一种新型醛酮还原酶的克隆表达、性质研究以及在不对称合成 (R)-CHBE中的应用
2016-04-26张瑾成
张瑾成,魏 淼,许 琳,严 明
(南京工业大学生物与制药工程学院,江苏南京211800)
一种新型醛酮还原酶的克隆表达、性质研究以及在不对称合成 (R)-CHBE中的应用
张瑾成,魏淼,许琳,严明
(南京工业大学生物与制药工程学院,江苏南京211800)
摘要:为开发催化4-氯乙酰乙酸乙酯(COBE)制备(R)-4-氯-3-羟基丁酸乙酯((R)-CHBE)的新型催化剂,挖掘到了来自白色念珠菌SC5314中的一种NADPH辅酶依赖型醛酮还原酶CAK基因(cak),并将该基因在大肠杆菌中表达。将重组酶进行纯化后,测定其酶学性质,并构建了以葡萄糖为辅底物的双酶偶联辅酶再生系统,考察其不对称转化制备(R)-CHBE的能力。结果表明:CAK对多种醛酮类化合物有催化活性,其催化COBE的最适反应温度为40 ℃,最适pH为5。CAK在40 ℃下以及酸性条件中能保持较好的稳定性。Mg(2+)、Na+、 K+对酶活有一定的激活作用,而Cu(2+)存在条件下酶会彻底失活。乙酸乙酯、邻苯二甲酸二丁酯对酶活的抑制作用较小。利用双酶偶联辅酶再生系统不对称转化制备(R)-CHBE。在合适的条件下,转化600 mmol/L的底物,产率达80.6%,产物对映体过量值(e.e.值)>99%。
关键词:醛酮还原酶;NADPH再生;蛋白纯化;酶学性质;(R)-4-氯-3羟基丁酸乙酯
(R)-4-氯-3-羟基丁酸乙酯((R)-CHBE)作为一种手性醇,是合成L-肉碱[1]、阿托伐他汀钙[2]、(-)-大内酰亚胺A[3]、(R)-γ-氨基-β-羟基丁酸(GABOB)[4]、负霉素[5]、(R)-4-氨基-3-羟丁酸[6]等手性药物的重要中间体,全细胞催化还原4-氯乙酰乙酸乙酯(COBE)制备手性CHBE具有反应条件温和、操作工艺相对简单、对环境污染小等优势,是目前首选的制备方法。早期的研究多采用野生细胞作为催化剂[7-8],其缺点是产率及光学纯度很低,且筛选野生型菌株非常困难。为了克服这些缺点,近年来的研究侧重于将能够不对称还原COBE的酶在工程菌中过表达用于催化。目前,文献报道的用于不对称还原COBE制备(R)-CHBE的酶以醛酮还原酶[9](AKR)、短链脱氢酶[10](SDR)、羰基还原酶[11](CAR)为主,然而其中大多数酶在制备(R)-CHBE中仍然不能同时获得较高的产率和光学纯度。He等[12]将一种来自于鲑色锁掷酵母的羰基还原酶SsCR用于不对称还原制备(R)-CHBE,其产率和产品的光学纯度均达到较高的水平(转化600 mmol/L COBE,产率为100%,光学纯度为大于99%),但反应需在有机-水两相并添加辅酶的情况下进行。
酶催化合成手性醇时需要辅酶NAD(P)H提供氢,额外添加会大大提高生产成本。引入另一种酶及辅底物构建双酶偶联反应,使NAD(P)+再生为NADPH,可以实现辅酶的循环,避免辅酶的持续添加。例如使用葡萄糖脱氢酶和葡萄糖就可以将NADP+再生为NADPH[13]。在全细胞催化中,为实现双酶耦联反应,可以将用于不对称还原COBE的酶与葡萄糖脱氢酶在大肠杆菌中共表达,相比于将两种酶单独表达,这种方式的效率通常会更高。
