基于籼粳杂交重组自交系的水稻籽粒性状的遗传变异研究
2016-07-14高维维陈立凯胡朝旭黄钰婷陈志强
高维维,陈立凯,胡朝旭,黄钰婷,黄 明,郭 涛,王 慧,陈志强
(华南农业大学 国家植物航天育种工程技术研究中心,广州 510642)
基于籼粳杂交重组自交系的水稻籽粒性状的遗传变异研究
高维维,陈立凯,胡朝旭,黄钰婷,黄明,郭涛,王慧,陈志强
(华南农业大学 国家植物航天育种工程技术研究中心,广州510642)
为研究多环境条件下籽粒性状的遗传变异及性状间相关性,解析籽粒性状遗传规律。以细长型籼稻品种‘玉针香’和短圆型粳稻品种‘02428’为亲本构建的含192个株系的RIL(重组自交系)群体为研究材料,在4种环境下分别对籽粒相关的16个性状进行相关性分析和遗传变异解析。结果表明,两亲本的粒型、垩白性状存在显著差异,其中,‘玉针香’精米长宽比、垩白粒率和垩白度分别为4.03、1.00%和0.18%,‘02428’精米长宽比、垩白粒率和垩白度分别为 1.45、87.40%和52.21%。RIL群体各性状的平均值介于两亲本之间,分布频率呈趋于单峰的正态分布,符合数量性状遗传规律。多环境下变异分析表明,大部分籽粒性状受环境及环境与株系互作不同程度的影响;而谷粒面积、谷粒圆度2个性状受环境、环境与株系互作的影响均不显著。遗传群体各性状变异系数为6.34%~84.55%,广义遗传力为4.26%~70.67%。粒长、粒宽、垩白粒率、垩白度间呈明显互作关系。在RIL群体中筛选获得4个在穗部性状、粒质量和稻米外观品质等方面综合表现较为优良的株系,具有较高育种价值。说明:基于遗传背景明确的重组自交系群体,可解析籽粒性状的遗传变异规律,并有效创制优异育种材料。
水稻;重组自交系;籽粒性状;遗传变异
水稻(OryzasativaL.)是中国最重要的粮食作物之一,与国家粮食安全密切相关[1]。随着国民经济的发展和人民生活水平的提高,人们对水稻产量和品质的要求逐步提高[2]。水稻籽粒性状包括粒长、粒宽、粒质量及垩白等,籽粒性状表现直接影响水稻产量和品质,籽粒性状的改良被认为是水稻育种的重要目标。研究水稻籽粒性状的遗传规律,有助于提高品种选育效率。
大量研究表明,水稻籽粒性状表现为典型的数量性状遗传[3-5]。符福鸿等[6]采用3×10NC正交设计研究表明,粒长与长宽比、千粒质量与粒宽之间均呈极显著正相关;粒长、粒宽和千粒质量之间相关性不显著。雷东阳等[7]采用‘IR75589-31S’‘IR60’‘IR70’和90个籼粳成分不同的株系进行不完全双列杂交,对杂交稻米外观品质遗传及性状间的相关性进行研究,认为粒长、粒宽和粒长宽比以加性效应为主,母体效应也起到一定的作用,垩白度主要受到加性效应和母体效应控制,同时还受到细胞质的影响;粒长、粒宽和粒长宽比的遗传力较高,垩白度的遗传力相对较低。
鉴于籽粒性状遗传的复杂性,RIL群体的构建及遗传解析得到高度重视。RIL群体中个体均为纯合体,利用RIL群体可以在多个不同的环境下进行性状评价,在数量性状研究上更准确地估计遗传变异组分[8]。林志强等[9]利用水稻长穗大粒品系‘FJCD’与籼型三系恢复系‘IR24’配制杂交组合,构建含178 个株系的RILs 群体,并在不同环境下进行主要品质性状遗传分析,结果表明,在不同生态条件下稻米主要品质性状间的相关性有82.22%的比率表现较为一致,不因环境的变化而改变其相关性。同时,利用RIL群体也定位大量与籽粒性状有关的QTL位点[10-14]。 进一步构建不同遗传背景的RIL群体,解析不同环境下籽粒性状的遗传表现,对于籽粒性状QTL位点鉴定和育种材料创制具有理论和实践意义。
