高体积分数CO2处理对柿果实褐变的影响
2016-05-03闵婷郑梦林李阳殷学仁陈昆松易阳
闵婷+郑梦林+李阳+殷学仁+陈昆松+易阳+王宏勋

摘要: 涩柿果实采收后须经人工脱涩才能用于鲜食,其中高体积分数CO2脱涩处理应用最广泛。经高体积分数CO2处理后,柿果实在贮藏过程中也会发生褐变。多酚氧化酶PPO作为酶促褐变最关键的基因,在各种果蔬中的研究较多,而关于PPO基因是否参与涩柿果实脱涩后褐变的研究尚未见报道。以“恭城水柿”为材料,经95% CO2处理2 d后切片,通过可溶性单宁含量测定、褐变程度观察、PPO基因克隆与表达分析研究CO2处理对“恭城水柿”褐变的影响。结果表明,经95% CO2处理后,柿果实的可溶性单宁含量明显下降,柿果实完成脱涩。脱涩后的柿鲜切片于20 ℃下放置12 h后发生明显褐变。本研究克隆获得PPO基因部分片段序列,聚类分析表明,该PPO基因与苹果PPO基因较同源;基因表达分析显示,该PPO基因在CO2处理后的鲜切柿片中的表达量比对照低,表明该PPO基因可能与柿子脱涩后的褐变无相关性。
关键词: 恭城水柿;脱涩;褐变;PPO
中图分类号: TS255.3 文献标志码: A 文章编号:1002-1302(2016)03-0288-03
柿(Diospyros kaki)属于柿科(Ebenaceae)柿属(Diospyros),起源于东亚,主要分布于中国、日本、韩国等东亚国家[1]。不同于其他水果,柿果实的独特之处在于其能累积大量单宁,其中可溶性单宁可导致涩味。柿果实根据其自然脱涩的能力分为甜柿和涩柿。甜柿能在树上自然完成脱涩,采收后可直接食用;涩柿果实成熟时可溶性单宁含量仍较高,采收后须经人工脱涩方能鲜食[2]。我国栽培的品种多为涩柿,涩柿的脱涩技术较为成熟,包括乙烯处理、CO2处理[3]、温水浸泡、交替冻融[4]等,其中高体积分数CO2处理应用最为广泛。然而,高体积分数CO2处理柿果实后,果实在贮藏过程中也会发生褐变,且处理时间越长则褐变程度越深[5],使柿果实在贮藏和货架期的果实品质下降。
褐变是果实品质劣变最明显的特征之一,不仅影响果品的营养、外观、风味,并已成为果蔬贮藏加工的主要障碍[6]。果蔬褐变多以酶促褐变为主,与酶促褐变密切相关的酶类主要为多酚氧化酶(polyphenol oxidase,PPO),多酚类物质在PPO的作用下氧化成醌,醌再聚合生成黑色或褐色的色素沉淀,组织表现为褐变。随着分子生物学的发展,马铃薯[7]、梨[8]、甘薯[9]、蘑菇[10]、莲藕[11]等植物的PPO基因相继被克隆,而关于柿果实中PPO酶活性及其特性的报道较少。
以“恭城水柿”为材料,经95% CO2处理2 d后切片,通过可溶性单宁含量分析、褐变程度观察、PPO基因克隆与表达分析研究CO2处理对“恭城水柿”褐变的影响,并进行PPO基因的克隆、进化树分析、基因表达分析,以期阐明柿果实脱涩后褐变的生物学机制。
1 材料与方法
1.1 原料与试剂
供试“恭城水柿”购自广西壮族自治区恭城县商业果园。甲醇、没食子酸、福林酚、NaCO3、NaCl、HCl、十六烷基三甲基溴化铵、三羟甲基氨基甲烷、乙二胺四乙酸、聚乙烯吡咯烷酮、LiCl、十二烷基硫酸钠、氯仿、异戊醇、无水乙醇均购自国药集团化学试剂有限公司。DEPC水、亚精胺、巯基乙醇均购自上海生工试剂公司。液氮购自湖北省武汉市明辉气体有限公司。双蒸水。
1.2 主要仪器
V-110D型紫外可见分光光度计(上海美谱达仪器有限公司),HH-S4型数显恒温水浴锅(金坛市医疗仪器厂),GL-20G-2型飞鸽牌离心机(上海安亭科学仪器厂),00101D型立式压力蒸汽灭菌锅(上海博讯实业有限公司医疗设备厂),S2-93型自动双重纯水蒸馏器(上海亚荣生化仪器厂),DHG-9140A型电热鼓风干燥器(上海一恒科学仪器有限公司),HYC-326A型医用冷藏箱(青岛海尔特种电器有限公司),DYCP-31DN型电泳仪(北京市六一仪器厂),CP214(C)型电子天平(上海奥豪斯仪器有限公司),PTT-A500型电子天平(福州华志科学仪器有限公司),IMS-20型雪科制冰机(常熟市雪科电器有限公司),凝胶成像仪(美国伯乐公司)。
