盐碱胁迫下不同水稻品种渗透调节物质及相关基因的变化
2016-06-15张婷婷杨美英王春红孙合美齐春艳侯立刚武志海
张婷婷,杨美英,王春红,孙合美,齐春艳,侯立刚,武志海
(1 吉林农业大学 a 生命科学学院,b 农学院,吉林 长春 130118;2 中国农业东北创新中心水稻研究所,吉林 公主岭 136102)
盐碱胁迫下不同水稻品种渗透调节物质及相关基因的变化
张婷婷1a,杨美英1a,王春红1a,孙合美1a,齐春艳2,侯立刚2,武志海1b
(1 吉林农业大学 a 生命科学学院,b 农学院,吉林 长春 130118;2 中国农业东北创新中心水稻研究所,吉林 公主岭 136102)
[摘要]【目的】 分析盐碱浓度、胁迫时间对不同水稻品种各龄苗有机渗透调节物质的影响及不同品种对盐碱胁迫响应的差异,为盐碱地改良水稻品种的选择及其敏感期防护提供理论依据。【方法】 选择耐盐碱水稻品种长白9号和普通栽培品种吉粳78为材料,研究3个盐碱浓度(10,40,80 mmol/L)、4个处理时间(24,48,72,84 h)对3种苗龄(30,45,70 d)水稻叶片脯氨酸、甜菜碱、肌醇含量及Δ1-吡咯琳-5-羧酸合成酶(P5CS)、甜菜碱醛脱氢酶(BADH1、BADH2)、肌醇单磷酸酶(IMP)基因表达量的影响,以正常水培的水稻为对照。【结果】 正常水培条件下长白9号叶片脯氨酸、甜菜碱含量高于吉粳78。盐碱胁迫条件下,长白9号和吉粳78叶片的脯氨酸、甜菜碱和肌醇含量均上升,P5CS、BADH1、BADH2、IMP基因表达量也高于对照。40 mmol/L NaCO3-NaHCO3处理较10和80 mmol/L处理更有利于植株积累渗透调节物质及上调相关基因的表达。长白9号渗透调节物质含量及其基因表达量的变化远大于吉粳78。盐碱胁迫24和48 h能使长白9号及时做出响应,并随着胁迫时间的延长甜菜碱、肌醇含量及BADH1、BADH2、IMP表达量持续升高,而吉粳78对盐碱胁迫响应较晚。2种水稻30,45 d龄苗渗透调节物质含量的变化及相关基因表达量的变化均比70 d明显。【结论】 长白9号耐盐碱的特性与其植株体内渗透调节物质丰富且对盐碱胁迫响应快的生理现象有关。2种水稻对盐碱胁迫具有一致的生理响应敏感阈值,在耐受条件下,盐碱地种植水稻更适宜大苗移栽。
[关键词]水稻;盐碱胁迫;渗透调节物质;脯氨酸;甜菜碱;肌醇
根据中国科学院的统计,分布在我国西北、东北及滨海地区的盐碱荒地和盐碱障碍耕地总面积超过3 333.3 万hm2,其中具有农业利用潜力的近1 333.3 万hm2,占我国耕地总面积的10%以上[1]。因此,合理有效地利用盐碱土,扩大作物种植面积,增加粮食产量已成为共识。
土壤盐碱化严重影响种子萌发和幼苗生长,使粮食和牧草产量、质量大幅度下降[2]。盐碱对植物的伤害主要为渗透胁迫和离子毒害2种。为了维持正常的细胞生理功能和渗透压,植物体有明显的渗透调节生理响应机制,其中脯氨酸、甜菜碱、肌醇和可溶性糖等是植物体在逆境胁迫条件下产生的主要有机渗透调节物质[3]。植物体可通过鸟氨酸途径和谷氨酸途径合成脯氨酸,逆境条件下脯氨酸主要合成途径是后者。Δ1-吡咯琳-5-羧酸合成酶(P5CS)是谷氨酸合成途径中的限速酶,它是一个双功能酶,具有谷氨酸激酶和谷氨酸-γ-半醛脱氨酶活性,可催化脯氨酸合成的前2步反应,同时它又可以被其产物脯氨酸反馈抑制[4]。甜菜碱是甘氨酸的衍生物,被认为是最好的渗透调节剂,能保护细胞内蛋白质和代谢酶的活性,甚至有稳定膜的作用[5],由胆碱经2步氧化生成,这2步反应分别由胆碱单氧化酶和甜菜碱醛脱氢酶(BADH)催化。目前,BADH基因已先后从菠菜、高粱、甜菜等植物中分离和鉴定[6]。肌醇是经肌醇单磷酸酶(IMP)催化脱磷酸作用生成的[7],由于含有多个羟基,可以有效地维持植物细胞中的水活度,从而能够有效地避免和减少盐渍环境中植物的渗透脱水[8]。
