贵州濒危树种岩生红豆森林群落特征及种群结构*
2016-02-26王茂师袁丛军安明态严令斌李鹤余德会
王茂师,袁丛军,安明态,严令斌,李鹤,余德会
(1.贵州林业勘察设计有限公司,贵州 贵阳550003;2.贵州大学林学院 贵州 贵阳550025;
3.贵州省林业科学研究院,贵州 贵阳550005;4.贵州雷公山国家级自然保护区管理局,贵州 雷山557100)
贵州濒危树种岩生红豆森林群落特征及种群结构*
王茂师1,袁丛军2,3,安明态2,严令斌2,李鹤2,3,余德会4
(1.贵州林业勘察设计有限公司,贵州贵阳550003;2.贵州大学林学院 贵州贵阳550025;
3.贵州省林业科学研究院,贵州贵阳550005;4.贵州雷公山国家级自然保护区管理局,贵州雷山557100)
摘要:通过典型群落样地调查,对不同干扰强度下岩生红豆所处森林群落特征及种群结构进行了研究,结果表明,(1)该种群所处群落乔木层物种较为丰富,多样性指数(SW)为2.66~3.26,生态优势度(ED)为0.11~0.17,各群落样地之间差异不显著;(2)岩生红豆能成为局部群落建群层的次优种或共优种,但数量很少,在整个群落乔木层处于明显弱势;(3)不同干扰强度下岩生红豆种群大小级指数不同,笔架山群落种群大小级指数(SDI)为负值(-0.003 7),种群连续;黔灵山的群落种群大小级指数(SDI)为正值(0.001 6),种群不连续;(4)群落更新层岩生红豆幼苗、幼树的年龄结构呈倒金字塔型,Ⅰ级幼苗数量较少,多为Ⅱ~Ⅳ级幼树,且数量稀少,天然更新明显不良。
关键词:不同干扰强度;物种多样性;种群结构;更新;岩生红豆
植物群落由不同植物种类组合而成,种群种类、大小及其结构组成特征直接影响着群落结构特征,亦是群落动态变化(发展趋势)的客观体现[1]。植物群落得以在时间上的不断延续、演替发展,空间上得以扩散拓展归结于植物种群的自然更新,其作为自然生态系统中植物资源再生产的一个重要生物学过程,是维持群落健康、稳定的一个重要因素[2]。伴随着人类生态足迹的不断扩增,适合植物的生境不断破碎化,珍稀濒危植物的保育成本与难度增大。此外,许多珍稀濒危植物由于自身种群数量、大小及结构与分布不合理,导致其在现有植物群落中的竞争能力差,并制约着种群的发展壮大[3]。自提出更新生态位的概念以来,种子繁殖和实生幼苗更新生态学研究得到广泛关注[4~6]。
植物更新繁殖包括有性种子繁殖和无性萌蘖繁殖两种方式,物种的更新策略取决于其自身生物学特性与环境压力权衡的结果[5~6]。天然更新能力取决于更新库的大小以及有利于更新库中植物个体生长发育的环境条件[7]。近年来,国内外的许多生态学者对植物天然更新做了大量研究工作,并取得了许多有利的研究成果[8]。然而多数研究工作均是从种子的更新方式、林窗更新理论来进行,而对植物萌生更新方式探讨研究较少[6]。