本研究利用基因数据挖掘的手段,发现一种白色念珠菌来源的醛酮还原酶CAK,将其在大肠杆菌中表达,研究其酶学性质。同时构建双酶偶联辅酶再生系统,将其应用于不对称转化制备(R)-CHBE中,并对多个催化条件进行优化,以期实现经济高效催化制备(R)-CHBE的目的。
1材料和方法
1.1材料
1.1.1菌株和试剂
E.coliDH5α、E.coliRosetta、质粒pET-22b(+)及白色念珠菌(CandidaalbicansSC5314)基因组,保藏于笔者所在实验室。
4-氯乙酰乙酸乙酯,Fluka公司;(R)/(S)-4-氯-3羟基丁酸乙酯标准品,Sigma-Aldrich公司;限制性内切酶、T4 DNA连接酶、Prime STAR HS DNA 聚合酶,宝生物工程(大连)有限公司;蛋白质marker,Fermentas公司;质粒抽提试剂盒及DNA凝胶回收试剂盒,天根生化科技(北京)有限公司;抗生素及其余试剂,生工生物工程(上海)股份有限公司。
1.1.2培养基
种子培养基使用LB培养基[14]。
发酵培养基(g/L):胰蛋白胨25、酵母提取物15、 NaCl 10、乳糖3、 葡萄糖2。用于诱导目标蛋白的表达。
1.2方法
1.2.1醛酮还原酶CAK的克隆
根据来自白色念珠菌CandidaalbicansSC5314醛酮还原酶cDNA序列(GenBank:XM_714761)设计引物。上游、下游引物分别引入NdeI及NcoI酶切位点。上游引物为5′- G ̄G ̄A ̄A ̄T ̄T ̄C ̄C ̄A ̄T ̄A ̄T ̄G ̄C ̄C ̄A ̄G ̄C ̄T ̄C ̄A ̄A ̄T ̄T ̄G ̄C ̄A-3′,下游引物为5′-C ̄A ̄T ̄G ̄C ̄C ̄A ̄T ̄G ̄G ̄T ̄T ̄A ̄A ̄T ̄C ̄A ̄T ̄C ̄A ̄A ̄A ̄G ̄T ̄T ̄G ̄T ̄T ̄G ̄A-3′(划线处为酶切位点)以白色念珠菌基因组为模板,通过PCR获得目的片段cak。cak片段和质粒pET-22b(+)经过NdeI和NcoI 37 ℃双酶切、连接后转化大肠杆菌DH5α。在含有氨苄青霉素(100 μg/mL)和氯霉素(34 μg/mL)的抗性平板上挑取阳性克隆后抽提质粒,送交南京金斯瑞生物科技有限公司测序。
1.2.2醛酮还原酶CAK的表达
经测序验证后,将重组质粒转化表达宿主E.coliRosetta。接种于含氨苄青霉素(100 μg/mL )和氯霉素(34 μg/mL)的液体 LB 培养基中,于 37 ℃培养,8 h后,将菌液按2%(体积分数)接种量转接于 50 mL(500 mL三角烧瓶)含氨苄青霉素(100 μg/mL)和氯霉素(34 μg/mL)的发酵培养基中,37 ℃、220 r/min振荡培养3 h后,转 30 ℃、220 r/min 振荡培养12 h。培养结束后,8 000 r/min离心20 min收集菌体。菌体用同体积 pH 7.0的Na3PO4缓冲悬浮后超声破碎,8 000 r/min低温离心20 min,收集到的上清液即为粗酶液。
1.2.3蛋白浓度测定及SDS-PAGE分析
蛋白浓度测定采用Bradford法[15]。SDS-PAGE采用分子克隆实验指南[16]提供的方法,分离胶质量分数为12%,浓缩胶质量分数为5%。粗酶液加入2× Loading Buffer后,95 ℃水浴5 min后上样,上样量为10 μL。