水稻是强耐盐作物,全生育期内几乎处于淹水状态,通过灌水淋洗盐分,不仅可以防止盐分向地表积累,而且水稻根系分泌的有机酸还能中和土壤碱性,可改良土壤物理性状[9]。灌溉种植水稻已成为提高盐碱地区水稻生产能力和改善生态环境的最有效的途径之一[10]。但不同地区盐碱土类型及盐碱程度不同,吉林省西部地区受地质气候条件的影响,约有170万hm2土地属内陆苏打盐渍型盐碱土[11],兼有不同的盐化和碱化特征[12]。长白9号是由吉林省农业科学院水稻研究所选育而成的耐盐碱水稻品种,在pH 8.0~8.5和土壤含盐量0.3%的盐碱条件下表现正常[13]。杨春武[14]报道,水稻生理参数在48 h变化最明显。张丽丽等[15]对长白9号进行了10,20,30 mmol/L Na2CO3-NaHCO3处理,发现30 mmol/L时,其叶片都有积累渗透调节物质的反应。本研究以耐盐碱水稻品种长白9号和普通栽培品种吉粳78为材料,采用qPCR的方法,研究3个盐碱浓度、4个处理时间对3种苗龄水稻叶片脯氨酸、甜菜碱、肌醇含量及Δ1-吡咯琳-5-羧酸合成酶(P5CS)、甜菜碱醛脱氢酶(BADH1、BADH2)、肌醇单磷酸酶(IMP)基因表达量的影响,综合分析盐碱处理浓度、胁迫时间对不同水稻品种各苗龄有机渗透调节物质形成的影响,探讨不同品种、不同苗龄水稻渗透能力对盐碱胁迫的响应程度,为改良盐碱地水稻品种的选择及其敏感期防护提供理论依据。
1材料与方法
1.1供试材料的处理
选择耐盐碱水稻品种长白9号和普通栽培品种吉粳78为材料,2品种均由吉林省农业科学院提供。水稻种子过夜浸泡后在气候培养箱中进行萌发,昼夜温度分别为(30±0.5) ℃和(25±0.5) ℃,每天光照16 h。将萌发2 d的水稻种子摆放在固定于塑料桶(容量2 L)的纱布上进行水培,营养液根据国际水稻研究所(IRRI)的常规营养液配方配制[14],营养液用量以正好浸没种子为宜,每周更换一次。水培试验在吉林农业大学植物培养室中进行,每天光照11 h,昼夜温度分别为(22±0.5) ℃和(17±0.5) ℃,相对湿度为(60±5)%。
1.2盐碱处理及取样
将Na2CO3、NaHCO32种碱性盐用常规营养液分别配制成10,40和80 mmol/L 3个浓度梯度的溶液。在水稻苗龄30,45和70 d时,分别选取长势均匀的2品种水稻苗各36桶,随机分成4组,进行3个梯度的盐碱胁迫处理,同时设常规营养液培养处理对照(CK)。分别在胁迫处理24,48,72和84 h取叶片样品,一部分样品经液氮冷冻后,转移至-80 ℃冰箱保存,用于RNA的提取;其他样品在120 ℃的烘箱中杀青10 min,于80 ℃下烘干24 h,然后用粉碎机(屹立不锈钢50 g粉碎机)粉碎,过孔径 0.425 mm筛,用于脯氨酸[16]、甜菜碱[17]和肌醇[18]含量的测定。
1.3盐碱胁迫对水稻幼苗叶片基因表达的影响