本研究以籽粒性状具有显著差异的籼稻品种‘玉针香’和粳型种质‘02428’为亲本构建重组自交系群体,调查4种环境下各亲本及群体的单株穗质量、有效穗数、千粒质量、谷粒长、谷粒宽、谷粒长宽比、谷粒面积、谷粒周长、谷粒圆度、精米长、精米宽、精米长宽比、垩白粒率、垩白面积、垩白度和精米白度,比较分析水稻籽粒性状的遗传力,并对不同性状进行相关性分析,以期为开展籽粒性状QTL定位和育种材料创制奠定基础。
1材料与方法
1.1试验材料
‘玉针香’/‘02428’重组自交系(RIL)群体以及母本‘玉针香’(P1)和父本‘02428’(P2)。其中,‘玉针香’为湖南省审定的优良籼稻品种,‘02428’为江苏省选育的优良广亲和粳型种质。RIL群体采用单粒传的方法构建,世代为F5~F6,含192个株系。
1.2试验方法与性状调查
自交系群体和2个亲本分别于2014年(晚季)、2015年(早季)在华南农业大学国家植物航天育种工程技术研究中心育种基地(广州天河、广州增城两地)种植。2014年广州天河试验基地简称环境1(E1)、2014年增城试验基地简称环境2(E2)、2015年广州天河试验基地简称环境3(E3)、2015年增城试验基地简称环境4(E4)。2014年晚季:7月28日播种,8月10日移栽;2015年早季:3月7号播种,4月1日移栽。试验采用随机区组设计,2个重复,各小区中每个株系及亲本均种植3 行,每行6株。株行距为20 cm×20 cm,单本栽插。田间管理(水、肥、病虫害防治等)按当地大田常规栽培要求实施。为保证谷粒含水量一致,成熟时收获烘干再进行有关性状测定。
参考《农作物品种区域试验技术规范 水稻》(NY/T 1300-2007),考查单株穗质量(Panicle mass per plant)、每株有效穗数(Effective panicle number)、千粒质量(1 000-grain mass)等性状。采用SmartGrain软件(http://www.nias.affrc.go.jp/qtl/SmartGrain/)并参照操作说明测量谷粒长(Grain length)、谷粒宽(Grain width)、谷粒长宽比(Length-to-width ratio of grain)、谷粒周长(Perimeter length of grain)、谷粒面积(Area size of grain)和谷粒圆度(Circularity)等指标,分析模式选择批量分析[batch analysis]。采用JMWT12 大米外观品质检测仪测量精米长(Milled rice length)、精米宽(Milled rice width)、精米长宽比(Length-to-width ratio of milled rice)、精米白度(Whiteness)、垩白粒率(Chalky rice rate)、垩白面积(Chalky area)以及垩白度(Chalkiness degree)。
1.3数据分析
采用Excel进行性状的基本统计分析,采用OriginPro 8.0绘制各性状测得数据平均值的频次分布图,采用SAS 9.3软件进行描述性统计分析、方差分析和遗传参数的估算。
2结果与分析
2.1亲本及RIL群体穗部和籽粒外观性状分析
采用2年2点的数据平均值对亲本及192个RIL分离群体的16个性状进行统计分析(表1),结果表明,4种环境下2亲本各性状均存在明显差异,其中,粒型、垩白性状存在极端差异(图1):‘玉针香’为细长粒,精米长宽比达到4.03,而‘02428’为椭圆粒,精米长宽比为1.45;‘玉针香’的垩白粒率和垩白度分别为1.00% 和0.18%,而‘02428’分别为87.40%和52.21%,差异极明显。另外,‘玉针香’单株穗质量和千粒质量分别为46.5 g和28.88 g,比‘02428’分别高17.6 g和 4.28 g。‘玉针香’的有效穗数、谷粒长、谷粒面积、谷粒周长、谷粒长宽比、精米长、精米长宽比均高于‘02428’。