1.2.4酶活力测定
酶活力测定体系包括1 mmol/L NADPH、10 mmol/L底物COBE和pH 7.0 Na3PO4缓冲液,加入适当粗酶液启动反应。于30 ℃下测定波长340 nm处吸光值的下降率。酶活定义:每分钟内氧化1 μmol NAD(P)H所需要的酶量为一个酶活单位U。其计算见式(1)。
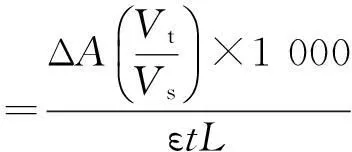
(1)
式中:ΔA为340 nm处吸光值的变化值;ε为消光系数,取值为6 220;Vt为总反应体积(mL);Vs为酶液体积(mL);L为光径,取值为1 cm;t为反应时间(min)。
1.2.5蛋白纯化
将粗酶液进行(NH4)2SO4沉淀以除去部分杂蛋白,再使用疏水层析柱进行进一步纯化。层析柱首先用pH 7.4、含有1.5 mol/L (NH4)2SO4的20 mmol/L磷酸盐缓冲液(PBS)预平衡。洗脱缓冲液为pH 7.4、20 mmol/L的磷酸盐,采用0~100%缓冲液梯度洗脱。收集到的活性蛋白经透析、离心处理后,最后得到的上清即为纯化后的酶液。
1.2.6酶的底物谱测定
醛酮还原酶CAK的底物谱测定选用的底物主要包括醛类和酮类化合物:乙醛、戊二醛、正丁醛、正戊醛、辛醛、苯甲醛、甘油醛、丙酮醛、柠檬醛、乙酰乙酸乙酯、COBE、丙酮、丁酮、3-戊酮、2,3-戊二酮、2-辛酮、乙酰丙酮、甲基异丁基甲酮。分别将上述物质以10 mmol/L的浓度加入酶活测定体系测定酶活。
1.2.7温度对酶活及稳定性的影响
将纯化后的酶液分别在不同温度的反应条件下测定其酶活力,研究温度对酶活的影响。将酶液置于20、25、30、35、40、45和50 ℃的水浴中,每隔10 min取样,分别测定残余酶活以研究酶的热稳定性。
1.2.8pH对酶活的影响及酶的pH稳定性
使用不同pH的缓冲体系测定酶活,研究pH对酶活的影响,缓冲体系为pH 4.0~5.5醋酸盐缓冲液、pH=5.5~7.5磷酸盐缓冲液、pH 7.5~9.0 Tris缓冲液。将酶液分别置于pH=4.0~9.0的缓冲液中,4 ℃保存24 h后测定酶活力,研究酶的pH稳定性。
1.2.9金属离子对酶活的影响
向纯化后的酶液中分别加入1 mmol/L的Mg2+、Mn2+、Zn2+、Ca2+、Fe3+、Cu2+、Ni2+、Co2+、Na+和K+,在4 ℃下孵育1 h后测定酶活,考察金属离子对酶活的影响。
1.2.10有机溶剂对酶活的影响
向酶活测定体系中加入10%~50%(体积分数)的有机溶剂(乙酸乙酯、乙酸丁酯、辛酸乙酯、三氯甲烷、邻苯二甲酸二丁酯、异丙醚),分别测定酶活,考察金属离子对酶活的影响。
1.2.11双酶共表达菌株的构建及表达
将葡萄糖脱氢酶gdh基因片段插入至重组质粒pET-cak中cak片段下游实现双酶共表达,具体方法:设计引物扩增gdh基因(来自巨大芽胞杆菌)并在gdh片段5′端引入SD-AS序列。