1.3.1RNA的提取及cDNA的合成采用RNAiso Plus 试剂(TaKaRa,中国大连)提取叶片样品总RNA。采用20 μL的反转录体系合成cDNA,其中包括Total RNA 1 μL,0.5 μg/μL Oligo(dt)181 μL,2×ES Reaction Mix 10 μL,Easy Script TM RT 1 μL,RNase-free water 7 μL。反转录得到的cDNA用于实时荧光定量PCR(qPCR)分析。
1.3.2反转录产物的qPCR扩增根据GenBank中公布的P5CS、IMP、BADH1、BADH2基因序列(4个基因的GenBank登录号分别为AY574031.1、AB012107.1、AB001348.1、AB096083.1),设计引物。选用Actin为看家基因,其引物见文献[19]。本试验所用引物均由上海生物工程有限公司合成,序列详见表1。
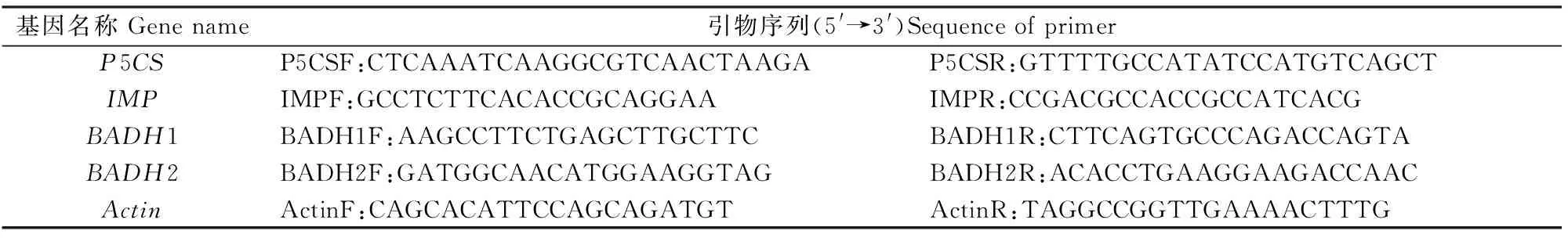
表 1 本试验供试PCR引物
采用TaKaRa 公司SYBR Premix ExTaqTMⅡ (TLi RNaseH Plus )试剂盒进行qPCR,反应体系为25 μL,其中包含SYBR®Premix ExTaqTM(2×) 12.5 μL、PCR Primers(10 μmol/L)各1.0 μL、ROX Reference Dye(50×) 0.5 μL、反转录cDNA模板2 μL,ddH2O 8 μL。反应于 Step One Plus PCR 仪上进行,每个样品3次平行反应,采用两步法PCR扩增标准程序扩增:95 ℃预变性30 s;95 ℃ 5 s,60 ℃ 30 s,循环40次。
通过扩增曲线和熔解曲线,确定引物的特异性。利用Step One 生物软件和Microsoft Excel、DPS分析处理试验结果,参照ABI公司提供的ΔΔCt方法计算相对表达量。
2结果与分析
2.1盐碱胁迫对水稻叶片渗透调节物质的影响
2.1.1脯氨酸盐碱胁迫对水稻叶片脯氨酸含量的影响结果见表2。由表2可以看出,长白9号CK样品脯氨酸含量略高于吉粳78 CK样品;盐碱处理2品种水稻不同苗龄幼苗叶片中脯氨酸含量与对照相比都有不同程度增加;长白9号3种苗龄样品脯氨酸含量都有较大变化,而吉粳78只有10 mmol/L Na2CO3-NaHCO3处理的30 d苗龄样品中脯氨酸含量变化明显,其他苗龄样品与CK相比脯氨酸含量的变化都小于长白9号。长白9号3种苗龄水稻脯氨酸含量均随着盐碱胁迫浓度的增加而增大,80 mmol/L Na2CO3-NaHCO3处理的脯氨酸含量达到最大值,并显著高于其他处理。45和70 d苗龄的吉粳78水稻叶片脯氨酸含量表现同样的变化规律,但在30 d苗龄时,10 mmol/L Na2CO3-NaHCO3处理的脯氨酸含量达到最大,之后随着处理浓度的增加脯氨酸含量逐渐下降。
2水稻品种各处理同一胁迫时间样品脯氨酸含量均随苗龄的增加呈现下降趋势。45和70 d苗龄2水稻品种各样品脯氨酸含量在胁迫84 h时达到最高,但苗龄为30 d时,长白9号和吉粳78各样品分别在胁迫48和84 h时达到最大值。

表 2 盐碱胁迫对水稻叶片脯氨酸含量的影响
注:同行数据后标不同小写字母表示差异显著(P<0.05)。下表同。
Note:Different lowercase letters in each row present significant difference atP<0.05 level.The same below.