群体各性状测得数据的平均值均介于两亲本之间,其中,单株穗质量、有效穗数、千粒质量等9个性状出现明显超亲分离(表1)。16个性状中,除垩白度外,其他各性状的偏度和峰度的绝对值均小于1,表明这些性状的观测值在群体中呈对称、均匀分布。各性状的变异系数为6.34%~84.55%。粒型、粒质量的变异系数均小于10%,而垩白、穗质量、穗数性状的变异系数较大。垩白粒率变异系数为84.55%;垩白面积和垩白度的变异系数次之,分别为28.17%和63.92%;单株穗质量、有效穗数的变异系数亦较大,分别为26.55%和21.49%,表明控制这些性状的基因在数量和效应上较为复杂。

表1 4种环境下亲本和RIL群体穗部和籽粒性状的表型变异分析

A.‘玉针香’和‘02428’的谷粒形态Grains of ‘Yuzhenxiang‘ and ‘02428’;B.‘玉针香’和‘02428’的精米籽粒形态Kernels of ‘Yuzhenxiang’ and ‘02428’
图1‘玉针香’和‘02428’的籽粒性状表现
Fig.1The grain traits of ‘Yuzhenxiang’ and ‘02428’
籼稻亲本‘玉针香’的稻穗比粳型种质‘02428’更长,而着粒密度则低于‘02428’,其衍生的重组自交系产生极其广泛的穗型变异分离。由图2-A可观察到,不同株系间穗部性状存在明显的差异,涵盖长穗低着粒密度、长穗高着粒密度、短穗低着粒密度、短穗高着粒密度以及中间型等各种类型。
粒长、粒宽是粒型的代表性指标,‘玉针香’籽粒细长、低垩白,而‘02428’籽粒宽短、高垩白。由表型变异分析结果可知,重组自交系后代的粒长性状未出现超亲分离,而精米粒宽则存在比亲本‘玉针香’更细的后代。与此对应,分析发现粒宽的变异系数比粒长更高,这与以往许多研究大不相同,表明本试验材料可能存在更多调控粒宽性状的微效基因,而且显隐性更为复杂。图2-B为不同重组自交株系的籽粒表型,由图2-B可知,群体后代的籽粒分离出短、小型,短、圆型,细、长型,短、宽型等不同类型。
2.2各性状在不同环境的比较及互作
数量性状往往受环境影响较大。为此,本研究采用一般线性模型分析籽粒主要性状与环境的作用,结果表明(表2),环境对垩白率、垩白度和精米白度的影响达到极显著水平,对谷粒周长影响达到显著水平;其中,垩白度和精米白度受环境影响最大,F值分别为62.66和81.44。环境与株系互作对千粒质量、谷粒周长、精米长、精米宽、精米长宽比和垩白率的影响达到极显著水平,说明籽粒这些性状受到遗传因素和环境因素的共同作用。谷粒面积、谷粒圆度2个性状受环境、环境与株系互作的影响均不显著,多环境下均值变异幅度小。

A.不同重组自交株系的穗部表型Phenotypes of panicles of different RILs;B.不同重组自交株系的籽粒表型Phenotypes of kernels of different RILs

图2 重组自交系群体穗部和籽粒的表型变异
注:E1~E4.4个鉴定环境。*和**表示显著水平分别为P<0.05和P<0.01。下同。
Note:E1-E4.Four environments.Value with * and ** mean differ significantly atP<0.05 andP<0.01,respectively.The same as below.