上游引物为C ̄A ̄T ̄G ̄C ̄C ̄A ̄T ̄G ̄G ̄T ̄A ̄A ̄G ̄G ̄A ̄G ̄G ̄A ̄T ̄A ̄T ̄A ̄C ̄A ̄T ̄A ̄T ̄G ̄T ̄A ̄T ̄A ̄C ̄A ̄G ̄A ̄T ̄T ̄T ̄A ̄A ̄A ̄A ̄G(含NcoI酶切位点),下游引物为A ̄T ̄A ̄A ̄G ̄A ̄A ̄T ̄G ̄C ̄G ̄G ̄C ̄C ̄G ̄C ̄T ̄T ̄A ̄C ̄T ̄A ̄G ̄C ̄C ̄T ̄C ̄T ̄T ̄C ̄C ̄T ̄G ̄C(含NotI酶切位点)经PCR扩增后,将重组质粒pET-cak和片段分别用NcoI和NotI双酶切,连接并经测序验证后转化入大肠杆菌Rosetta中表达。制备粗酶液后用SDS-PAGE检测。单独测定粗酶液中2种酶的酶活:GDH酶活测定体系为1 mmol/L NADP+,10 mmol/L底物葡萄糖,pH 7.0 Na3PO4缓冲液,加入适量粗酶液启动反应。于30 ℃下测定波长340 nm处吸光值的上升率。酶活定义:每分钟内还原1μmol NADP+所需要的酶量为一个酶活单位U。
1.2.12生物转化COBE至(R)-CHBE
低温离心收集发酵培养基培养的含重组质粒pET-cak-gdh的双酶共表达重组菌,以20 g/L的添加量添加入反应体系中。反应体系包括200~600 mmol/L的底物COBE、800 mmol/L的葡萄糖以及Na3PO4缓冲液。转化反应在恒温摇床200 r/min的条件下进行,并定时调节pH至初始值。
1.2.13底物及产物的气相检测方法
COBE与CHBE浓度采用气相色谱仪(Agilent 7820A)测定。PEG20M毛细管柱(20 m×0.32 mm×0.25 μm);载气为N2,分流比1∶ 20;气化和检测室的温度220 ℃,梯度升温(初始温度130 ℃,保持3 min;以20 ℃/min程序升温至135 ℃,保持7 min;再以40 ℃/min程序升温至150 ℃,保持3 min;再以20 ℃/min程序升温至160 ℃,保持1 min);检测器为FID。COBE和CHBE的出峰时间分别为9.713 min和13.838 min。
产物对映体过量值(e.e.)的测定也同样使用气相色谱仪,分析条件:色谱柱CP-Chirasil Dex CB(25 m×0.25 mm×0.25 μm);载气为高纯氮,分流比1∶ 50;进样室和检测室温度250 ℃。(R)-CHBE和(S)-CHBE出峰时间分别为16.813 min和17.413 min。
2结果与讨论
2.1基因数据挖掘获得醛酮还原酶CAK
以目前文献报道的几种可用于不对称还原COBE合成(R)-CHBE的氧化还原酶(ARI、ByueD、gox2036、LEK、CmAR等)的氨基酸序列作为探针,在NCBI数据库中进行搜索,发现1种来自白色念珠菌Candidaalbican的假定蛋白,其氨基酸序列与氧化还原酶LEK的氨基酸序列相似度大于50%,据此认为该假定蛋白具有不对称还原制备(R)-CHBE的潜力。又经BLAST工具搜索,发现该蛋白的氨基酸序列与多种醛酮还原酶的氨基酸序列具有显著的相似性(超过40%相似性),这说明此蛋白属于醛酮还原酶超家族,将此酶命名为CAK。
2.2醛酮还原酶CAK的克隆、表达及纯化