2.1.2甜菜碱盐碱胁迫对水稻叶片甜菜碱含量的影响结果见表3。由表3可以看出,盐碱处理前长白9号水稻样品甜菜碱含量稍高于吉粳78。 Na2CO3-NaHCO3盐碱胁迫下2水稻品种各样品中甜菜碱含量均高于CK。
10 mmol/L Na2CO3-NaHCO3盐碱胁迫长白9号品种30和70 d苗及吉粳78 45和70 d苗24 h就能显著提高水稻叶片中的甜菜碱含量。长白9号、吉粳78品种3种苗龄叶片甜菜碱含量均在40 mmol/L Na2CO3-NaHCO3处理时显著增加,并达到最大值。
2水稻品种CK样品甜菜碱含量表现为胁迫30 d>70 d>45 d。30 d苗龄时,CK及相同浓度盐碱处理下长白9号样品甜菜碱含量大于吉粳78,且2品种均在84 h达到最大值;而45和70 d苗龄2品种水稻甜菜碱含量则都是在40 mmol/L Na2CO3-NaHCO3处理48 h时达到最大值。
2.1.3肌醇从表4看可以出,长白9号CK样品肌醇含量明显低于吉粳78 CK样品。与CK相比,盐碱胁迫可以使各苗龄水稻叶片中肌醇含量有不同程度提高,长白9号3个苗龄样品肌醇含量的变化都高于吉粳78。
3种苗龄长白9号水稻叶片肌醇含量均随着盐碱胁迫浓度的增大而增大,在40 mmol/L Na2CO3-NaHCO3处理达到最大值,之后呈下降趋势;45和70 d苗龄的吉粳78水稻叶片肌醇含量也在40 mmol/L达到最大值,而30 d苗龄吉粳78,40与80 mmol/L Na2CO3-NaHCO3处理样品肌醇含量差异很小。
2水稻品种CK肌醇含量都随苗龄的增加而减少,这与脯氨酸含量变化一致。45 d苗龄2水稻品种在盐碱胁迫24 h达到最大值,而其他苗龄并没有表现出明显的变化规律。

表 3 盐碱胁迫对水稻叶片甜菜碱含量的影响

表 4 盐碱胁迫对水稻叶片肌醇含量的影响
2.2盐碱胁迫对水稻幼苗叶片基因表达的影响
2.2.1P5CS基因表达量的差异盐碱胁迫对2种水稻叶片P5CS基因表达量的影响结果如图1所示。由图1-A可知,CK及Na2CO3-NaHCO3盐碱处理长白9号的P5CS基因表达量均高于相同处理下吉粳78。随着盐碱胁迫浓度的增大,2水稻品种P5CS基因表达量均有增加,且始终高于CK;40 mmol/L Na2CO3-NaHCO3处理下2水稻品种样品P5CS基因表达量达到最大值,

图 1盐碱胁迫对长白9号和吉粳78水稻叶片P5CS基因表达量的影响
A.苗龄30 d,处理时间48 h;B.苗龄45 d。下图同
Fig.1Effect of saline-alkali stress expressions ofP5CSgene in leaves of Changbai 9 and Jijing 78
A.Age 30 d,treat time 48 h;B.Treat time 45 d.The same below
由图1-B可知,受40 mmol/L Na2CO3-NaHCO3盐碱胁迫24 h 2品种水稻叶片中P5CS基因表达量下降;随着胁迫时间的延长P5CS基因表达量升高,但是2品种表现不同,长白9号叶片中P5CS基因表达量随着胁迫时间的延长而升高,于84 h达到最大值,而吉粳78P5CS基因表达量在胁迫72 h达到最大,84 h略有降低。
由图1-C可知,盐碱处理的不同苗龄2品种水稻P5CS基因表达量都高于CK,但随着苗龄的增加,长白9号品种CK及盐碱处理样品基因表达量持续降低;吉粳78水稻品种CK样品降低趋势明显,但处理45 d苗龄样品却略高于其他2苗龄样品。
2.2.2BADH基因表达量的差异由图2-A1、A2可以看出,Na2CO3-NaHCO3盐碱胁迫对BADH基因家族不同成员的影响不同;长白9号CK样品BADH1、BADH2基因表达量均高于吉粳78 CK。不同浓度Na2CO3-NaHCO3盐碱胁迫均能增加2品种BADH1、BADH2基因在叶片中的表达量,长白9号2基因表达量增加速度明显高于吉粳78,其中BADH1表达量增加幅度最大,而BADH2表达量的增加幅度明显低于BADH1。
