可可广翅蜡蝉的行为节律观察
2016-03-01范广玉刘宁国刘学敏陈文龙
范广玉, 刘宁国, 杨 群, 刘学敏, 陈文龙
(贵州大学 昆虫研究所, 贵州山地农业病虫害重点实验室, 贵州 贵阳 550025)
可可广翅蜡蝉的行为节律观察
范广玉, 刘宁国, 杨 群, 刘学敏, 陈文龙*
(贵州大学 昆虫研究所, 贵州山地农业病虫害重点实验室, 贵州 贵阳 550025)
为进一步做好可可广翅蜡蝉的预测预报及防治工作,采用室内饲养观察的方法对其羽化、交配、产卵的行为及节律进行研究,并对其田间产卵特征进行调查。结果表明:可可广翅蜡蝉成虫羽化发生在晚上和凌晨,白天未见羽化;成虫交配主要发生在白天,晚上很少交配,凌晨未见;成虫全天均可产卵,高峰期集中在上午与凌晨;田间主要将卵产在茎粗为1.5 mm左右的嫩茎上,产卵深度约0.9 mm,产卵刻痕主要集中在距顶梢3 cm范围,平均每刻痕卵量约为11粒。
可可广翅蜡蝉; 羽化; 交配; 产卵; 行为节律
生物的生理活动及外在行为具有一定的周期节律特征[1-4]。其昼夜节律具有种的特异性,这不仅是昆虫个体间基因交流的基础,也是生产上进行预测及防治的重要依据[5-8]。近年来,随着气候条件、茶园生态环境、茶树品种及其栽培方式的改变,茶树蜡蝉种群正逐渐从次要害虫变为主要害虫[9],截止目前,有关蜡蝉种类如柿广翅蜡蝉(RicaniasublimbataJacobi)、八点广翅蜡蝉(R.speculumWalker)、碧蛾蜡蝉(GeishadistinctissimaWalker)、白蛾蜡蝉(LawanalimitataMelichar)、缘纹广翅蜡蝉(R.marjinlisWalker)等的行为节律均已有相关报道[10-33]。
可可广翅蜡蝉(RicaniacacaonisWalker)在贵州的具体发生及危害状况尚未见报道,笔者通过调查发现,该虫在贵州都匀市、贵阳市均有发生,其危害程度每年呈上升趋势,尤其在贵阳黔陶地区危害严重。因此,为做好该虫预报及防治工作,探明其行为节律意义重大。邵元海等[9,34-35]对该虫的生态学及防治进行报道,但对其生物学及行为节律研究较少,且多局限于江苏地区,而地域性差异致使该虫在不同地区的生活习性必然有所不同。因此,探明可可光翅蜡蝉在贵州地区的生物学及生态学特性是实现当地对该虫的预测预报及有效防治的重要途径,而其行为节律更是重中之重。为此,笔者于2013年采用室内饲养观察结合田间调查的方法对其羽化、交配、产卵的行为及节律进行研究,以期为该虫的预测预报及有效防治提供理论依据。
1 材料与方法
1.1 研究对象
以可可光翅蜡蝉为研究对象,虫卵采自贵阳市花溪熙皇赵司贡茶茶场。
1.2 试验时间及地点
试验于2013年3—10月在贵州大学昆虫研究所与贵阳市花溪熙皇赵司贡茶茶场(海拔1 300~1 500 m的山坡上,新老茶园共2 00 hm2)进行。
1.3 成虫的行为节律观察
1.3.1 成虫羽化 在若虫孵化前,将剪自茶园的带卵枝条基部用湿棉团包裹,置于300 mL的透明塑料杯中,杯口用纱网封盖,待若虫孵化后使用嫩茶枝进行单头饲养,记录蜕皮次数,对蜕皮至末龄的若虫进行不定时观察。自有虫羽化开始,每2 h观察1次,记录每次观察所发生羽化的虫口数量,并对其羽化行为进行拍照描述,共100次重复。