植物种群自然更新过程对生态因子的响应是植物生长生境适宜度的集中体现,往往受多个因子的综合作用[6]。已有研究表明非生物因子土壤中有效N是幼苗生长的限制因子之一,林下灌木层、草本层以及林分郁闭度对乔木幼苗存活、生长均具有显著的影响[9~12],尤其是人为破坏严重的具有珍稀濒危植物组成的植物群落。因此,开展以零星分布为主,不同人为干扰强度下的濒危树种岩生红豆森林群落特征及种群结构研究,对阐明该濒危种的物种生物生态特性、种群更新对策、群落稳定性及其演替规律等都具有重要意义。
岩生红豆(Ormosiasaxatilis)隶属豆科(Leguminosae)红豆属(Ormosia)常绿植物,为高大乔木,树皮灰绿色,是1981年发表的新种[13]。其树干通直,树形美观,是优良园林绿化树种和珍贵用材树种,具有很高的观赏价值和经济价值,由于数量稀少,已列入贵州省省级重点保护树种。现存植株多为伐桩萌蘖幼苗幼树,成熟个体已不足100株,且生境破坏仍在进行,资源数量仍在下降,该种已达极危等级,资源保护与拯救迫在眉睫[14]。然而有关岩生红豆种群研究工作较少,仅进行引种试验[15]、群落物种多样性对不同干扰的响应研究[16]以及群落主要树种生态位研究工作[14]。这在一定程度上揭示了该种群资源特征与现状,然而针对天然分布的岩生红豆所处森林群落特征及种群结构研究工作报道较少。本项目对岩生红豆天然群落的群落结构、幼苗幼树更新等特征进行研究,旨在为濒危树种岩生红豆进一步开展濒危机制和群落演替的研究以及天然林的保护提供参考。
1研究区概况
选择贵州省目前已知岩生红豆分布数量最多、最集中的贵阳市黔灵山和黔西南望谟县笔架山这两个分布区为主要调查研究区,其他分布点(多为散生个体)作辅助性调查。其中:(1)黔灵山位于贵阳市中心区西北,海拔1 100~1 300 m,为典型的喀斯特地貌,地形起伏明显。由于特殊的地理位置和地形地貌,该区冬无严寒、夏无酷暑、热量充沛、生长期长,年均降雨量约1 129.5 mm,年均温15.3℃,无霜期270天。由于毗邻市区,森林旅游发达,猕猴活动十分频繁,森林群落受到很大干扰;群落乔木层胸径16.4 cm、树高11.0 m,以岩生红豆、云贵鹅耳栎(Carpinuspubescens)、朴树(Celtissinensis)等为主。(2)望谟笔架山亦为典型的喀斯特地貌,地势西北高东南低,海拔1 080~1 200 m。该区属亚热带温湿季风气候,具有明显的春早、夏长、秋晚、冬短的特点。年平均气温为19℃,年均降水量1 222.5 mm,无霜期339天,冬无严寒,夏无酷暑,雨热同季。该研究区森林群落受到人类活动干扰较小,现有群落乔木层胸径10.1 cm、树高6.9 m,以青冈栎(Cyclobalanopsisglauca)、岩樟(Cinnamomumsaxatile)、圆果化香(Platycaryalongipes)等为主;灌木层以齿叶黄皮(Clausenadunniana)、密花树(Myrsineseguinii)、岩生红豆等为主;草本层以长茎沿阶草(Ophiopogonchingii)、苔草(Carexsp.)、芒(Miscanthussinensis)等为主[16]。