图2-B1、B2显示,盐碱胁迫24 h能提高BADH1在2水稻叶片中的表达量,能降低BADH2在叶片中的表达量;从盐碱胁迫48 h开始,BADH2成员在2水稻叶片中的表达均上调,于84 h达到最大值,其中长白9号的变化比吉粳78明显。
从图2-C1、C2可以看出,2品种不同苗龄CK样品BADH1和BADH2表达量表现为苗龄30 d>70 d>45 d;Na2CO3-NaHCO3盐碱胁迫对30 d苗龄样品BADH1和BADH2基因表达量的影响明显大于70和45 d苗龄样品。
2.2.3IMP基因表达量的差异由图3-A可知,CK间相比,长白9号叶片中IMP基因的表达量低于吉粳78; Na2CO3-NaHCO3盐碱处理2品种水稻叶片IMP基因表达量表现出不同的变化趋势,随盐碱胁迫浓度的增加,长白9号叶中IMP基因表达量明显上调,于40 mmol/L时达到最大值,而吉粳78变化不明显。
由图3-B可知,长白9号受盐碱胁迫24 h就能作出上调IMP基因表达的响应,48 h达到最大值,而吉粳78样品在盐碱胁迫72 h时IMP的表达量才高于对照,84 h达到最大值;盐碱处理下长白9号IMP基因表达量变化值始终大于吉粳78。
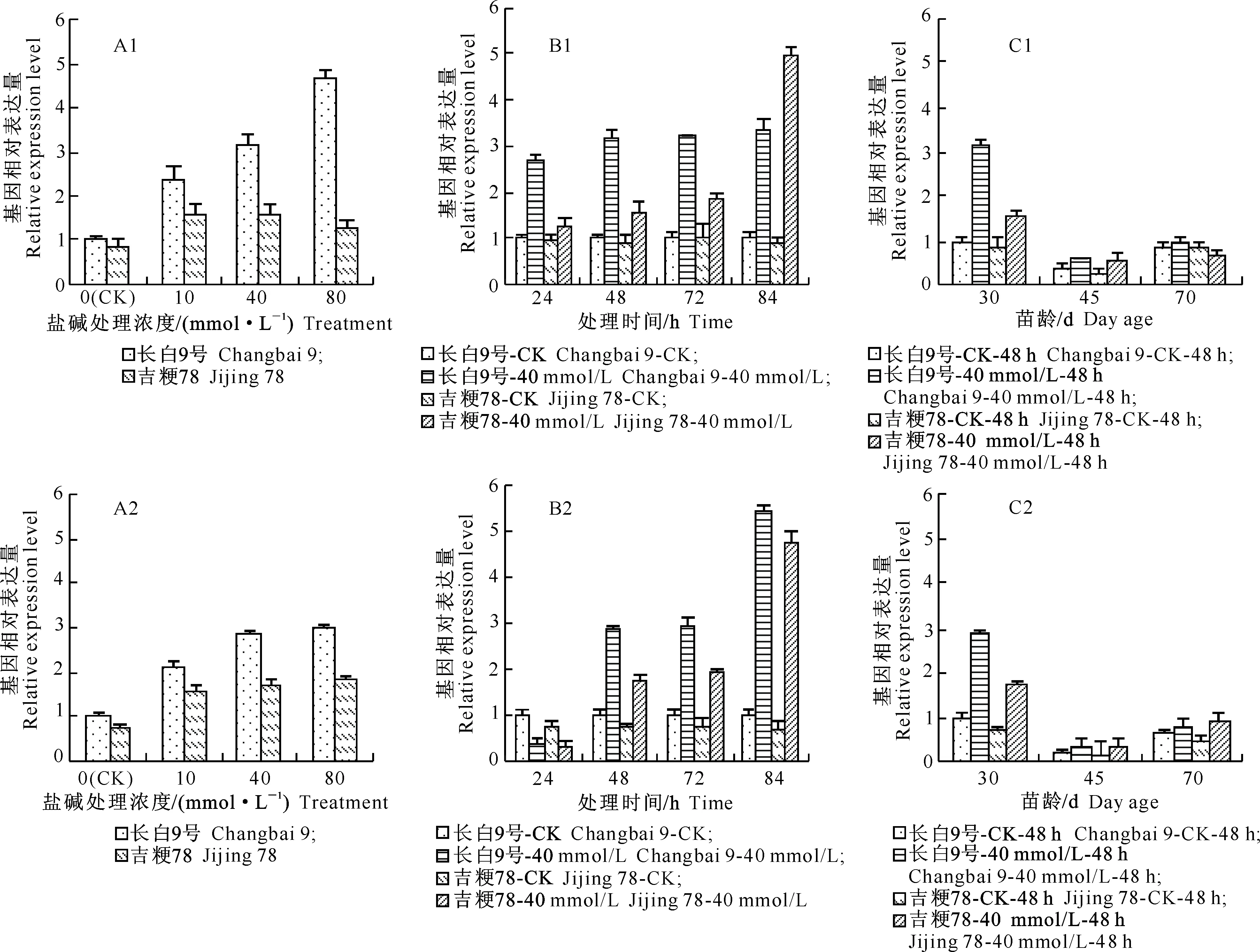
图 2 盐碱胁迫对不同品种水稻叶片BADH1基因(A1~C1)和BADH2基因(A2~C2)表达量的影响

图 3盐碱胁迫对长白9号和吉粳78水稻叶片IMP基因表达量的影响
Fig.3Effect of saline-alkali stress on expressions ofIMPgene in leaves of Changbai 9 and Jijing 78
由图3-C可知,2水稻品种CK中IMP基因表达量都随苗龄的增加而呈明显的下降趋势, 苗龄30 d处理样品IMP基因表达量与CK相比变化最大。
3讨论
通过积累渗透调节物质植物可以抵抗盐碱胁迫的伤害[20-21]。杨涓等[3]研究显示,在高盐环境下植物细胞中常积累一些小分子有机物(如脯氨酸、甜菜碱、糖醇等)维持高的细胞质渗透压,以确保细胞的正常生理功能。张乐新等[22]和Reda等[23]报道,过表达P5CS和BADH基因能够提高植物脯氨酸和甜菜碱含量,且植株的抗逆性提高。本研究结果显示,在未经盐碱胁迫时,耐盐碱品种长白9号叶片中的脯氨酸、甜菜碱含量高于普通品种吉粳78;盐碱胁迫能够提高2种水稻叶片中脯氨酸、甜菜碱和肌醇的含量,盐碱胁迫24和48 h就能使长白9号及时做出响应,并随着胁迫时间的延长甜菜碱、肌醇含量持续提高,而吉粳78则相对晚一些才能做出响应。同时,2种水稻30 d苗在不同盐碱浓度胁迫下P5CS、BADH1、BADH2、IMP4个基因的表达量也均上升,而且各基因表达量与其渗透调节产物具有相似的变化规律。品种间相比较,长白9号基因表达量上调比吉粳78明显。
许多报道表明,脯氨酸和甜菜碱是较为重要和有效的有机渗透调节物质[24-25],而肌醇[8]发现较晚且作用也没有脯氨酸和甜菜碱明显。本试验显示,盐碱胁迫导致长白9号与吉粳78各龄苗叶片中脯氨酸、甜菜碱、肌醇含量均上升,各处理脯氨酸和甜菜碱含量及变化均明显高于肌醇。2品种30 d龄苗P5CS、BADH1、BADH2 3个基因在不同盐碱浓度、不同胁迫时间下表达量均上升,而IMP基因表达量只在长白9号中明显上升,在吉粳78中没有明显变化。本试验利用10,40和80 mmol/L Na2CO3-NaHCO3对长白9号和吉粳78水稻品种进行处理,结果表明各盐碱处理样品脯氨酸、甜菜碱含量均高于CK。张丽丽等[15]研究表明,10,20,30 mmol/L Na2CO3-NaHCO3对长白9号处理之后,叶片脯氨酸含量随浓度的增大而增大。而本试验发现,40 mmol/L Na2CO3-NaHCO3处理较10和80 mmol/L处理更有利于植株积累渗透调节物质及上调基因表达。
宋冬明等[26]研究表明,水稻幼苗期对盐碱处理较敏感,随着植株生育进程的推进,对盐碱的耐性增加。从脯氨酸、甜菜碱和肌醇3种渗透调节物质的含量及4个与渗透胁迫相关基因的表达量来看,本试验中耐盐碱品种长白9号和普通品种吉粳78都表现为30 和45 d龄苗变化均比70 d龄苗明显,而45 d龄苗与30 d龄苗相比较,盐碱胁迫样品与CK间渗透调节物质的含量变化较小,说明45 d龄苗对盐碱处理表现更为稳定,对盐碱耐受性较好。