将PCR扩增后的cak基因片段和质粒pET-22b(+)分别双酶切后进行连接,经测序验证完全正确。将得到的重组质粒命名为pET-cak。将pET-cak转化表达宿主大肠杆菌Rosetta,经发酵培养,超声破碎菌体得到了粗酶液。文献报道的醛酮还原酶绝大多数为NADPH辅酶依赖型,本实验酶活测定也选择NADPH作为辅酶,使用NADH作为辅酶检测不到任何活性。经计算,粗酶液比酶活为4.3 U/mg。粗酶液经纯化后,用SDS-PAGE分析,结果如图1所示。由图1可知,目标蛋白大小与预期一致(约3.3×104),纯化后的酶液比酶活达到9.3 U/mg。
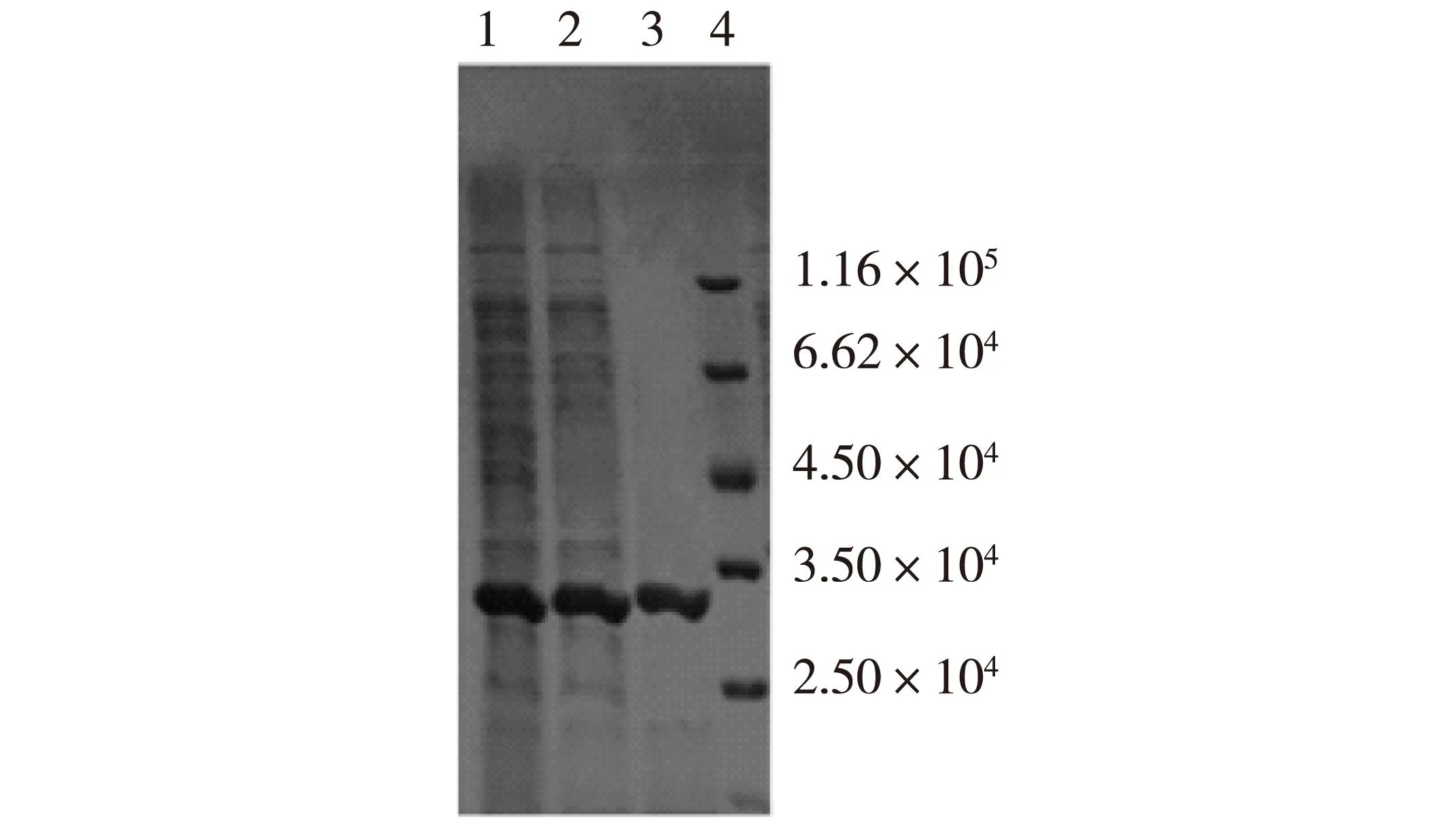
1—重组菌粗酶液; 2—经(NH4)2SO4初步纯化;3—经疏水层析纯化; 4—标准蛋白质图1 蛋白纯化SDS-PAGE电泳图Fig.1 SDS-PAGE analysis of purified CAK
2.3酶的底物谱测定
醛酮还原酶的底物谱主要由醛类和酮类化合物构成,CAK的底物谱测定结果如表1所示。由表1可见,CAK对于醛类的底物谱较宽,对多种醛类有催化活力且活力较高。其对丙酮醛的催化活力最高,
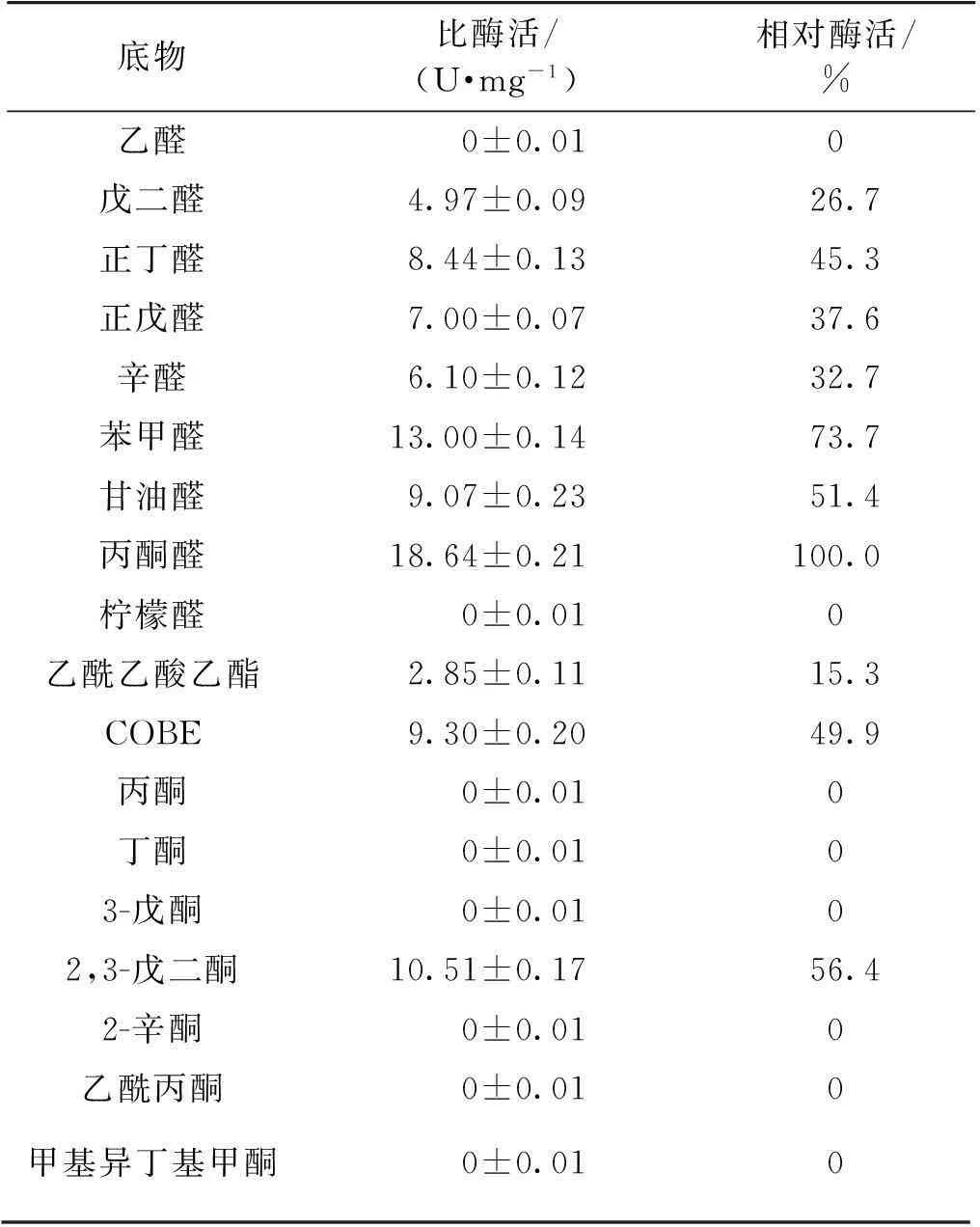
表1 CAK的底物谱测定
酶活达到18.64 U/mg。相比较而言,CAK能够催化的酮类化合物种类比较少,在所有测定的酮类化合物中,除乙酰乙酸乙酯、COBE、2,3-戊二酮这类带有α酮基的化合物外,其他酮类均测不到活性。
2.4温度对酶活的影响结果
考察温度(20~50 ℃)对CAK酶活的影响,结果如图2所示。由图2可知:在40 ℃时,醛酮还原酶CAK的活力达到最高;低于40 ℃时,酶活力受到了抑制;而当温度超过40 ℃时,虽然较高的温度可使酶促反应速率加快,但是此时蛋白变性对酶活的抑制作用更加明显,所以酶活力开始呈现下降趋势。
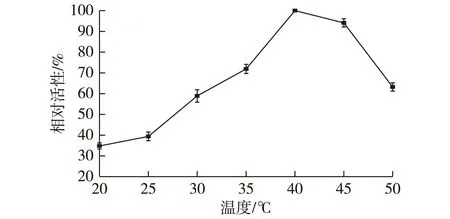
图2 温度对CAK酶活的影响Fig.2 Effects of temperature on activity of CAK
2.5CAK的热稳定性
不同的温度条件下研究CAK的热稳定性,结果见图3。由图3可知:其在40 ℃以下能保持较好的稳定性;而当在45 ℃条件下保温0.5 h后,酶活几乎完全丧失。这表明在超过45 ℃的条件下,酶的空间结构很容易被破坏。Wang等[17]报道了一种Lodderomyceselongisporus来源的用于制备(R)-CHBE的醛酮还原酶,其在40 ℃下保温0.5 h后仅有20%的残余酶活,相比较而言,CAK的热稳定性要略高一些。
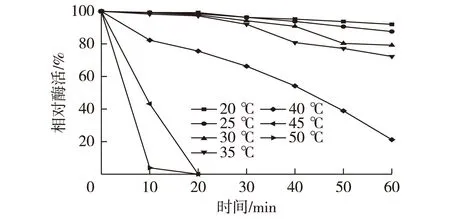
图3 CAK的温度稳定性曲线Fig.3 Thermostability of CAK
2.6pH对酶活的影响
考察不同pH对醛酮还原酶的酶活影响,结果见图4。由图4可知:在pH为5时酶活最高,在pH 4~7之间,酶活力保留了最大酶活的70%以上。在碱性条件下,酶活会受到较大程度的抑制。这可能是因为在偏酸性的环境中,CAK酶分子活性中心的空间构象更有利于与底物的结合,表现出了更高的催化活性。
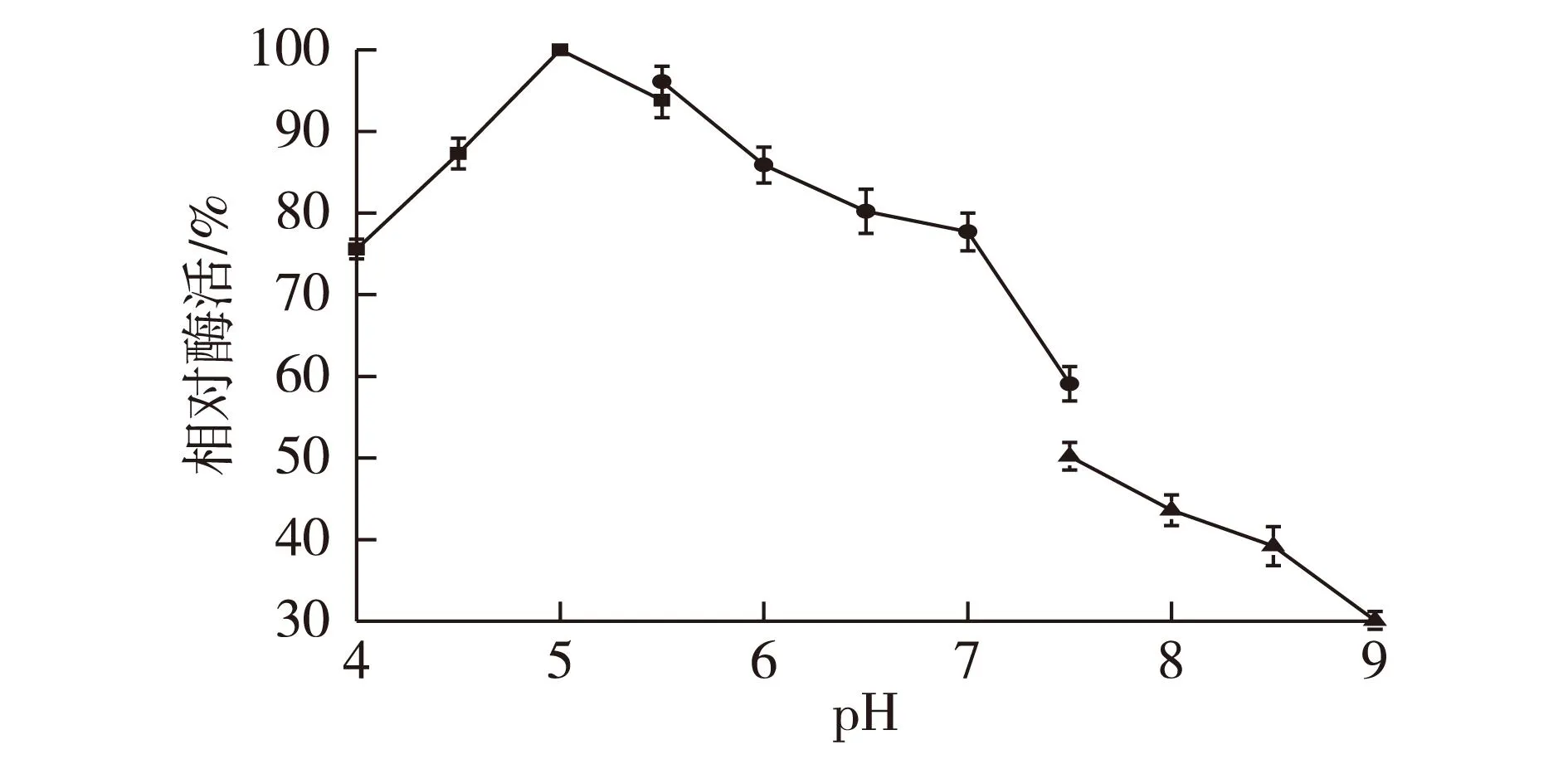
图4 pH对CAK酶活的影响Fig.4 Effects of pH on activity of CAK
2.7CAK的pH稳定性
