供试菌株8103-3、8103-4、8103-6、8103-7、8103-8、萌-3、萌-7、萌MD-2、萌发菌、石斛-1、石斛-2、萌-云南共12株萌发菌菌株:由陕西省资源生物重点实验室食药用菌菌种保藏中心提供。
1.1.2改良CPDA培养基包括马铃薯200.0 g,玉米粉 50.0 g,葡萄糖20.0 g,KH2PO4 5.0 g,MgSO4·7H2O 3.0 g,NH4NO3 5.0 g,维生素B1 10 mg,琼脂 15.0 g,加水至1 000 mL,pH值自然。CPDA液体培养基,根据上述改良CPDA培养基配制,但不加添琼脂。
1.2方法
1.2.1拮抗试验制备试管斜面培养基,活化各菌株,每株3管;取试管活化菌株在同一培养皿上对称接种2个菌株,24 ℃ 恒温培养7 d,观察菌丝间的拮抗反应情况。
1.2.2萌发试验青冈木树叶浸泡24 h后,平铺在培养皿表面,接种各萌发菌菌株,每株3个,24 ℃ 恒温培养7 d,选取未开裂、颜色较深、富有弹性且大小相近的天麻果实,均匀播撒在培养皿中,24 ℃ 恒温培养60 d,期间定期定量加水,观察萌发情况。
1.2.3液体发酵制备CPDA液体培养基,分装于250 mL三角瓶中,每瓶装液量150 mL,于1.034×105 Pa灭菌 30 min,冷却后将活化的菌株接种至液体摇瓶中,静置24 h后,24 ℃、160 r/min振荡培养7 ~12 d。用3层无菌纱布过滤菌丝球,再用无菌水洗涤过滤2~3次,最后用无菌滤纸吸干水分收集菌丝体。4 ℃冰箱保存,备用。
1.2.4DNA的提取及扩增采用CTAB法提取供试菌丝体的基因组DNA[10];PCR扩增:ITS引物:ITS 1(5′-3′)TCCGTAGGTGAA-CCTGGG,ITS 4(5′-3′)TCCTCCGCTTATTGA-TATGC。PCR反应体系:2×Taq PCR Green Mix 15 μL,引物ITS 1、ITS 4各2 μL,模板DNA 2 μL,加灭菌双蒸水至30 μL。PCR扩增程序:94 ℃预变性5 min;94 ℃变性1 min,55 ℃ 退火45 s,72 ℃延伸1 min,30个循环;72 ℃延伸10 min。采用1%的琼脂糖凝胶进行45 min电泳。
1.2.5测序及构建系统进化树经琼脂糖凝胶电泳检测的上述PCR产物由上海生工生物技术服务有限公司进行双向测序。将所测的序列用Clustal-W软件进行对位排列,然后手工适当校正。随后将调整后的序列提交GenBank数据库并进行Blast检索,下载同源性较高的序列。利用Mega 6软件构建系统发育树。
2结果与分析
2.112株天麻萌发菌的拮抗反应
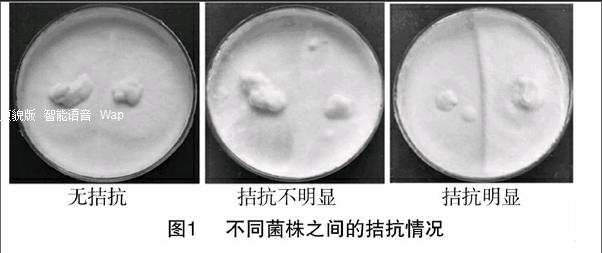
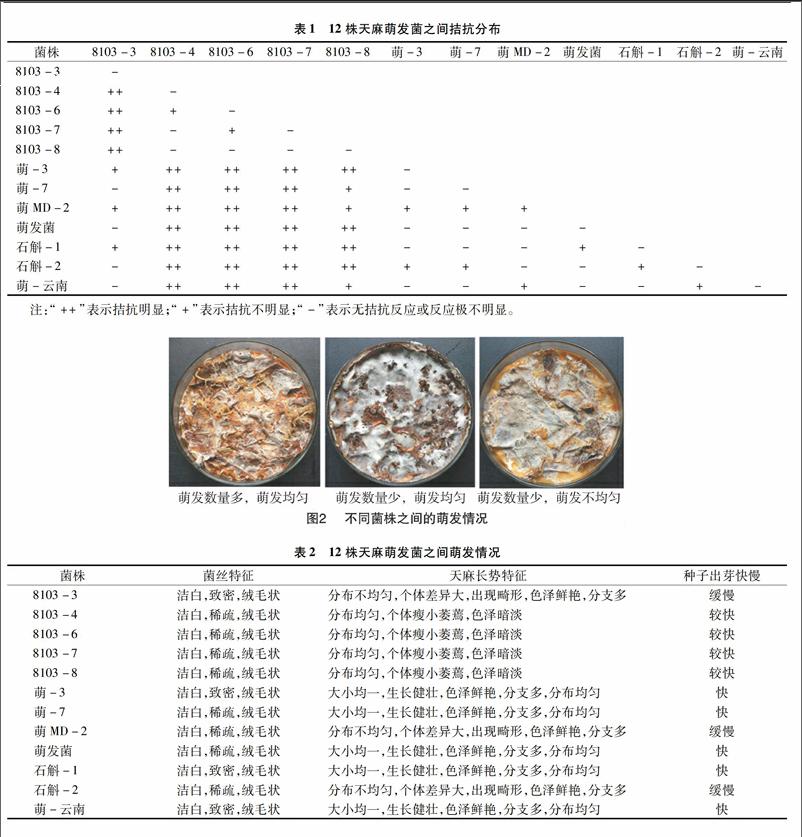
12株天麻萌发菌通过相互之间的两两接触性试验,结果见图1、表1,分析得出8103-4、8103-6、8103-7、8203-8之间无拮抗反应或反应不明显,与其他菌株之间拮抗反应明显,故此4株菌亲缘性相近;萌-3、萌-7、萌发菌、石斛-1、萌-云南之间无拮抗反应或反应不明显,与8103-3,萌MD-2,石斛-2之间拮抗反应微弱,故可将此8株菌分成2组来进一步研究。
2.212株萌发菌对天麻种子萌发研究