[参考文献]
[1]徐鹏程,冷翔鹏,刘更森,等.盐碱土改良利用研究进展 [J].江苏农业科学,2014,42(5):293-298.
Xu P C,Leng X P,Liu G S,et al.Review of improvement and utilization of saline-alkali soil [J].Jiangsu Agricultural Sciences,2014,42(5):293-298.(in Chinese)
[2]董芸雷,妥德宝,李彬,等.脱硫石膏和有机肥不同施用量改良河套灌区盐碱化土壤田间试验研究 [J].海南师范大学学报,2013,26(3):299-302.
Dong Y L,Tuo D B,Li B,et al.Field experiment study of the improvement of saline-alkali soil in hetao irrigation area by different dosage of desulfurization gypsum and organic fertilizer [J].Journal of Hainan Normal University,2013, 26(3):299-302.(in Chinese)
[3]杨涓,许兴.盐胁迫下植物有机渗透调节物质积累的研究进展 [J].宁夏农学院学报,2003(4):86-91.
Yang J,Xu X.Progress of research on accumulation of osmotic substance under salt-stress in plants [J].Journal of Ningxia Agricultural College,2003(4):86-91.(in Chinese)
[4]Verbruggen N,Hermans C.Proline accumulation in plants:A re-view [J].Amino Acids,2008,35(4):753-759.
[5]Timothy A G.Assigning enzymatic function to betain-homocys-teine S-methyltransfearse-2(BHMT2) as an S-methylmethionine(SMM)-specific homocysteine(Hcy) methyl transferase [J].FASEB J,2006,20:A606.
[6]曾幼玲,幸婷,蔡忠贞,等.盐生植物盐爪爪甜菜碱醛脱氢酶基因的克隆及在盐胁迫下的BADH基因的表达 [J].云南植物研究,2007,29(1):79-84.
Zeng Y L,Xing T,Cai Z Z,et al.Molecular cloning and expression analysis of betain aldehyde dehydrogenase gene from the halophyteKalidiumfoliatumin Xinjiang on salinity [J].Acta Botanica Yunnanica,2007,29(1):79-84.(in Chinese)
[7]Makoto S,Kunihiko T,Mio K,et al.Expression pattern of inositol phosphate-related enzymes in rice:Implications for the phytic acid biosynthetic pathway [J].Gene,2007,405(4):55-64.
[8]王艳青,陈雪梅,李悦,等.植物抗逆中的渗透调节物质及其转基因工程发展 [J].北京林业大学学报,2001,23(4):66-70.