1.3.2 成虫交配 将采自茶园的雌、雄成虫进行配对,置于装有茶树嫩枝的300 mL塑杯进行饲养,每杯1对,共50次重复。嫩枝基部用湿棉团包裹并对茶枝进行不定期更换,以保证枝叶新鲜。每2 h观察1次,记录每次观察发生交配的虫口对数,并对其交配行为进行拍照描述。
1.3.3 雌成虫产卵 1) 室内行为习性观察。将已发生交配的雌成虫置于300 mL的塑杯进行单头饲养,取直径1.5 mm左右的茶树嫩枝,将叶片剪除(以便于观察产卵行为),其基部用足够大的湿棉团包裹,置于杯底并固定,供成虫产卵;将剪取的叶片取2~3片置于杯中,对叶片进行每日更换以保证新鲜,供雌成虫取食。每日观察其产卵情况,自有虫产卵开始,每3 h观察1次,记录每次观察发生产卵的虫口数,并对其产卵行为进行拍照描述。2) 野外产卵特征调查。于产卵期过后,取一块茶树种植密度大、生长年龄长的样地,按Z字取样法抽取30株茶树,每株随机抽取1枝具3~4个分支的枝条,记录每枝条的产卵刻痕数,每刻痕所在部位,每刻痕距枝条顶梢距离,每刻痕深度,每刻痕卵粒数,每卵粒间距。
1.4 数据统计与分析
试验数据分析应用软件为EXCEL 2003及SPSS 18.0。
2 结果与分析
2.1 成虫羽化(封二图Ⅰ)
当5龄若虫到达末期准备发生羽化时,胸背裂开一条较窄缝隙,但并未完全裂开,裂缝处仍有一层薄膜相连,此时若虫仍有运动能力,但常栖居于一处静止不动,等待羽化。在发生羽化时,其腹部伸直拉长,头部顶壳裂开,其头部慢慢自裂缝处钻出。之后随着腹部不断的进行前后蠕动,裂缝不断加深,沿胸背中脊向胸背末端逐渐延伸,直至腹部第二腹节处停止,在此过程中其翅随着胸背的露出而逐渐显化,前、中、后足亦慢慢自原来体壳中抽出。当翅刚显化出来时并未伸展,其近外缘处向下弯曲,折叠于胸部两侧,形似蘑菇。待后足完全抽出后,腹部暂停蠕动约30 s,之后前、中、后足开始慢慢摆动,前足向前方伸展,超过头部,中足做前后重复性摆动,后足向两侧张开,呈150°~180°角,并且上翘顶住翅的边缘,促进翅的伸展,随之整个躯体自原来体壳中爬出。
以上整个过程,从头部顶壳裂开到整个躯体钻出体壳,需经40~60 min,此后再经10~20 min,翅慢慢伸展开来。至此,整个羽化过程结束。刚羽化的成虫通体乳白,前、后翅均透明,翅脉不甚清晰,亦不见斑纹、刻点,此时成虫常停留在皮壳前方,不进行运动取食,之后随时间推移,前翅颜色逐渐加深,由透明的白色逐渐转为较透明的灰色,进而转为不透明的黑灰色或黄褐色,翅脉逐渐清晰,其翅上的各种斑纹和刻点亦慢慢显化,而后翅亦由原来的白色透明转为浅灰色透明。成虫从刚羽化完成到发展为骨骼坚硬、能飞善蹦的成虫约需要7~10 h。
从图1可见,可可广翅蜡蝉羽化均发生在晚上和凌晨,白天不发生羽化。其中,羽化高峰期集中在晚上22:00~00:00及凌晨00:00~02:00,二者之间无显著差异,但显著高于其他时间段;其次为20:00~22:00与02:00~04:00,而18:00~20:00及04:00~06:00羽化量较少。
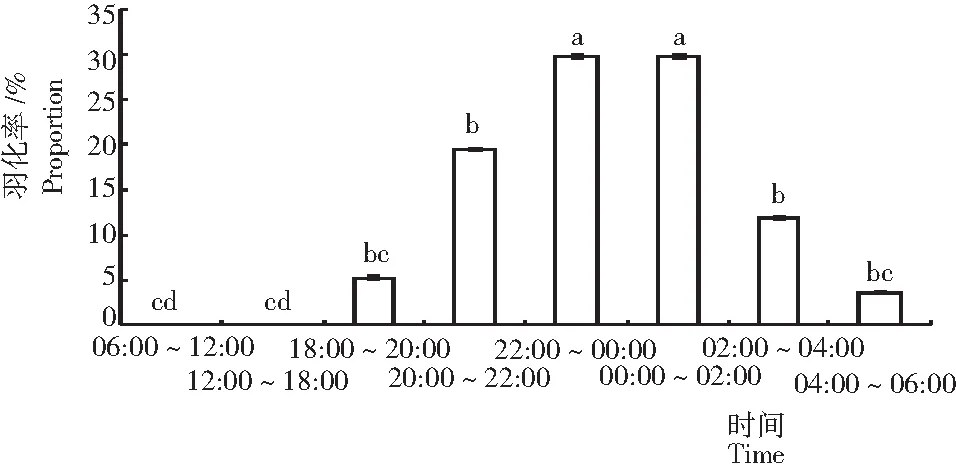
注:图中小写字母代表α=0.05水平的显著性差异(Duncan法)(下同)。
Note: Lowercase letters in the Fig. indicate significance of difference at 0.05 level. The same below.