2.3籽粒性状在RIL群体中的分离
基于RIL群体中的16个目标性状在4种环境下的平均值,对这些性状的分布进行统计分析,性状分布频次见图3(横坐标为RIL群体的性状平均值,纵坐标为RIL个数)。频次图表明,这些性状群体均值与亲本均值相近,分离模式呈连续变异。除垩白粒率外,其他各性状分布频率接近单峰的正态分布,符合典型的数量性状遗传规律,受多基因控制。

图3 籽粒性状在RIL群体的分布
2.4性状的遗传变异规律
4种环境下,采用SAS 9.3中GLM(General linear models)对RIL群体16个性状进行方差分析,结果表明,各株系间均存在显著差异,除有效穗数的P值<0.01,其他P值均<0.001(表3)。垩白粒率、垩白面积和垩白度的表型变异系数较大,分别为98.38%、46.32%和94.22%;单株穗质量和有效穗数的表型变异系数分别为43.91%和39.29%;其他性状的表型变异系数较小,为7.72%~15.17%。在遗传变异系数方面,垩白粒率和垩白度的变异系数也较大,分别为73.47%和43.71%;垩白面积、单株穗质量、谷粒长宽比和精米长宽比的遗传变异系数分别为16.52%、14.57%、12.75%和11.98%;其他性状的遗传变异系数较小,为5.06%~9.23%。
群体遗传参数结果表明,各性状的广义遗传力为4.26%~70.67%,其中谷粒长、谷粒宽、谷粒长宽比、谷粒周长、谷粒圆度、精米长、精米宽、精米长宽比的广义遗传力介于59.33%和70.67%,受基因型控制为主。有效穗数广义遗传力最低,为4.26%,表明该性状的表型变异受环境及随机误差的影响较大。
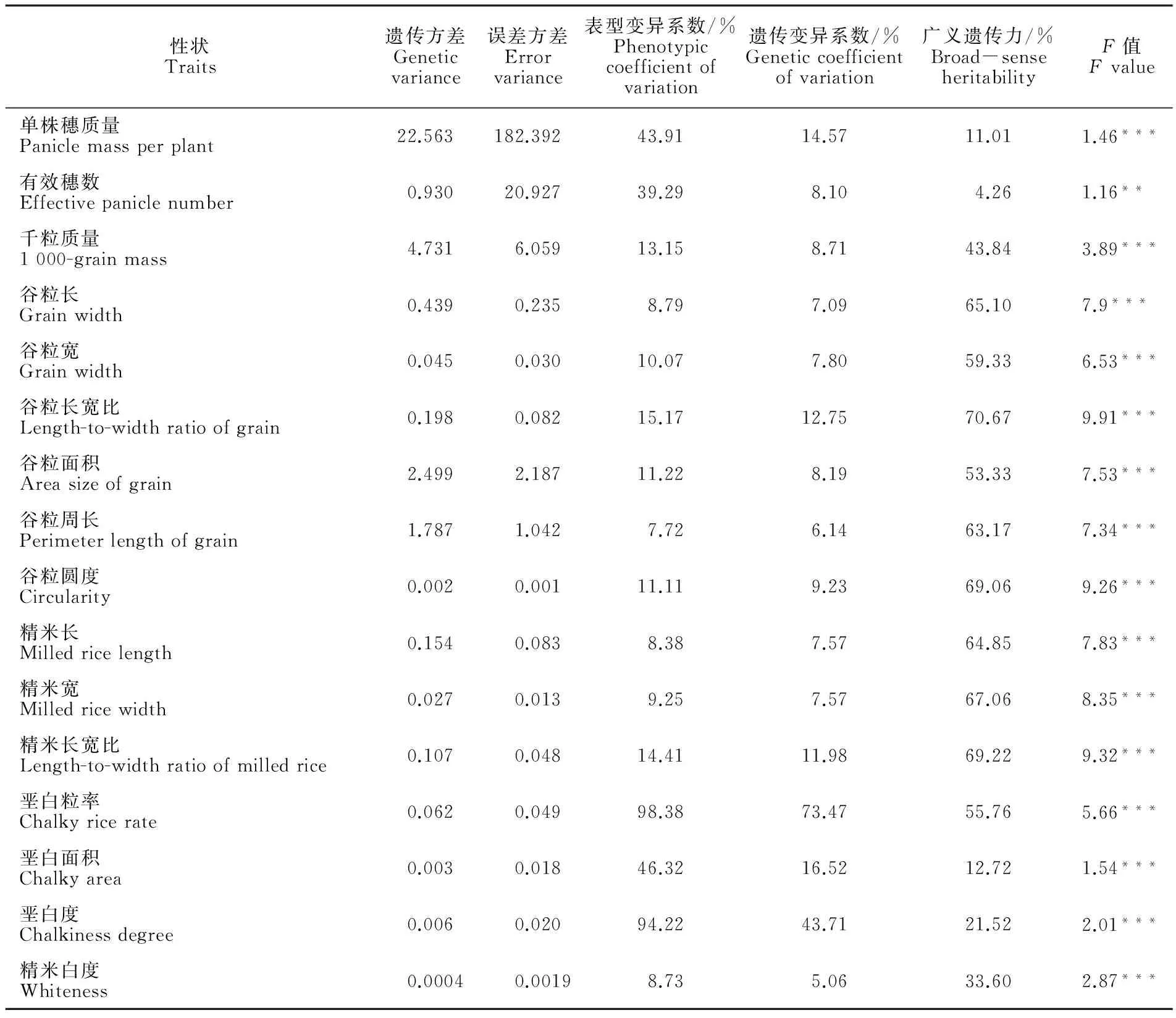
表3 重组自交系群体籽粒性状遗传参数
2.5不同性状间相关性分析
采用SAS统计软件对4种环境下RIL群体各性状表型值进行相关分析,发现多个性状间存在不同程度的相关(表4)。
单株穗质量与有效穗数、千粒质量、精米长呈极显著正相关,与精米长宽比呈显著正相关,与垩白粒率呈显著负相关,与精米白度呈极显著负相关,与其他性状的相关性不显著。粒质量是产量构成因子,其与精米粒长和精米粒宽呈显著正相关,且系数达到0.45以上,表明粒质量受粒型影响较大。同时,粒质量与垩白3个性状指标均达到极显著正相关。
籽粒外观性状方面,粒长、粒宽与垩白粒率、垩白度呈明显互作网络关系。首先粒长与粒宽表现负相关,相关系数r=-0.351;粒长与垩白性状存在显著负相关,与垩白粒率和垩白度的相关系数分别为-0.193和-0.114;而粒宽与垩白性状则显著正相关,与垩白粒率、垩白面积和垩白度相关系数分别为0.644、0.181和0.432。此外,白度与垩白粒率和垩白度呈显著正相关,并且相关系数均较大。
2.6综合性状优良株系的筛选
对4种环境下192个重组自交系的性状表现进行筛选,获得4个综合性状优良的株系,分别为GL52、GL53、GL106和GL117,其主要性状值见表5。这些株系的单株穗质量34.35~46.23 g,有效穗数为13~17,千粒质量均超过21 g,精米长宽比均大于2.7,垩白粒率均低于2%。表明这些株系在穗部性状、粒质量和稻米外观品质等方面综合表现较为优良,可用于育种实践。
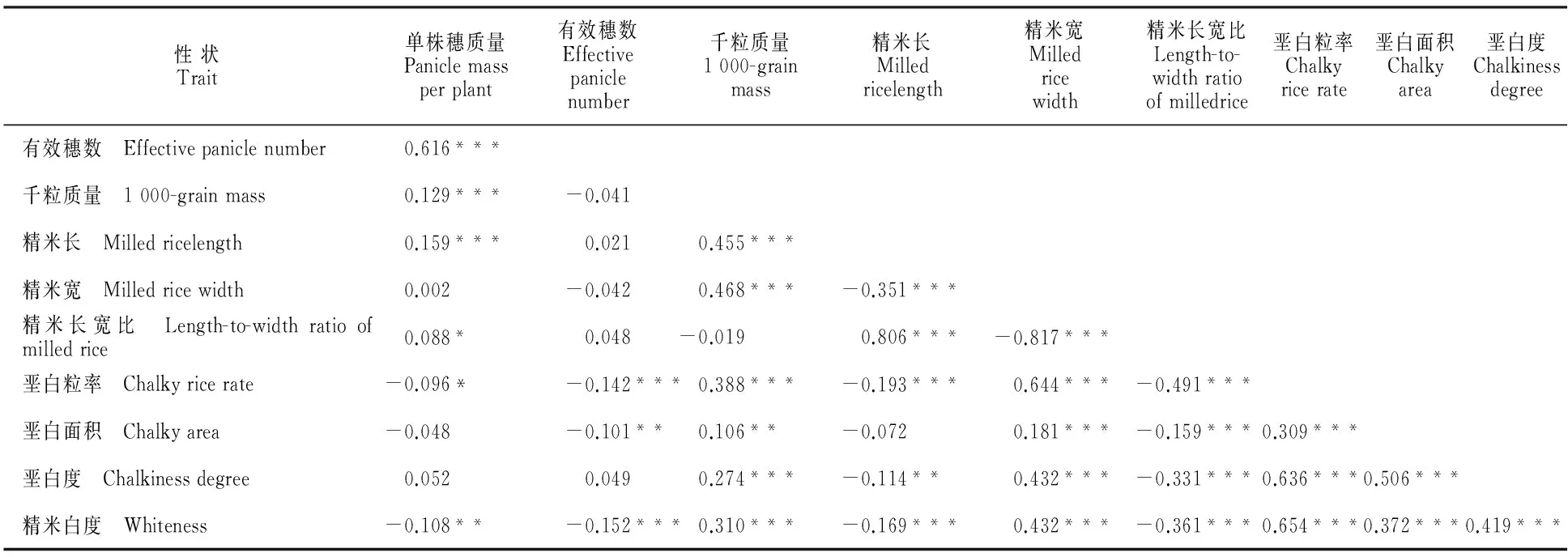
表4 RIL群体籽粒性状间的相关系数及其显著性

表5 4个优良株系在4种环境下主要性状平均值
3讨 论