图5比较了酶在不同pH条件下保存24 h后的残余酶活。由图5可知:CAK在酸性条件下较为稳定。而当pH大于8时,酶活损失严重。其原因可能是在碱性环境下,OH-浓度的增加,使得酶活性中心的结构更容易发生不可逆的改变,从而导致其催化活力急剧降低。因此,CAK不宜保存在碱性的缓冲液中。
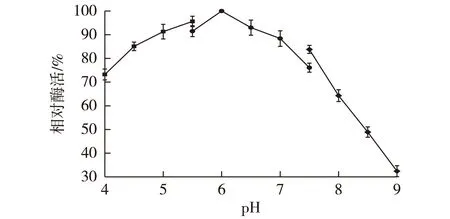
图5 CAK的pH稳定性曲线Fig.5 pH stability of CAK
2.8金属离子对CAK酶活的影响
考察多种金属离子对CAK酶活的影响,结果见表2。由表2可知:其中Mg2+、Na+、K+对酶活有一定的激活作用,其余金属离子均对酶活有抑制作用,其中添加Cu2+使酶完全失活。该结果表明,CAK对某些金属离子尤其是Cu2+较为敏感,少量的Cu2+就可以使酶的空间结构破坏,导致其完全丧失活性,因此,在催化体系中应该避免Cu2+的引入。

表2 金属离子对酶活的影响
2.9有机溶剂对CAK酶活的影响
催化转化COBE的反应中,为增加底物的溶解性,降低底物和产物对酶的毒害作用,通常会加入有机溶剂,但有机溶剂本身也可能会抑制酶活性[18]。COBE和CHBE在乙酸乙酯、乙酸丁酯、辛酸乙酯、三氯甲烷、邻苯二甲酸二丁酯、异丙醚这几种有机溶剂中的油水分配系数都较高,表明这些有机溶剂具备用于有机-水双相催化制备CHBE的潜力。本实验考察上述有机溶剂及其加入量对酶活的影响,以不加有机溶剂体系的相对酶活为100%,结果见图6。由图6可知:所有的有机溶剂均对酶活有抑制作用,其中乙酸乙酯、邻苯二甲酸二丁酯的抑制作用相对较小,当有机相体积分数达到50%,时CAK尚有60%左右的残余酶活。
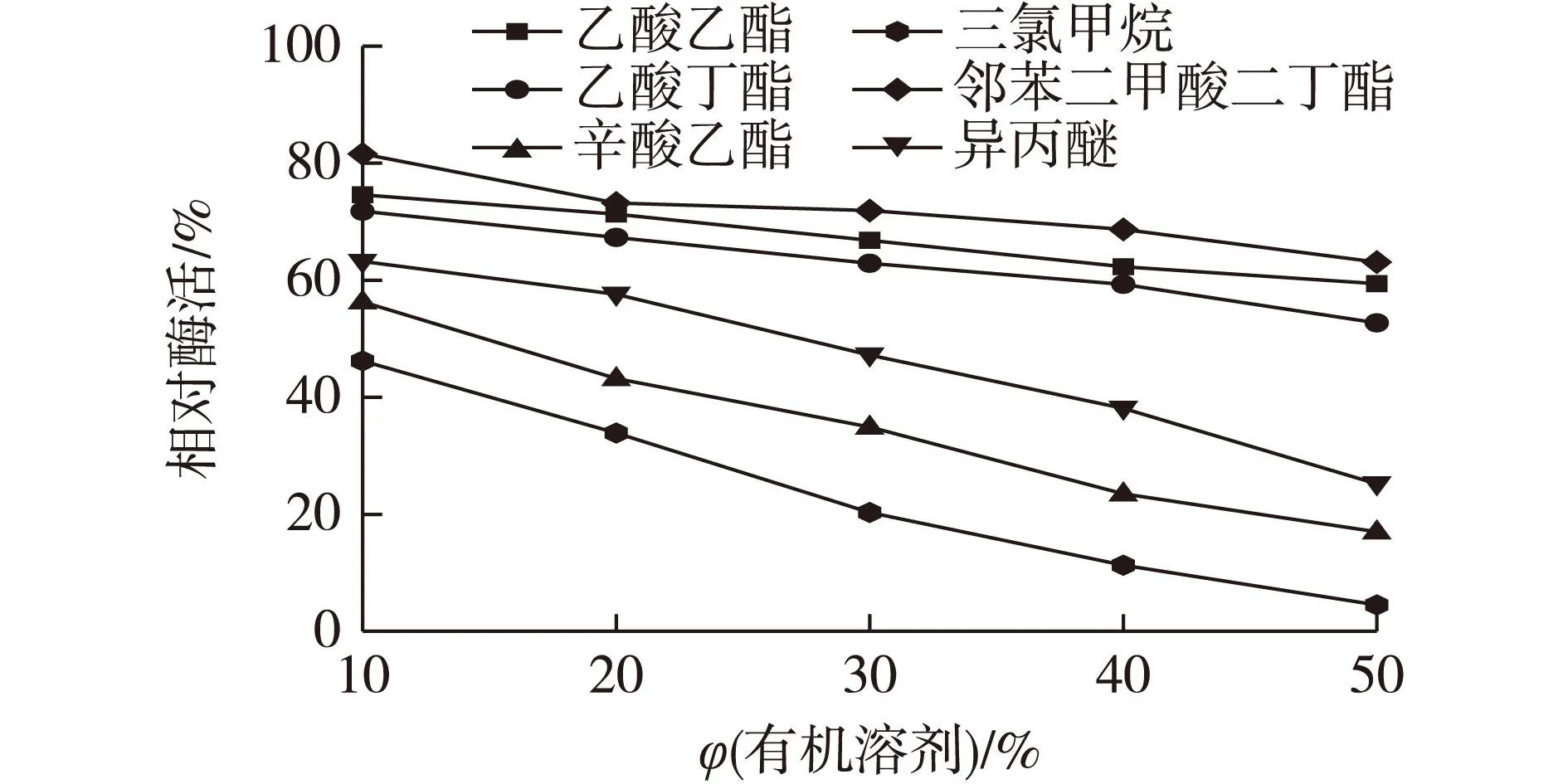
图6 有机溶剂对酶活的影响Fig.6 Effects of organic solvents on activity of CAK
2.10CAK和GDH的共表达及酶活测定
双酶共表达重组菌粗酶液经SDS-PAGE鉴定,结果见图7。由图7可见,目的条带明显,相对分子质量与预期一致,2种酶在大肠杆菌中均得到了良好的可溶性表达。经计算,粗酶液中CAK和GDH的酶活分别为2.5和6.2 U/mg(以1 mg蛋白计)。
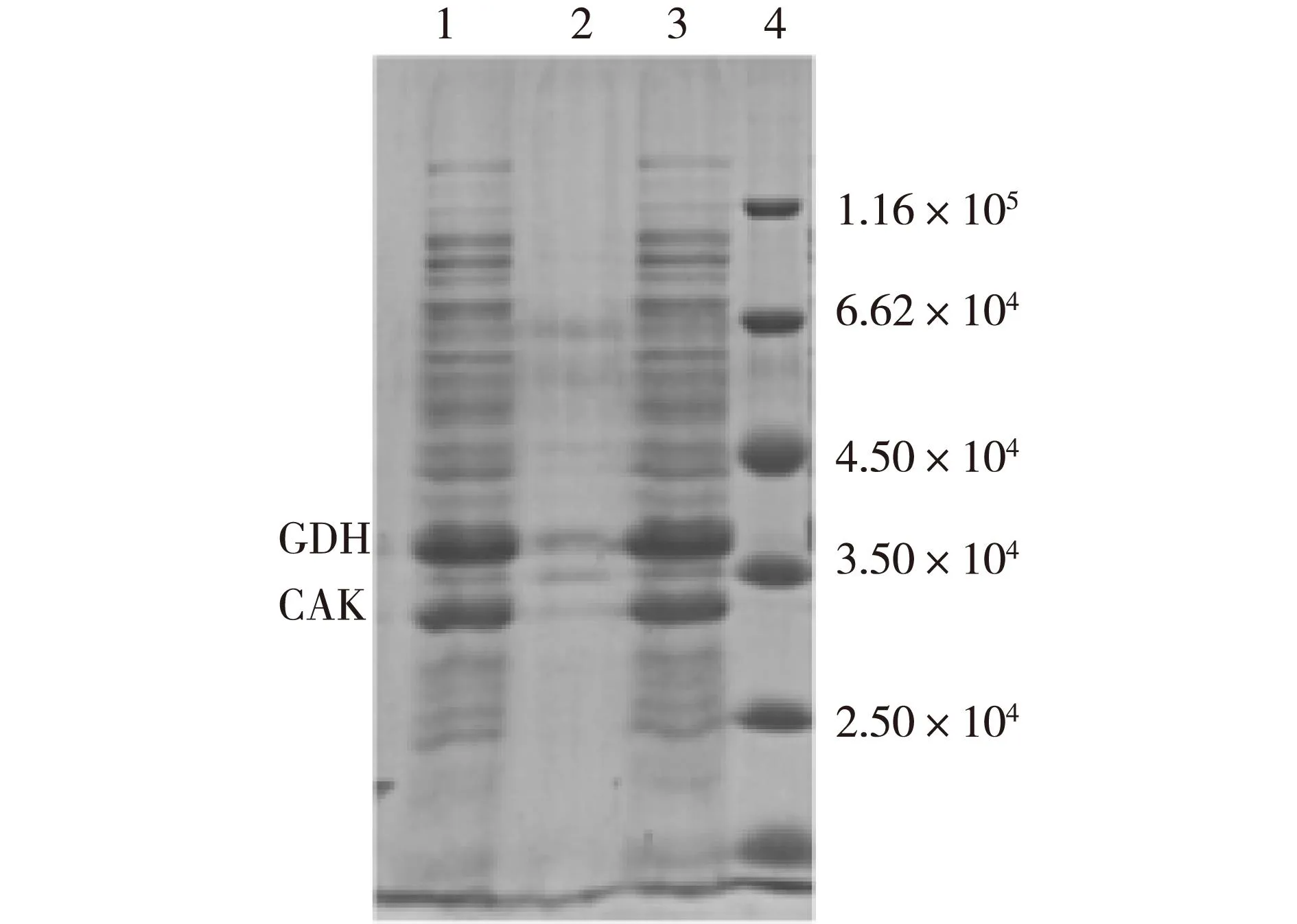
1—双酶共表达粗酶液; 2—细胞破碎液(沉淀); 3—细胞破碎液(全菌); 4—标准蛋白质图7 双酶共表达蛋白电泳Fig.7 SDS-PAGE analysis of co-expressing
2.11底物浓度对辅酶再生系统不对称催化还原COBE的影响
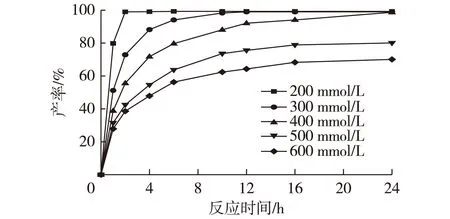
图8 底物浓度对辅酶再生体系催化 合成(R)-CHBE的影响Fig.8 Effects of COBE concentration on the synthesis of (R)-CHBE by coenzyme regeneration system