1.3 试验方法
1.3.1 脱涩处理 成熟“恭城水柿”果实自商业果园采收后,于2 d内运至实验室。选择成熟度相对一致、果形匀称、大小中等、无机械损伤的果实进行相关采后试验。将果实平均分为2组,每组约50个,一组经95% CO2处理2 d后转货架[12];另一组作为对照,置于20 ℃下直至试验结束。
1.3.2 切片处理 将脱涩试验完成后的处理组和对照组果实去皮,并切片为3.0 cm×3.0 cm×0.3 cm大小,2组各切30片,置于20 ℃下12 h后拍照,液氮取样用于后期试验。
1.3.3 可溶性单宁含量测定 采用福林酚方法[12]进行可溶性单宁含量测定。没食子酸用于标准曲线的制作,每次测量重复3次,每个点进行3次生物学重复。可溶性单宁的质量分数用果实质量的百分含量表示。
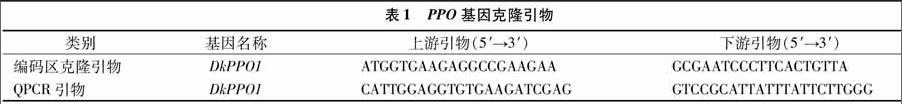
1.3.4 PPO基因的克隆与表达 进行PPO基因的克隆[11],根据前期RNA-seq获得PPO基因序列,利用Primer 3在线软件(http://bioinfo.ut.ee/primer3-0.4.0/primer3/)设计引物(表1)。按照试剂盒PCR扩增所得的目标片段连接,转化到大肠杆菌DH5α(TaKaRa公司),挑选阳性克隆并进行测序验证。
进行PPO基因表达试验[12],包括模板合成、引物特异性检测、实时定量PCR(QPCR)分析。
2 结果与分析
2.1 95% CO2处理对柿果实可溶性单宁含量的影响
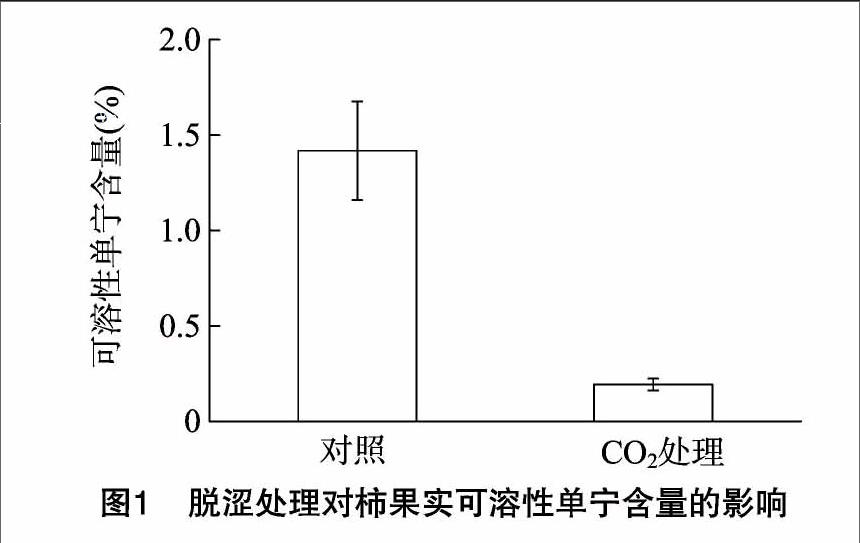
可溶性单宁是柿果实涩味的主要呈现者,可溶性单宁含量越高则涩味越严重。试验测定了对照组及95% CO2处理2 d后的柿果实可溶性单宁含量。结果表明,经95% CO2处理后,可溶性单宁含量仅为0.19%,明显低于对照组的1.40%(图1),这与已有研究的结论[12]相一致。可见,95% CO2处理能明显降低“恭城水柿”的可溶性单宁含量,实现柿果实的脱涩。
2.2 95% CO2处理对柿果实褐变的影响
将95% CO2处理后的柿果实切片为3.0 cm×3.0 cm×0.3 cm,于室温放置12 h后观察拍照(图2)。结果表明,对照果实无明显变化;经95% CO2处理后的果实切片褐变现象明显,这与已有研究的结论[5]相一致。可见,95% CO2处理能使“恭城水柿”在贮藏后期发生褐变。
2.3 柿果实PPO基因的克隆与表达
根据前期RNA-seq结果设计PPO基因克隆引物、荧光定量PCR引物(表1),并克隆获得PPO基因部分片段序列,命名为DkPPO。与马铃薯、苹果、番茄的PPO基因进行聚类分析显示,DkPPO与苹果PPO基因同源关系较近(图3)。进一步基因表达分析表明,该PPO基因在CO2处理后的鲜切柿片中的表达量比对照低(图4),表明该PPO基因可能未参与柿果实脱涩处理后的褐变。
3 结论
本研究探讨了CO2处理对“恭城水柿”褐变的影响。结果表明,柿果实经95% CO2处理后,其可溶性单宁含量明显下降,柿果实完成脱涩。脱涩后的柿鲜切片于20 ℃下放置12 h后发生明显褐变。克隆获得PPO基因部分片段序列,基因表达结果显示,DkPPO基因在CO2处理后的鲜切柿片中的表达量比对照低,表明该PPO基因可能未参与柿果实脱涩处理后的褐变。CO2处理导致“恭城水柿”褐变的原因有待进一步研究。
参考文献:
[1]Wang R Z,Yang Y,Li G C. Chinese persimmon germplasm resources[J]. Acta Horticulturae,1997,436:43-50.