图 1 可可广翅蜡蝉的羽化节律
Fig.1 Emergence rhythm ofR.cacaonis
2.2 成虫交配(封二图Ⅱ)
在进行交配时,雌、雄虫身体以尾节为顶角保持140°~180°,雄虫尾节向一侧弯曲,之后抱器张开,紧握雌虫第九腹板基部的凹陷处,为其交配提供着力点,此时阳茎外露。雄虫通过雌虫交配孔使阳茎插入雌虫体内,此交配孔位于雌虫第7与第8腹节之间的节间膜处,阳茎经过不断抽插后直至交配囊,此时雄虫尾节后上角便抵在雌虫交配孔旁边的节间膜处,当雄虫阳茎停止抽插后,其肛节紧扣雌虫第7腹板,并可见雄虫肛刺突轻微晃动。可可广翅蜡蝉在交配过程中如遇惊扰,可进行小范围的爬行或跳跃,此后交配行为仍然继续。在交配即将结束时,雌虫身体进行轻微的左右晃动,后足凌空轻踢,身体向前爬行,此时尾节两侧腹板张开,促使雄虫将阳茎拔出体外,约1~3 min后雄虫抱器松开,阳茎自雌虫体内抽出,雄虫抱器与肛节各归原位,雌虫尾节两侧腹板亦重新闭合,至此交配行为结束,整个交配过程约需40~70 min。
从图2可见,可可广翅蜡蝉成虫的交配主要发生在白天,晚上很少有虫交配,而在凌晨未发现有虫交配,交配高峰期发生在上午08:00~12:00,且以08:00~10:00居多,显著高于其他时间段。下午以14:00~16:00发生交配的虫口数居多,但与12:00~14:00及16:00~18:00时间段之间差异不显著。
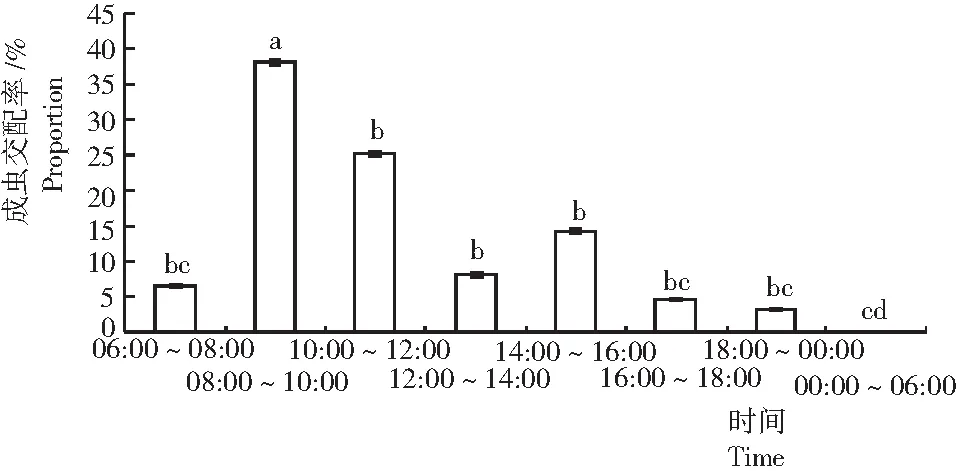
图2 可可广翅蜡蝉的交配节律
2.3 产卵(封二图Ⅲ)
2.3.1 产卵行为及节律观察 可可广翅蜡蝉雌虫自交配至产卵所需时间浮动较大,短则需要2 d,长则需要5 d,其产卵须经刻槽、产卵、掩埋3个阶段。在产卵时腹部末端弯曲下垂,与产卵枝条垂直,第九腹板张开,第1、2产卵瓣外露,形如利刀,两对产卵瓣交替钻刻和切割枝条,一边切割一边将木屑挖出,最终形成能够盛卵的倾斜刻槽,角度约45°。待形成的刻槽适合盛卵时产卵瓣便停止切割,此时腹部第九节两侧腹板张开将枝条夹住,接着腹部不断地收缩与舒张,将卵通过产卵孔排出体外,斜产于凹槽之内,卵的尖端朝上,膨大一端朝下。待将卵盛好后第九腹板松开枝条,产卵瓣将挖出的木屑覆于卵上,至此整个产卵过程结束。产卵需10~13 min/粒, 待一颗卵产完之后虫体向前爬行约 0.8 mm 继续产卵,最终形成鱼鳍状的条形刻痕。