利用醛酮还原酶CAK和葡萄糖脱氢酶共表达的重组大肠杆菌细胞,以及在催化体系中添加的辅底物葡萄糖来实现辅酶再生。反应体系的pH设定为6.0,温度设定为35 ℃。图8表示了不同底物浓度下,双酶共表达重组菌全细胞催化还原COBE的产率随时间变化的曲线。由图8可知:在底物浓度为400 mmol/L以下时,经过24 h的催化反应,底物几乎都能转化完毕(产率大于99%)。当底物浓度超过500 mmol/L时,反应24 h的产率降低至80%以下,这可能是因为体系中积累了高浓度的产物CHBE,抑制了醛酮还原酶CAK的活性,使反应难以继续进行。测定最终产物的对映体过量值(e.e.值),结果均在99%以上。
2.12反应温度对辅酶再生系统不对称催化还原COBE的影响
考察温度对催化体系的影响,采用较高的初始底物浓度(600 mmol/L),pH设定为6.0,经24 h反应,测定并计算最终的产率,结果如图9所示。由图9可见,30 ℃的温度条件下产率最高。较高和较低的反应温度均不利于转化,这是因为低温(低于30 ℃)的转化条件降低了酶促反应速率,必然会使最终产率降低。而较高的温度则会使酶失活,另外,温度越高,底物和产物也就越容易发生水解,这些原因都导致了最终产率的降低。
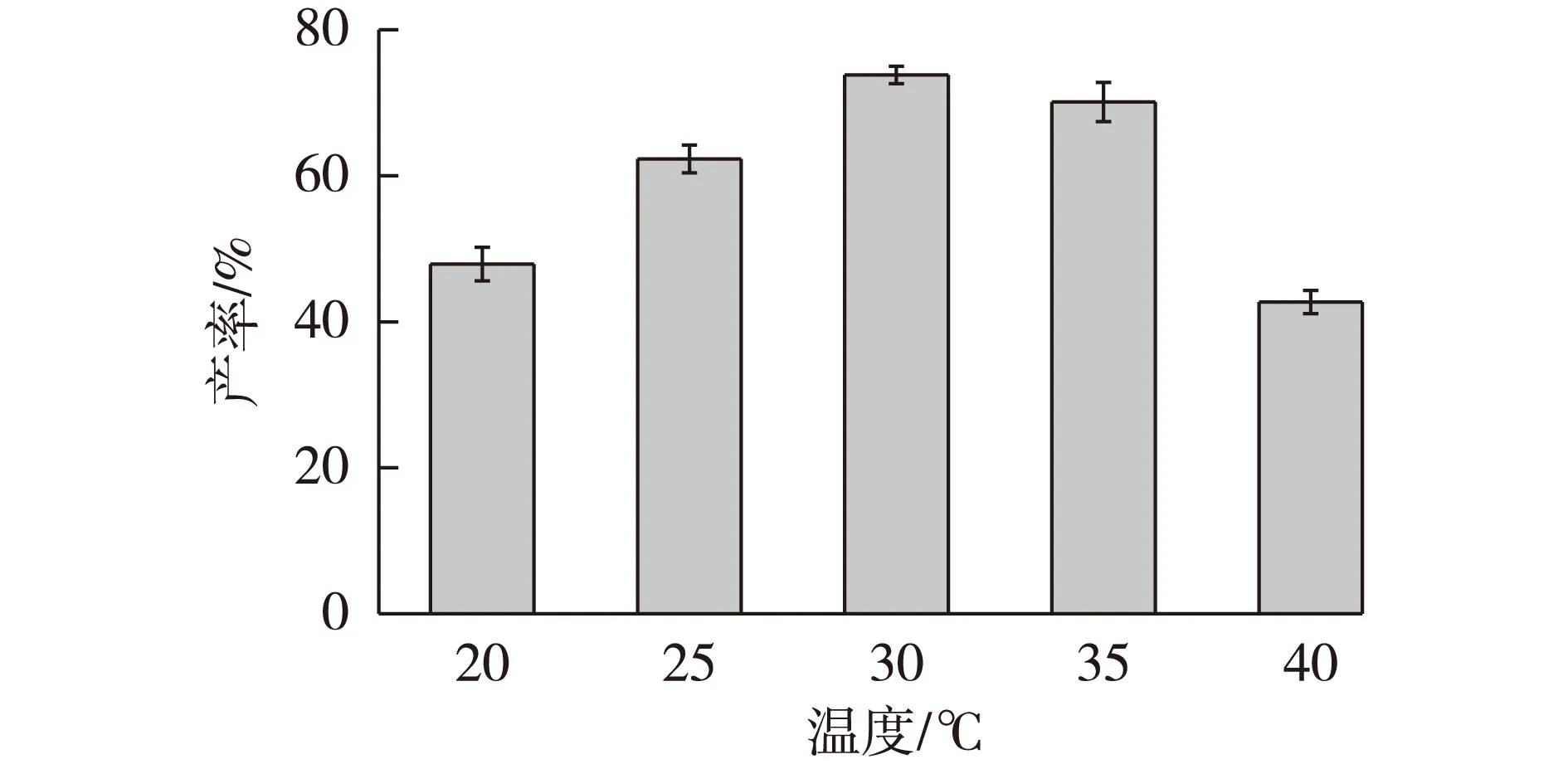
图9 反应温度对辅酶再生体系催化 合成(R)-CHBE的影响Fig.9 Effects of temperature on the synthesis of (R)- CHBE by coenzyme regeneration system
2.13pH对辅酶再生系统不对称催化还原COBE的影响
考察不同pH的缓冲体系(pH 4~8)用于催化反应,转化600 mmol/L底物,于30 ℃反应24 h后,测定并计算产率,结果见图10。由图10可知:当反应体系的pH为7时可达到最高产率(80.6%),这与CAK的最适pH(pH为5)有一定出入。这是因为GDH在pH大于7时才能表现出高活性,在酸性条件下其酶活力较低,使得辅酶再生的效率有所下降。
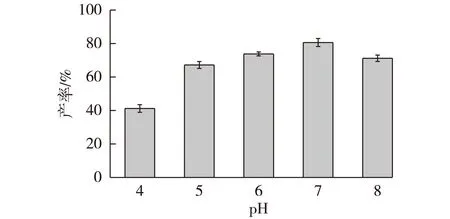
图10 pH对辅酶再生体系催化合成 (R)-CHBE的影响Fig.10 Effects of pH on the synthesis of (R)-CHBE by coenzyme regeneration system
3结论
从白色念珠菌中克隆了1种具有不对称还原COBE的醛酮还原酶CAK,并在大肠杆菌中成功表达。对CAK进行纯化后,研究其酶学性质:CAK对多种醛酮类化合物有催化活性;其催化COBE的最适反应温度为40 ℃,最适pH为5;酶在40 ℃以下能保持较好的热稳定性,在弱酸性条件下酶的性质最稳定,碱性环境中酶活损失较大;Mg2+、Na+、 K+的添加对酶有激活作用;多种有机溶剂均对酶活有抑制作用,而乙酸乙酯、邻苯二甲酸二丁酯对CAK的酶活影响相对较小。通过CAK和葡萄糖脱氢酶在大肠杆菌中共表达,构建辅酶再生体系,利用该体系催化不对称合成(R)-CHBE,在最适的转化条件下(30 ℃,pH=7)转化600 mmol/L的底物,产率达80.6%,且产物的e.e.值超过99%,表明这种制备(R)-CHBE的方式具有良好的应用前景。
参考文献:
[1]DAVIES S G,LCHIHARA O.Asymmetric synthesis of (+)-negamycin[J].Tetrahedron:Asymmetry,1996,7:1919-1922.