多数研究表明,稻米籽粒性状属于数量性状,受环境影响较大,应在多种环境下进行鉴定才能正确评价性状表现[15]。对于数量遗传研究,理想的作图群体应具备系间性状变异大、系内性状稳定的典型特征[16]。而 RIL群体为稳定群体,可在多个环境下进行重复试验,因此可以更好地进行多环境下的鉴定和遗传分析。本研究中,RIL群体各性状遗传变异分析及描述性统计分析表明,各性状在不同环境下均存在较广泛的变异,变异系数为6.34%~84.55%,表明该群体符合理想RIL作图群体的标准。从各性状的频次分布图可以看出,多数性状表现出双向超亲分离,在后代群体中的分离表现基本一致,频次分布图呈正态分布,而后代群体表现为正态分布是数量性状的遗传特点[17],说明这些性状在后代群体中的遗传是受微效多基因的数量性状座位控制的,可以进行遗传图谱的构建和QTL定位。
进一步评价不同性状的广义遗传力,本研究发现谷粒长、谷粒宽、谷粒长宽比、谷粒周长、谷粒圆度、精米长、精米宽、精米长宽比、千粒质量、垩白粒率等主要受遗传效应控制,粒长、粒宽、长宽比遗传力较高,垩白率遗传力略低,这与前人研究结果基本一致[18-20]。而垩白度、单株穗质量、有效穗数受环境影响大,遗传力较低,这与前人研究结果存在一定差异,张利华等[21]选择米质不同的7 个不育系和7 个恢复系,采用NC Ⅱ设计,于2000 年冬至2001 年春在海南进行人工杂交并分析稻米外观品质遗传力,结果表明,垩白率、垩白面积、垩白度的广义遗传力较高,达到90%以上;本研究采用的材料为重组自交系群体,其个体均为纯合体,可在多个不同的环境下对遗传力进行估计,而不育系与恢复系杂交后代可能会发生分离,适合在同一环境下进行遗传分析,因此结果可能与本研究结果不一致。
已有研究表明,可采用RIL群体不同性状间的表型相关系数鉴定群体中各株系的表现[22]。本研究中,单株穗质量与有效穗数、千粒质量、精米长呈极显著正相关,与精米长宽比呈显著正相关,与垩白粒率呈显著负相关,与前人研究结果相符[23-24]。此外,本研究发现,垩白率与精米白度呈极显著正相关,与赵国珍等[25]对垩白度与精米白度的描述相符。因此进行育种选择时可以同时选择高单株穗质量、高精米长宽比、低垩白、低精米白度的优良株系。通过筛选不同环境性状下的表现值,本研究获得4个综合性状优良的株系,在单株穗质量、千粒质量、精米长宽比、垩白率、垩白度、精米白度等指标等方面均得到改良,具有较高的育种价值。
本研究构建的192个重组自交系材料遗传背景明确,各性状在不同环境下均存在较广泛的变异,且谷粒长、谷粒宽、谷粒长宽比、谷粒周长、谷粒圆度、精米长、精米宽、精米长宽比、千粒质量、垩白粒率等主要受遗传效应控制。本研究将进一步利用该群体进行高遗传力性状的QTL定位研究,为水稻籽粒性状分子改良提供理论参考。
Reference:
[1]邓兴旺,王海洋,唐晓艳,等.杂交水稻育种将迎来新时代[J].中国科学(生命科学),2013,43(10):864-868.
DENG X W,WANG H Y,TANG X Y,etal.Hybrid rice breeding welcomes a new era of molecular crop design[J].ScientiaSinica(Vitae),2013,43(10):864-868(in Chinese with English abstract).
[2]CALINGACION M,LABORTE A,NELSON A,etal.Diversity of global rice markets and the science required for consumer-targeted rice breeding[J].PLoSOne,2014,9(1):e85106.
[3]石春海,申宗坦.早籼粒形的遗传和改良[J].中国水稻科学,1995,9(1):27-32.