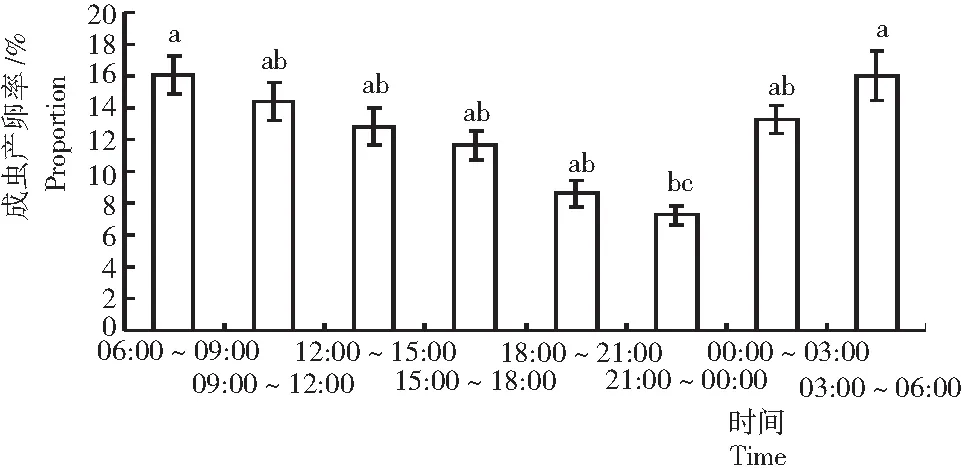
图3 可可广翅蜡蝉的产卵节律
从图3可见,可可广翅蜡蝉全天均可产卵,产卵高峰期集中在上午与凌晨,占整日产卵量的60%以上;自早晨06:00至晚上00:00产卵量逐渐降低,之后进入凌晨后产卵量又逐渐升高。
2.3.2 野外产卵特征 可可广翅蜡蝉的产卵刻痕呈条形,为土黄色,每卵粒间呈鱼鳍状排列。从表1可见,每枝条平均刻痕数为2~4个,而分布于茎上的刻痕数为2~3个,为叶片上刻痕数的5~8倍。对于茎上的产卵刻痕,平均每刻痕含11~13粒卵,卵粒间距约为0.73 mm;对于叶片上的产卵刻痕,平均每刻痕含7~9粒卵,其卵粒数少于茎上刻痕,但其产卵粒间距约为1.00 mm左右,大于茎上刻痕的卵粒间距。
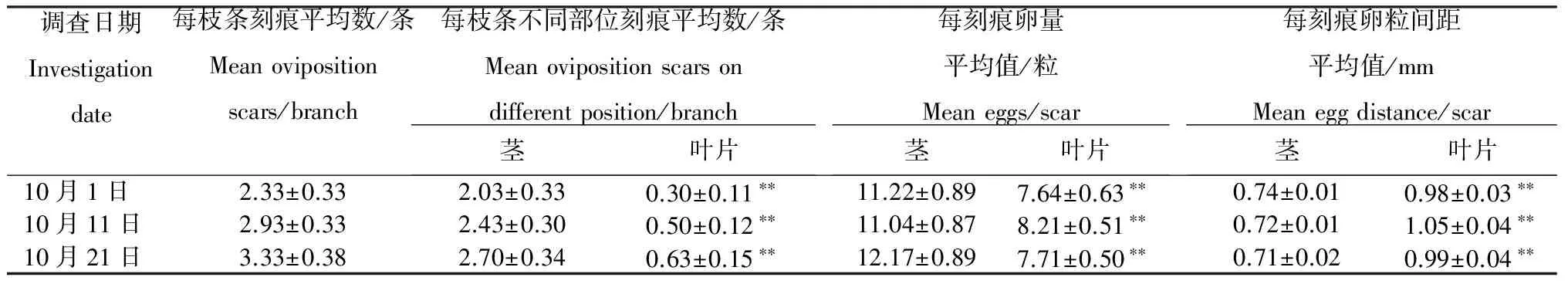
表1 可可广翅蜡蝉的产卵特征
注:*表示经t-检验差异显著(P<0.05), **表示经t-检验差异极显著 (P<0.01)。
Note: * and ** indicates significance of difference atP<0.05 andP<0.01 level respectively.
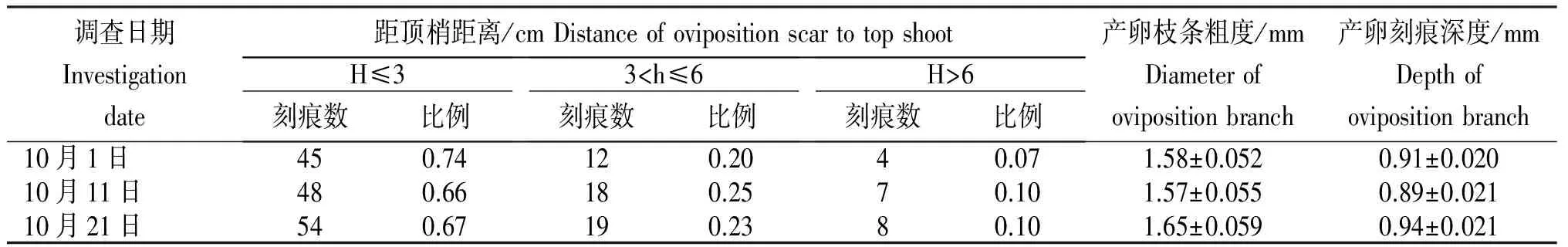
表2 茎上可可广翅蜡蝉产卵刻痕的特征
由于可可广翅蜡蝉主要产卵于枝条嫩茎上,并且危害巨大, 所以进一步对该刻痕进行深入调查。 从表2可见,该虫产卵时选择的枝条粗度在1.5 mm左右,主要为当年抽发的已发生木质化或半木质化的较嫩枝条,其产卵深度约为0.9 mm,可深达木质部,严重影响枝条养分的运输。产卵刻痕主要分布在距梢顶≤3 cm的枝条上,占总刻痕数的65%~75%;其次为3~6 cm,占总刻痕数的20%~25%;而在>6 cm的枝条上产卵刻痕数很少, 仅在10%左右。