[2]MIYASHITA A,YASUDA A,TAKAYA H,et al.Synthesis of 2,2′-bis(diphenylphosphino)-1,1′-binaphthyl (BINAP),an atropisomeric chiralbis(triaryl)phosphine,and its use in the rhodium(I)-catalyzed asymmetric hydrogenation of alpha-(acylamino) acrylic acids[J].J Am Chem Soc,1980,102(27):7932-7934.
[3]HIKICHI S,HAREAU G P J,SATO F.Efficient and practical synthesis of optically active 5-t-butyldimethylsiloxy-2-c ̄y ̄c ̄l ̄o ̄h ̄e ̄x ̄e ̄n ̄o ̄n ̄e as a convenient chiral 2,5-cyclohexadienone synthon[J].Tetrahedron Lett,1997,38:8299-8302.
[4]WADA M,KAWABATA H,KETAOKA M,et al.Purification and characterization of an aldehyde reductase fromCandidamagnoliae[J].J Mol Catal B:Enzymatic,1999,6:333-339.
[5]KITA K,NAKASE K,YANASE H,et al.Purification and characterization of new aldehyde reductases fromSporobolomycessalmonicolorAKU4429[J].J Mol Catal B:Enzymatic,1999,6:305-313.
[6]MATSUYAMA A,YAMAMOTO H,KOBAYASHI Y.Practical application of recombinant whole-cell biocatalysts for the manufacturing of pharmaceutical intermediates such as chiral alcohols[J].Org Proc Res Develop,2002,6:558-561.
[7]SHUMIZU S,KATAOKA M,MORISHITA M,et al.Microbial asymmetric reduction of ethyl-4-chloro-3-oxobutanoate to optically active ethyl-4-chloro-3-hydroxybutanoate[J].Biotechnol Lett,1990,12(8):593-596.
[8]YU M A,WEI Y M,ZHAO L,et al.Bioconversion of ethyl 4-chloro-3-oxobutanoate by permeabilized fresh brewer’s yeast cells in the presence of allyl bromide[J].J Ind Microbiol Biotechnol,2007,34:151-156.
[9]NI Y,LI C X,WANG L J,et al.Highly stereoselective reduction of prochiral ketones by a bacterial reductase coupled with cofactor regeneration[J].Org Biomol Chem,2011,9:5463-5468.
[10]JING K,XU Z,LIU Y,et al.Efficient production of recombinant aldehyde reductase and its application for asymmetric reduction of ethyl 4-chloro-3-oxobutanoate to ethyl (R)-4-chloro-3-hydroxybutanoate[J].Prep Biochem Biotechnol,2005,35:203-215.
[11]LIU X,CHEN R,YANG Z W,et al.Characterization of a putative stereoselective oxidoreductase fromGluconobacteroxydansand its application in producing ethyl (R)-4-chloro-3-hydroxybutanoate ester[J].Mol Biotechnol,2014,56:285-295.
[12]HE Y C,ZHANG D P,TAO Z C,et al.Discovery of a reductase-producing strain recombinantE.coliCCZU-A13 using colorimetric screening and its whole cell-catalyzed biosynthesis of ethyl (R)-4-chloro-3-hydroxybutanoate[J].Bioresour Technol,2014,172:342-348.
[13]KATAOKA M,ROHANI L P S,YAMAMOTO K,et al.Enzymatic production of ethyl (R)-4-chloro-3-hydroxybutanoate:asymmetric reduction of ethyl 4-chloro-3-oxobutanoate by anEscherichiacolitransformant expressing the aldehyde reductase gene from yeast[J].Appl Microbiol Biotechnol,1997,48(6):699-703.
[14]SAMBROOK J,FRISCH E F,MANIATIS T.Molecular cloning:a laboratory manual[M].New York:Cold Spring Harbor Laboratory Press,1989.
[15]BRADFORD M M.A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding[J].Anal Biochem,1976,72(1):248-254.
[16]GREEN M R,SAMBROOK J.Molecular cloning:a laboratory manual[M].New York:Cold Spring Harbor Laboratory Press,2012.
[17]WANG Q,YE T,MA Z,et al.Characterization and site-directed mutation of a novel aldo-keto reductase fromLodderomyceselongisporusNRRL YB-4239 with high production rate of ethyl (R)-4-chloro-3-hydroxybutanoate[J].J Ind Microbiol Biotechnol,2014,41(11):1609-1616.
[18]何军邀,孙志浩,郑璞,等.水/有机溶剂两相体系微生物催化不对称还原制备(S)-4-氯-3-羟基丁酸乙酯[J].高校化学工程学报,2007,21(2):310-315.
(责任编辑荀志金)
Overexpression,characterization of a novel aldo-keto reductase for asymmetric synthesis of (R)-CHBE
ZHANG Jincheng,WEI Miao,XU lin,YAN Ming
(College of Biotechnology and Pharmaceutical Engineering,Nanjing Tech University,Nanjing 211800,China)
Abstract:An NADPH-dependent aldo-keto reductase(CAK)gene from Candida albicans SC5314 was discovered for reducing 4-chloroacetoacetate ethyl ester (COBE) to ethyl (R)-4-chloro-3-hydroxybutanoate ((R)-CHBE). The CAK gene was cloned and overexpressed in Escherichia coli Rosetta. The catalytic properties of purified CAK were studied. Furthermore,an enzyme-coupled coenzyme regeneration system with co-substrate glucose was constructed,and its capability of reducing COBE to (R)-CHBE was studied. The results showed that CAK had catalytic activity for a variety of aldehydes and ketones. CAK had a maximum activity at 40 ℃ and pH 5. The enzyme was stable below 40 ℃ and under acidic conditions. Mg(2+),Na+ and K+ could enhance the activity of CAK. When Cu(2+) was added, no enzyme activity was detected. The inhibit effect of ethyl acetate and dibutyl-O-phthalate was relatively light. When the coenzyme regeneration system for conversion reaction was applied,the molar conversion yield reached 80.6% with 600 mmol/L COBE,and the e.e. value was over 99%.
Keywords:aldo-keto reductase; NADPH regeneration;protein purifying; enzyme property;(R)-4-chloro-3-hydroxybutanoic acid ethyl ester
中图分类号:Q815
文献标志码:A
文章编号:1672-3678(2016)02-0033-08
作者简介:张瑾成(1989—),男,江苏无锡人,研究方向:生物催化;严明(联系人),副教授,E-mail :yanming@njtech.edu.cn
基金项目:国家重点基础研究发展计划(973计划)(2011CBA00804);国家高技术研究发展计划(863计划)(2012AA022101);江苏高校优势学科建设工程
收稿日期:2015-03-17
doi:10.3969/j.issn.1672-3678.2016.02.007