[2]Min T,Yin X R,Shi Y N,et al. Ethylene-responsive transcription factors interact with promoters of ADH and PDC involved in persimmon(Diospyros kaki)fruit de-astringency[J]. Journal of Experimental Botany,2012,63(18):6393-6405.
[3]Taira S,Ono M,Otsuki M. Effects of freezing rate on astringency reduction in persimmon during and after thawing[J]. Postharvest Biology and Technology,1998,14(3):317-324.
[4]Akagi T,Ikegami A,Tsujimoto T,et al. DkMyb4 is a Myb transcription factor involved in proanthocyanidin biosynthesis in persimmon fruit[J]. Plant Physiology,2009,151(4):2028-2045.
[5]Novillo P,Salvador A,Llorca E,et al. Effect of CO2 deastringency treatment on flesh disorders induced by mechanical damage in persimmon:Biochemical and microstructural studies[J]. Food Chemistry,2014,145:454-463.
[6]宋晓雪,胡文忠,毕 阳,等. 鲜切果蔬酶促褐变关键酶的研究进展[J]. 食品工业科技,2013,34(15):390-393.
[7]Hunt M D,Eannetta N T,Yu H F,et al. cDNA cloning and expression of potato polyphenol oxidase[J]. Plant Molecular Biology,1993,21(1):59-68.
[8]Nishimum M F C,Some Properties of Japanese pear(Pyrus pyrifolia) po1yphenol oxldase and changes in browning potential during fruit maturation[J]. Journal of the Science of Food and Agriculture,2003,83:1156-1162.
[9]Liao Z,Chen R M. Molecular cloning and characterization polyphenol oxidase gene from sweet potato[J]. Molecular Biology,2006,40(6):907-913.
[10]Li N Y,Wm C,Jin Q L,et al. Molecular cloning and expression of polyphenol oxidase gene from the mushroom,Agaricus bisporus[J]. Agricultural Science in China,2011,10(2):185-194.
[11]张跃进,郝晓燕,梁宗锁,等. 莲藕多酚氧化酶基因(PPO)的克隆与表达分析[J]. 农业生物技术学报,2011,19(4):634-641.
[12]Min T,Fang F,Ge H,et al. Two novel anoxia-induced ethylene response factors that interact with promoters of deastringency-related genes from persimmon[J]. PLoS One,2014(9):e97043.