3 结论与讨论
通过对可可广翅蜡蝉3种行为节律的观察发现,成虫羽化主要集中在晚上和凌晨,白天不发生羽化,羽化高峰期集中在22:00~02:00,接近羽化总量的60%。成虫交配主要集中在白天,上午较多,占总交配对数的65%以上,且以08:00~10:00最多,可达35%~40%。雌成虫交配后短则2 d,长则5 d即可产卵,雌虫全天均可产卵,其产卵高峰期集中在上午与凌晨,占总产卵量的60%以上。
通过田间调查发现,该虫可将卵产于粗度为1.5 mm左右的嫩茎及叶片背面主脉上,但以嫩茎上的产卵刻痕数居多,约为叶片主脉的5~8倍。茎上每刻痕的平均卵量多于叶片,且卵粒密度也大于叶片,造成此种现象的原因可能是嫩茎直径大于叶片主脉,能够更好地对卵粒进行掩埋,从而提供优越的保护场所,易于越冬。对于嫩茎上的产卵刻痕,其刻痕平均深度为0.9 mm,主要分布在距顶梢3 cm以内的枝条上,约占总刻痕数量的65%~75%,造成此种刻痕分布的主要原因可能是越靠近顶梢,其茎干越嫩,不但易于成虫刺吸取食,更易于该虫产卵器划破枝条进行产卵。
本文对可可广翅蜡蝉的羽化、交配和产卵节律进行时间上的量化分析,并对其行为过程进行观察和描述,这虽然对该虫的田间防治及进一步科学研究均具有重要意义,但多局限于现象描绘,因此在下一步工作中,应对其内在行为机制进行深入研究,从而探明各行为的生理意义。
[1] 涂小云,陈元生.蛾类昆虫行为节律[J].生物灾害科学,2013,36(1):18-21.
[2] 吴少会,向 群,薛芳森.昆虫的行为节律[J].江西植保,2006,29(4):147-157.
[3] Pittendrigh C S.Temporal organization:reflection of Darwinian watcher[J].Annual Review of Physiology,1993,55:17-54.
[4] Takahishi J S.Molecular neurobiology and genetics of circadian rhythm in mammals[J].Annual Review of Neuroscience,1995,18:3531-3553.