SHI CH H,SHEN Z T.Inheritanceand improvement of grain shape in indica rice[J].ChineseJournalofRiceScience,1995,9(1):27-32(in Chinese with English abstract).
[4]MCKENZIE K S,RUTGER J N.Genetic analysis of amylose content,alkali spreading score,and grain dimensions in rice[J].CropScience,1983,23(2):306-313.
[5]WU P,ZHANG G,HUANG N.Identification of QTLs controlling quantitative characters in rice using RFLP markers[J].Euphytica,1996,89(3):349-354.
[6]符福鸿,王丰,黄文剑,等.杂交水稻谷粒性状的遗传分析[J].作物学报,1994,20(1):39-45.
FU F H,WANG F,HUANG W J,etal.Genetic analysis on grain characters in hybrid rice[J].ActaAgronomicaSinica,1994,20(1):39-45(in Chinese with English abstract).
[7]雷东阳,谢放鸣,陈立云.杂交水稻稻米外观品质性状间相关性及遗传分析[J].农业现代化研究,2010,31(2):212-215.
LEI D Y,XIE F M,CHEN L Y.Genetic and correlation analysis of grain appearance quality in hybrid rice[J].ResearchofAgriculturalModernization,2010,31(2):212-215(in Chinese with English abstract).
[8]YANG G,LI Y,WANG Q,etal.Detection and integration of quantitative trait loci for grain yield components and oil content in two connected recombinant inbred line populations of high-oil maize[J].MolecularBreeding,2012,29(2):313-333.
[9]林志强,郑燕,许旭明,等.水稻长穗大粒RIL群体主要品质性状的遗传分析[J].热带作物学报,2010,31(11):1881-1889.
LIN ZH Q,ZHENG Y,XU X M,etal.Genetic analysis of main grain quality traits of rice on an RIL population of long panicle and big grain character[J].ChineseJournalofTropicalCrops,2010,31(11):1881-1889(in Chinese with English abstract).
[10]邢永忠,谈移芳,徐才国,等.利用水稻重组自交系群体定位谷粒外观性状的数量性状基因[J].植物学报,2001,43(8):840-845.