12株天麻萌发菌与天麻种子伴栽后萌发情况见图2、表2。萌-3、萌-7、萌发菌、石斛-1、萌-云南 5株菌中天麻种子萌发数量多,萌发均匀,天麻生长健壮,色泽鲜艳;8103-4、8103-6、8103-7、8203-8这4株菌中天麻种子萌发数量稀少或没有,天麻瘦小萎蔫,色泽暗淡;8103-3、萌MD-2、石斛-2中天麻种子萌发数量较少,天麻种子萌发不均匀,天麻生长良好,色泽鲜艳。
2.3供试菌株ITS-PCR扩增后电泳结果
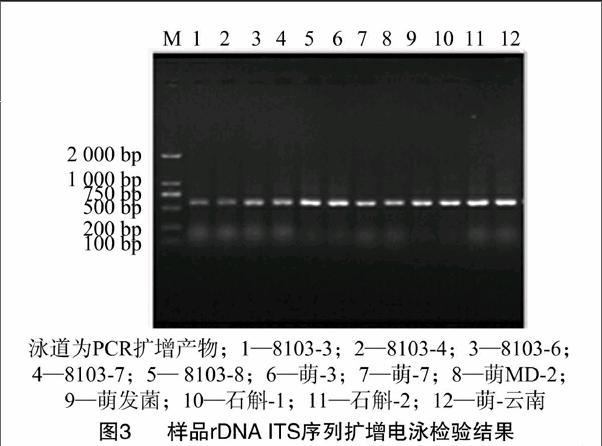
取5 μL PCR产物,用10 g/L琼脂糖凝胶(TBE缓冲液)电泳检测,于Tanon 3500R凝胶成像系统(中国TANON公司产品)拍照,电泳结果见图3。
由图3可见,12个天麻萌发菌菌株的基因组DNA经PCR扩增后获得了单一的条带,菌种的rDNA ITS区域的序列片段长度大小为600 bp。将所获得的12个ITS序列提交给NCBI公共数据GenBank进行比对,分析得到8103-3、8103-4、8103-6、8103-7、8103-8、萌-3、萌-7、萌MD-2、萌发菌、石斛-1、石斛-2、萌-云南收录号依次为:KT149154、KT149155、KT149156、KT149157、KT149158、KT149159、KT149160、KT149161、KT149162、KT149163、KT149164、KT149165。
2.3系统发育树分析
从GenBank上进行Blast检索,下载同源性较高的菌株序列,用Clustal-W软件对所有序列进行人工校正,再用MEGA 6采用临近相邻法聚类,经自举法检验(1 000次重复)构建系统发育树。
菌株菌丝特征天麻长势特征种子出芽快慢8103-3洁白,致密,绒毛状分布不均匀,个体差异大,出现畸形,色泽鲜艳,分支多缓慢8103-4洁白,稀疏,绒毛状分布均匀,个体瘦小萎蔫,色泽暗淡较快8103-6洁白,稀疏,绒毛状分布均匀,个体瘦小萎蔫,色泽暗淡较快8103-7洁白,稀疏,绒毛状分布均匀,个体瘦小萎蔫,色泽暗淡较快8103-8洁白,稀疏,绒毛状分布均匀,个体瘦小萎蔫,色泽暗淡较快萌-3洁白,致密,绒毛状大小均一,生长健壮,色泽鲜艳,分支多,分布均匀快萌-7洁白,稀疏,绒毛状大小均一,生长健壮,色泽鲜艳,分支多,分布均匀快萌MD-2洁白,稀疏,绒毛状分布不均匀,个体差异大,出现畸形,色泽鲜艳,分支多缓慢萌发菌洁白,稀疏,绒毛状大小均一,生长健壮,色泽鲜艳,分支多,分布均匀快石斛-1洁白,致密,绒毛状大小均一,生长健壮,色泽鲜艳,分支多,分布均匀快石斛-2洁白,稀疏,绒毛状分布不均匀,个体差异大,出现畸形,色泽鲜艳,分支多缓慢萌-云南洁白,致密,绒毛状大小均一,生长健壮,色泽鲜艳,分支多,分布均匀快8103-4、8103-6、8103-7、8103-8这4个菌在同一个分支上,亲缘关系密切,与已知序列相似度为99%,但在进化树上形成了单独的分支,这可能是新的分类单元;8103-3、萌-3、萌-7、萌MD-2、萌发菌、石斛-1、石斛-2、萌-云南这8个菌在同一个分支上,其中8103-3、萌发菌与标准菌Mycena citrinomarginat亲缘关系比较近,萌MD-2、石斛小菇-1与标准菌种Mycena purpureofusca亲缘关系非常近。
3讨论与结论