[5] 秦玉川.昆虫行为学导论[M].北京:科学出版社,2008.
[6] 张清泉,张雪丽,陆 温,等.昆虫交配行为、繁殖适度和性信息素在国内的研究进展[J].广西农业科学,2009,40(2):164-168.
[7] 刘吉敏,黄其椿,檀志全,等.龙眼树荔枝蝽越冬成虫发生规律及交配节律[J].南方农业学报,2012,43(8):1135-1138.
[8] 孙丽娟,戴华国,衣维贤,等.二化螟水稻类群与菱白类群成虫羽化节律和交配节律研究[J].昆虫知识,2002,39(6):421-423.
[9] 徐德良.茶树蜡蝉种群生态及控制技术研究[D].苏州:苏州大学,2009.
[10] 陈彩贤.三斑广翅蜡蝉生活习性观察[J].广西农业科学,1998(3):139-141.
[11] 邓晗嵩.同翅目蛾蜡蝉科(f-tomopter:flatidae)生活习性及防治研究[J].黔西南民族师范高等专科学校学报,2007(4):122-124.
[12] 董文玲.白蛾蜡蝉生物学特性及防治研究[J].林业调查规划,2006,31(A02):159-161.
[13] 李慧玲,刘丰静,王定锋,等.白蛾蜡蝉的发生和防治[J].茶叶科学技术,2012(4):24-25.
[14] 何 娟,于玮台,陈文龙.贵阳地区茶树碧蛾蜡蝉的发生规律及防治[J].贵州农业科学,2013,41(7):105-107.
[15] 刘永生,胡 波,张清良,等.八点广翅蜡蝉生物学特性与防治初报[J].湖北林业科技,1999(2):29-30.
[16] 刘永生,张清良.柿广翅蜡蝉生物学特性及防治初报[J].山西果树,2000(4):5-6.
[17] 李苏萍,陈秀龙,韩国柱,等.山东广翅蜡蝉生物学特性及防治措施[J].中国森林病虫,2006,25(3):36-38.
[18] 罗晓明,罗天相,刘 莎.柿广翅蜡蝉的发生与防治[J].河南农业科学,2014(3):41-42.
[19] 刘朝萍.柑橘白蛾蜡蝉的发生与防治技术[J].植物医生,2009,22(5):25-25.