XING Y ZH,TAN Y F,XU C G,etal.Mapping quantitative trait loci for grain appearance traits of rice using a recombinant inbred line population[J].ActaBotanicaSinica,2001,43(8):840-845(in Chinese with English abstract).
[11]BAI X,LUO L,YAN W,etal.Genetic dissection of rice grain shape using a recombinant inbred line population derived from two contrasting parents and fine mapping a pleiotropic quantitative trait locus qGL7[J].BMCGenetics,2010,11:16.
[12]晁园,冯付春,高冠军,等.利用重组自交系群体定位水稻品质相关性状的QTL[J].华中农业大学学报,2012,31(4):397-403.
CHAO Y,FENG F CH,GAO G J,etal.Mapping QTLs related with rice qualities of appearance,cooking and eating using a recombinant inbred line population[J].JournalofHuazhongAgriculturalUniversity,2012,31(4):397-403(in Chinese with English abstract).
[13]SHANMUGAVADIVEL P S,MITHRA S V A,DOKKU P,etal.Mapping quantitative trait loci (QTL) for grain size in rice using a RIL population fromBasmati×indicacross showing high segregation distortion[J].Euphytica,2013,194(3):401-416.
[14]杨占烈,戴高兴,翟荣荣,等.多环境条件下超级杂交稻协优9308重组自交系群体粒形性状的QTL分析[J].中国水稻科学,2013,27(5):482-490.
YANG ZH L,DAI G Y,ZH R R,etal.QTL analysis of rice grain shape traits by using recombinant inbred linesfrom super hybrid rice Xieyou 9308 in multi-environments[J].ChineseJournalofRiceScience,2013,27(5):482-490(in Chinese with English abstract).
[15]闫殿海,王金明.数量性状的主效和微效基因检测方法研究[J].现代农业科技,2012(8):11-15.
YAN D H,WANG J M.Research on detection method of major gene and minor gene of quantitative trait[J].ModernAgriculturalScienceandTechnology,2012(8):11-15(in Chinese with English abstract).
[16]GOUESNARD B,REBOURG C,WELCKER C,etal.Analysis of photoperiod sensitivity within a collection of tropical maize populations[J].GeneticResourcesandCropEvolution,2002,49(5):471-481.
[17]石培春,王光利,王小国,等.小麦RIL群体中籽粒淀粉性状的分离与主要淀粉性状的相关分析[J].新疆农业科学,2011,48(10):1789-1794.
SHI P CH,WANG G L,WANG X G,etal.The separation of starch properties of wheat grain among a RIL population and the correlation analysis among starch properties[J].XinjiangAgriculturalSciences,2011,48(10):1789-1794(in Chinese with English abstract).
[18]李仕贵,黎汉云,周开达,等.杂交水稻稻米外观品质性状的遗传相关分析[J].西南农业大学学报,1995,17(3):197-201.
LI SH G,LI H Y,ZHOU K D,etal.Genetic analysis on exterior quality traits in hybrid rice[J].JournalofSouthwestAgriculturalUniversity,1995,17(3):197-201(in Chinese with English abstract).
[19]刘勇.水稻籽粒性状的遗传分析及粒长基因定位[D].四川雅安:四川农业大学,2001.
LIU Y.Genetic analysis of grain traits and gene mapping of grain length in rice (Oryzasativa) [D].Ya’an Sichuan:Sichuan Agricultural University,2001 (in Chinese with English abstract).
[20]宗寿余,吕川根,邹江石.籼型两系杂交水稻稻米品质性状的配合力及遗传力分析[J].金陵科技学院学报,2007,23(3):47-52.
ZONG SH Y,LÜ CH G,ZOU J SH.Analysis on combining ability and heritability of appearance and milling quality traits in two-line indica hybrid rice[J].JournalofJinlingInstituteofTechnology,2007,23(3):47-52(in Chinese with English abstract).
[21]张利华,王林友,王建军.籼型杂交稻稻米碾磨品质与外观品质的配合力及遗传力研究[J].核农学报,2003,17(6):417-422.
ZHANG L H,WANG L Y,WANG J J.Studies on combing abilityand heritabilityof milling and physical propertiesin indica hybrid rice[J].ActaAgriculturaeNucleataeSinica,2003,17(6):417-422(in Chinese with English abstract).
[22]PERCY R G,CANTERLL R G,ZHANG J.Genetic variation for agronomic and fiber properties in an introgressed recombinant inbred population of cotton[J].CropScience,2006,46(3):1311.
[23]李贤勇,何永歆,李顺武,等.水稻千粒重与垩白粒率的相关性分析[J].西南农业学报,2003,16(4):20-23.