Wang Y Q,Chen X M,Li Y,et al.The osmotic solute in plant resisistance to adverse conditions and progress in relative genic engineering [J].Journal of Beijing Forestry University,2001,23(4):66-70.(in Chinese)
[9]严海霞,何文寿.宁夏银北地区盐碱地改良与水稻种植技术探讨 [J].湖北农业科学,2010,49(11):2963-2965.
Yan H X,He W S.Improvement of saline-alkali soil and rice cultivation technology in northern Yinchuan of Ningxia [J].Hubei Agricaltural Sciences,2010,49(11):2963-2965.(in Chinese)
[10]王云贺,王志春,杨帆,等.不同改良物质对苏打碱土盐碱度及水稻生长的影响 [J].华南农业大学学报,2013,34(4):445-449.
Wang Y H,Wang Z C,Yang F,et al.Effects of different ameliorate materials on soda alkaline soil salinity-alkalinity and rice growth [J].Journal of South China Agricultural University,2013,34(4):445-449.(in Chinese)
[11]孙宇男,耿玉辉,赵兰坡.硫酸铝改良苏打盐碱土后各离子的变化 [J].中国农学通报,2011,27(23):255-258.
Sun Y N,Geng Y H,Zhao L P.The change of adding aluminum sulfate on saline alkali soil,sion [J].Chinese Agricultural Science Bulletin,2011,27(23):255-258.(in Chinese)
[12]赵国臣,齐春艳,侯立刚,等.吉林省苏打盐碱地水稻生产历史进程与展望 [J].沈阳农业大学学报,2012,43(6):673-680.
Zhao G C,Qi C Y,Hou L G,et al.Progress and prospect of rice production in Jilin province at soda saline-alkaline land [J].Journal of Shenyang Agicultural University,2012,43(6):673-680.(in Chinese)
[13]李彻.吉林省水稻育种取得突破长白9号种植面积超过外引品种 [J].中国稻米,1995(6):22.
Li C.Breakthroughts in rice breeding in Jilin province Changbai 9 planting area more than irfpa varieties [J].China Rice,1995(6):22.(in Chinese)
[14]杨春武.虎尾草和水稻抗碱机制研究 [D].上海:东北师范大学,2010.
Yang C W.Mechanisms of alkali tolerance inChlorisvirgataand rice [D].Shanghai:Northeast Normal University,2010.(in Chinese)
[15]张丽丽,马殿荣,陈温福,等.杂草稻幼苗对Na2CO3+NaHCO3胁迫响应及其生理基础 [J].华北农学报,2011,26(6):102-107.
Zhang L L,Ma D R,Chen W F,et al.Responses of Na2CO3+NaHCO3stress on seedling of tolerant weedy rice and physiological basis [J].Acta Agriculturae Booreali-Sinica, 2011,26(6):102-107.(in Chinese)
[16]艾琳,张萍,胡成志.低温胁迫对葡萄根系膜系统和可溶性糖及脯氨酸含量的影响 [J].新疆农业大学学报,2004,27(4):47-50.
Ai L,Zhang P,Hu C Z.Effects of low temperatures tolerance on the electrolyte osmotic rate and the content of soluble sugar proline of grape roots [J].Journal of Xinjiang Agricultural University,2004,27(4):47-50.(in Chinese)
[17]刘志刚,钱捷,杨明康.甜菜碱含量测定方法研究进展 [J].江西化工,2012,3(3):9-11.
Liu Z G,Qian J,Yang M K.Research progress on content determination of betaine [J].Jiangxi Chemical Industry,2012,3(3):9-11.(in Chinese)
[18]王秀丽,李叶云,周会.败酱草中肌醇含量测定 [J].安徽中医学院学报,2002,21(1):51-54.
Wang X L,Li Y Y,Zhou H.Research on content determination of inositol in patrina [J].Journal of Anhui University of Chinese Medicine,2002,21(1):51-54.(in Chinese)
[19]韦克苏,张其芳,程方民,等.花后高温下水稻可溶性淀粉合酶同工型基因的表达模式 [J].作物学报,2009,35(1):18-24.
Wei K S,Zhang Q F,Cheng F M,et al.Expression profiles of rice soluble starch synthase gene in response to high temperature stress at filling stage [J].Acta Agronomica Sinica,2009,35(1):18-24.(in Chinese)
[20]Munns R,Tester M.Mechanisms of salinity tolerance [J].Plant Biol,2008,59:651-681.
[21]王宁,曹敏健,于佳林.NaCl胁迫对玉米幼苗有机渗透调节物质的影响 [J].玉米科学,2009,17(4):61-65.
Wang N,Cao M J,Yu J L.Effects of NaCl stress on organic osmotica of maize seedlings [J].Journal of Maize Sciences,2009,17(4):61-65.(in Chinese)
[22]张乐新,苏蔓,马甜.羊草Δ1-吡咯琳-5-羧酸合成酶基因的克隆与分析 [J].草业学报,2013,22(4):197-204.