真菌的拮抗反应是体细胞不亲和的直接体现。在真菌界,体细胞不亲和现象普遍存在。此方法具有简单、快捷、直观等优点。是常用的真菌菌种鉴定方法;ITS rDNA区进化速率较快,在绝大多数真核生物中表现出极为广泛的序列多态性,对高等植物和大型真菌鉴定有很好的准确性和灵敏性,适用于不同真菌鉴定及系统发育分析。结合拮抗反应和ITS序列分析对天麻萌发菌进行亲缘关系分析具有极高的准确性。
本研究应用拮抗反应和萌发特性对萌发菌菌株进行研究,对其中菌丝粗壮,菌落致密,生长速度快萌发效果好的菌种进行标记筛选,通过对12株天麻萌发菌间的拮抗反应以及与天麻种子萌发试验的分析,大致将12株萌发菌分为3类:萌-3、萌-7、萌发菌、石斛-1、萌-云南,此5株菌株长势相同,萌发效果好,株长势相同,与其他菌株拮抗反应明显,萌发效果不理想;103-3、萌MD-2、石斛-2此3株菌与8103-4、8103-6、8103-7、 8103-8拮抗反应明显与其他菌株拮抗反应不明显,萌发效果较好。应用ITS序列分析技术,对秦巴山区天麻萌发菌菌种进行了测序分析,将其分为2类,其中8103-4、8103-6、8103-7、 8103-8这4个菌在同一个分支上,相似度为100%,亲缘关系相近,但在进化树上形成了单独的分支,形成了新的分类单元;萌-3、萌-7、萌MD-2、萌发菌、石斛小菇-1、石斛小菇-2、萌-云南8个菌在同一个分支上,其中8103-3、萌发菌与标准菌Mycena citrinomarginata亲缘关系比较近,萌MD-2、石斛-1、萌-3、萌-7、石斛-2、萌-云南与标准菌株Mycena purpureofusca亲缘关系较近。这与拮抗试验及萌发试验得到的结论大致相同。由于在实际生产种存在突变、变异、进化的原因,菌株间存在物种多样性的影响,对秦巴山区12个天麻萌发菌供试菌株进行RAPD和酯酶同工酶指纹图谱的分析还有待进一步研究。
参考文献:
[1] 徐锦堂. 中国天麻栽培学[M]. 北京:北京医科大学、中国医科大学联合出版社,1993.117-119
[2]谢果珍,申爱荣,谭著明,等. 天麻共生菌研究进展[J]. 湖南中医杂志,2015,31(4):206-208.
[3]冉孝琴,桂阳,张丽娜,等. 天麻种子自然萌发和生长特性[J]. 湖北农业科学,2014,53(10):2325-2329.
[4]高兴盛. 天麻有性繁殖的新方法[J]. 吕梁高等专科学校学报,2006,22(2):11-12.
[5]徐锦堂. 我国天麻栽培50年研究历史的回顾[J]. 食药用菌,2013,21(1):58-63.
[6]冉孝琴. 贵州天麻萌发菌优良菌株的筛选[D]. 贵阳:贵州师范大学,2014:3-18.
[7]李方安,潘燕,秦芸. 紫萁小菇生物学特性的初步研究[J]. 中国农学通报,2013,29(25):142-145.
[8]刘华晶,许修宏,姜廷波,等. 基于ITS序列分析探讨大兴安岭地区野生黑木耳菌株的遗传多样性[J]. 四川农业大学学报,2011,29(1):40-44.
[9]张平,谢娜,赖腾强. 无孢灵芝菌丝生物学特性研究[J]. 河北北方学院学报:自然科学版,2014,30(5):36-40.
[10]刘丽,张永军,许长征,等. 一种改良的CTAB法提取产多糖真菌DNA[J]. 中国生物工程杂志,2014,34(5):75-79.石杜娟,安正弼,徐伟韦,等. 基于Ebb & Flow灌溉系统的不同浓度营养液对蝴蝶兰生长和开花的影响[J]. 江苏农业科学,2016,44(5):249-252.