[20] 梅志坚.茶树碧蛾蜡蝉的发生与防治[J].茶叶科学技术,2004(3):43-43.
[21] 沈 强,王菊英,柳建定,等.山东广翅蜡蝉的生物学特性及防治[J].昆虫知识,2007,44(1):116-118.
[22] 江伟林.山东广翅蜡蝉在莲都地区的发生及综合防治[J].浙江柑橘,2013(1):35-36.
[23] 汪荣灶,祝 婕.茶广翅蜡蝉生物学初步研究[J].中国茶叶,2010(5):29.
[24] 于玮台,陈文龙.碧蛾蜡蝉行为习性研究[J].山地农业生物学报,2013,32(2):123-127.
[25] 喻爱林,单继红,涂业苟,等.油茶高产无性系碧蛾蜡蝉的生物学特性及防治[J].江西植保,2006,29(4):181-182.
[26] 喻爱林.油茶八点广翅蜡蝉的生物学特性及防治[J].江西林业科技,2007(3):34-35.
[27] 赵丰华,吕立哲,任红楼.信阳茶树新害虫——蜡蝉[J].中国茶叶,2010(10):16-17.
[28] 肖铁光,周社文,王文艺,等.V脊菱蜡蝉生物学特性和消长规律初步研究[J].湖南农业大学学报:自然科学版,2006,32(1):70-72.
[29] 赵丰华,吕立哲,任红楼,等.豫南茶园柿广翅蜡蝉生物学特征[J].中国茶叶,2011(5):18-19.
[30] 顾昌华.白斑广翅蜡蝉的生物学特性[J].贵州农业科学,1997,25(5):50-51.
[31] 柴正群,杨发军,朱建青,等.云南普洱茶树碧蛾蜡蝉的危害与防治[J].现代园艺,2012(21):64-64.
[32] 陈庆红.柿广翅蜡蝉生物学特性及其防治初步研究结果[J].湖北植保,2000(3):24-25.
[33] 谭万忠,李 培,张克诚,等.重庆地区茶树病害种类及武夷菌素对茶病的田间控制效果[J].西南大学学报:自然科学版,2008,30(4):123-127.
[34] 邵元海,熊 辉.可可广翅蜡蝉的生物学特性及防治[J].蚕桑茶叶通讯,2009(5):38-39.
[35] 邵元海,周静峰,徐德良.可可广翅蜡蝉若虫空间分布型及抽样技术研究[J].茶叶,2009,35(4):219-220,223.
(责任编辑: 王 海)
Behavior Rhythm ofRicaniacacaonis
FAN Guangyu, LIU Ningguo, YANG Qun, LIU Xuemin, CHEN Wenlong*
(InstituteofEntomology,GuizhouKeyLaboratoryforPlantPestManagementofMountainousRegion,GuizhouUniversity,Guiyang,Guizhou550025,China)
The behavior and rhythm of emergence, mating and oviposition ofR.cacaoniswere observed by indoor raising method and the oviposition feature in field was investigated to further complete forecasting and control ofR.cacaoniswell. Results: The emergence ofR.cacaonisadults occurs at night and before dawn. The mating behavior ofR.cacaonisadults happens in the day mainly.R.cacaonisadults lay eggs whole day and the peak time concentrates in the morning and before dawn.R.cacaonisadults mainly lay eggs in young stems with 1.5 mm diameter in field and the oviposition depth is about 0.9 mm. The oviposition scars mainly concentrate in 3 cm range to top shoot and the average eggs per scar is about 11.
Ricaniacacaonis; emergence; mating; oviposition; behavior rhythms
2015-09-08; 2015-12-30修回
贵州省科技厅农业攻关项目“茶树重要病虫害无公害防除技术研究与推广”[黔科合字NY字(2010)3024]
范广玉(1987-),男,在读硕士。研究方向:动物生态及害虫综合治理。E-mail:383441879@qq.com
*通讯作者:陈文龙 (1965-),男,教授,硕士生导师,从事植物病虫害研究。E-mail:CWL001@163.com
1001-3601(2016)04-0158-0053-04
S433
A