Zhang L X,Su M,Ma T.Cloning and analysis of the Δ1-pyrroline-5-carboxylate synthetase fromLeymuschinensis[J].Acta Prataculturas Sinica,2013,22(4):197-204.(in Chinese)
[23]Reda E A M,Hirofumi S,Kounosuke F.Effect of salinity on osmotic adjustment,glycinebetaine accumulation and the betaine aldehyde dehydrogenase gene expression in two halophytic plants,SalicorniaeuropaeaandSuaedemaritime[J].Plant Science,2004,166:1345-1349.
[24]Wang Z Q,Yuan Y Z,Qu J Q,et al.Glutamine synthetase and glutamate dehydrogenase contribute differentially to proline accumulation in leaves of wheat seedlings exposed to different salinity [J].Plant Physiol,2007(164):695-701.
[25]Timothy L F,Daniel L E W,Robert J H.The effect of salt on betaine aldehyde dehydrogenase transcript levels and 2-acetyl-1-pyrroline concentration in fragrant and non-fragrant rice (Oryzasativa) [J].Plant Science,2008(175):539-546.
[26]宋冬明,贺梅,李春光.水稻耐盐研究进展及展望 [J].北方水稻,2012,43(1):74-77.
Song D M,He M,Li C G.Research progress and expectation of rice salt tolerant [J].North Rice,2012,43(1):74-77.(in Chinese)
Changes in osmolytes and related genes of different rice varieties under saline-alkali stress
ZHANG Ting-ting1a,YANG Mei-ying1a,WANG Chun-hong1a,SUN He-mei1a,QI Chun-yan2,HOU Li-gang2,WU Zhi-hai1b
(1 aCollegeofLifeScience,bCollegeofAgronomy,JilinAgriculturalUniversity,Changchun,Jilin130118,China;2InstituteofRiceResearchJilinAcademyofAgriculturalSciences,Gongzhuling,Jilin136102,China)
Abstract:【Objective】 This study analyzed the changes in osmolytes and genes expressions of 2 rice varieties under different saline-alkali concentrations and stress times to provide basis for species selection and plant protection.【Method】 Rice varieties Changbai 9 (saline-alkali-tolerant) and Jijing 78 (normal) were selected to study the changes in contents of proline,betaine and inositol and expressions of P5CS,BADH1,BADH2,and IMP in leaves of seedlings at 3 different ages (30,45,70 d) under 3 NaCO3-NaHCO3 concentrations (10,40,80 mmol/L) and 4 saline-alkali stress times (24,48,72,84 h).【Result】 The contents of proline and betaine in leaves of Changbai 9 were higher than that of Jijing 78 under normal hydroponic condition.With saline-alkali stress,the contents of proline,betaine and inositol in leaves of both Changbai 9 and Jijing 78 increased and the expressions of P5CS,BADH1,BADH2,and IMP were also higher than CK.The treatment with 40 mmol/L NaCO3-NaHCO3 was more conducive to accumulate osmolytes and regulate gene expression than treatments with 10 and 80 mmol/L NaCO3-NaHCO3.However,the changes in osmolytes contents and genes expressions in Changbai 9 leaves were more significant than that of Jijing 78.Changbai 9 responded in time after 24 and 48 hours stress and the contents of betaine and inositol as well as expression levels of BADH1,BADH2,and IMP increased continuously with the extension of stress time.Jijing 78 behaved differently and slowly.The seedlings at the ages of 30 and 45 days had more significant responses than that at the age of 70 days.【Conclusion】 Changbai 9 could tolerate high saline-alkali stress because it is rich in osmolytes and responded quickly to stress.The sensitive thresholds of both rice varieties were consistent,and saline-alkali-tolerant variety was more suitable for large seedling transplanting within thresholds.
Key words:rice;saline-alkali stress;osmolytes;proline;betaine;inositol
DOI:网络出版时间:2016-03-1408:4510.13207/j.cnki.jnwafu.2016.04.006
[收稿日期]2014-08-15
[基金项目]吉林省自然科学基金项目(201215183);吉林省教育厅重点项目(2012044);吉林省博士后科研启动基金项目
[作者简介]张婷婷(1988-),女,辽宁抚顺人,硕士,主要从事水稻生物化学与分子生物学研究。[通信作者]武志海(1975-),男,内蒙古呼和浩特人,副教授,主要从事水稻栽培生理研究。E-mail:wuzhihai1116@163.com
[中图分类号]S511.01
[文献标志码]A
[文章编号]1671-9387(2016)04-0039-09
网络出版地址:http://www.cnki.net/kcms/detail/61.1390.S.20160314.0845.012.html
E-mail:zhangtingting0526@126.com