LI X Y,HE Y X,LI SH W,etal.Correlation between 1 000-grain weight and chalky rice rate of indica rice[J].SouthwestChinaJournalofAgriculturalSciences,2003,16(4):20-23(in Chinese with English abstract).
[24]林荔辉,吴为人.水稻粒型和粒重的QTL定位分析[J].分子植物育种,2003,1(3):337-342.
LIN L H,WU W R.Mapping of QTLs underlying grain shape and grain weight in rice[J].MolecularPlantBreeding,2003,1(3):337-342(in Chinese with English abstract).
[25]赵国珍,袁平荣,杨世准,等.异地栽培对云南粳稻育成品种品质性状的影响[J].生态环境学报,2009,18(4):1460-1466.
ZHAO G ZH,YUAN P R,YANG SH ZH,etal.Effects of different planting places on rice quality of the improved japonica rice cultivars from Yunnan province,China[J].EcologyandEnvironmentalSciences,2009,18(4):1460-1466(in Chinese with English abstract).
GUO Tao,male,professor,master supervisor.Research area: crop genetics and breeding,functional components.E-mail: guo.tao@vip.163.com.
(责任编辑:郭柏寿Responsible editor:GUO Baishou)
Genetic Variation of Grain Traits Based on Indica-Japonica Cross of RILs in Rice (Oryzasativa)
GAO Weiwei,CHEN Likai,HU Chaoxu,HUANG Yuting,HUANG Ming,GUO Tao,WANG Hui and CHEN Zhiqiang
(National Engineering Research Centre of Plant Space Breeding,South China Agricultural University,Guangzhou510642,China)
Using the RIL population derived from the Indica-Japonica cross,the genetic variation and correlation of rice grain traitsand other traitswere analyzed under multiple environments,and to provide efficient genetic dissection of grain traits in rice.A RIL (Recombinant Inbred Line) population with 192 lines developed from a cross between indica ‘Yuzhenxiang’(long grain) and japonica cultivars ‘02428’(short grain),was used in this study.The correlation and genetic variation analysis were conducted about 16 kernel traits under four environments.The result showed that a significant difference for grain shape and chalkiness traits was observed between the two parents.The kernel length-to-width ratio of ‘Yuzhenxiang’ was 4.03,the chalky rice rate was 1.00%,and the chalkiness degree was 0.18%,whereas,the kernel length-to-width ratio of ‘02428’ was 1.45,the chalky rice rate and chalkiness degree were 87.40% and 52.21%,respectively.The average values of the traits in RIL were between the two parents; all traits tended to be normal distribution with a single peak,and met with quantitative genetics.Analysis of genetic variability under different environment indicated that grain traits were vary affected by environment and their interactions,but influence of environments to grain area and circularity was non-significant.The coefficient variation was 6.34% to 84.55%,and the generalized heritability ranged from 4.26% to 70.67%.There was a significant interrelate relationship among kernel length,kernel width,chalky rice rate and chalkiness degree.Fourrecombinant inbred lines with fine performance in panicle traits,grain mass,and appearance quality traits were obtainedfrom the RIL population,which possessed superior utilization value in breeding.In conclusion,this study dissected the genetic variation of grain-related traits in rice,and developed elite lines for breeding efficiently.
Rice; Recombinant inbred line; Kernel traits; Genetic variation
2015-11-03
2016-01-21
The National Natural Science Foundation of China (No.31200250); National High Technology Research and Development Program of China (No.2012AA101201); International Science & Technology Cooperation Program of Guangzhou (No.2014J4500030); Modern Agro-industry TechnologyResearch System (No.CARS-01-12).
GAO Weiwei,female,master student.Research area: rice genetics and breeding,functional components.E-mail: 18819266044@163.com
CHEN Zhiqiang,male,professor,doctoral supervisor.Research area: crop genetics and breeding,functional components.E-mail: chenlin@scau.edu.cn
1004-1389(2016)07-0979-10
2015-11-03修回日期:2016-01-21
国家自然科学基金(31200250);国家高技术研究发展计划(863)(2012AA101201);广州市对外科技合作项目(2014J4500030);现代农业产业技术体系建设专项(CARS-01-12)。
高维维,女,硕士研究生,研究方向为水稻遗传育种。E-mail:18819266044@163.com
陈志强,男,教授,博士生导师,主要从事水稻遗传育种研究。E-mail:chenlin@scau.edu.cn
S511.01
A
网络出版日期:2016-06-30
网络出版地址:http://www.cnki.net/kcms/detail/61.1220.S.20160630.1624.010.html
郭涛,男,教授,硕士生导师,主要从事水稻遗传育种研究。E-mail:guo.tao@vip.163.com