2研究方法
2.1 样地设置方法
根据研究区岩生红豆的分布状况,选择典型地段设置群落样地(表1),每块样地进行GPS定位。共设置样地5块(由于岩生红豆野生资源少,分布面积小),其中黔灵山2块,望谟笔架山3块,样地面积为20 m×20 m(其中,乔木层划分为16个5 m×5 m样方;灌木层和草本层分别划分为10个2 m×2 m和1 m×1 m的小样方),并详细记录样地内每株岩生红豆(包括幼苗、幼树)的株数、胸径(幼苗、幼树的基径)、冠幅、枝下高等指标,记录所处生境(包括海拔、坡度、土壤和岩石类型、生长状况、群落类型)及分布、自然更新与受破坏情况,同时记录各样方内伴生树种及其株数、胸(地)径、树高、冠幅等。通过对岩生红豆的幼苗或幼树根系进行适当刨根观察,记录其根系主要特点,判断更新苗是实生苗、根蘖苗或桩蘖苗。同时调查离伐桩或枯桩不同距离范围内根蘖苗的数量、地径和植株高(cm)。
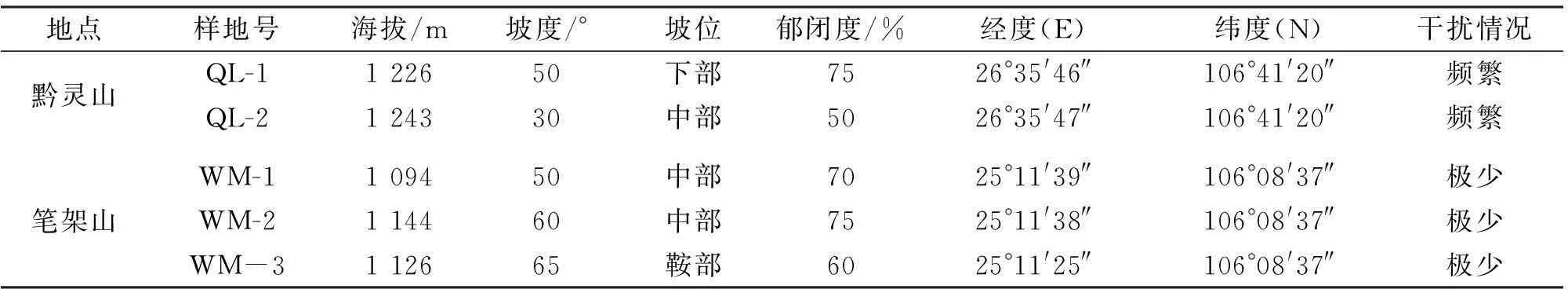
表1 样地调查基本情况
2.2 数据处理
2.2.1群落层次结构划分
基于生活型和森林群落实际特征,群落中起着控制和支配作用的群落层次归为群落建群层,即为乔木树种种类和林木直径≥5.0 cm的乔木树种总和。更新层即为群落中乔木树种林木直径<5.0 cm的所有林木总和。
2.2.2物种多样性测定
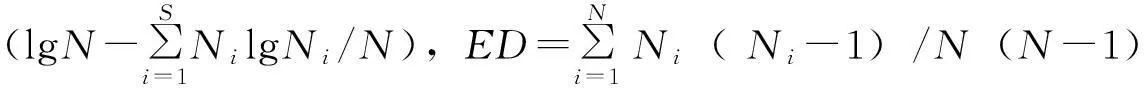
2.2.3重要值
对样地内的目的树种及灌木层的重要值进行分析,计算公式为[18],重要值=相对频度+相对密度+相对显著度;相对重要值=重要值/3。
2.2.4优势种数目确定
2.2.5径级结构划分
以个体高度(苗期)及径级代替龄级来反映岩生红豆的种群结构[20~21]。其中,幼苗(高度H<40 cm)、小幼树(40 cm≤ H <100 cm)和中幼树(100 cm≤ H <200 cm)3个级别的径级分别为Ⅰ、Ⅱ、Ⅲ级,H≥200 cm,但胸径(DBH)<5 cm的大幼树为Ⅳ级。以后以胸径(DBH)5 cm为起点,胸径每增加5 cm新增一个龄级,依次编号为Ⅴ、Ⅵ、Ⅶ、……。
2.2.6大小级指数
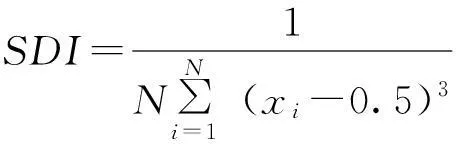
3结果与分析
3.1 所处群落建群层的物种多样性特征
研究区岩生红豆群落建群层(乔木层)物种丰富度、SW多样性指数较高而生态优势度较低(表2),森林群落之间物种多样性差异不明显,多样性指数(SW)波动不大,在2.66~3.26之间,生态优势度(ED)在0.11~0.17之间,说明岩生红豆所处的自然群落在组成上趋于多个树种组成的混交林。
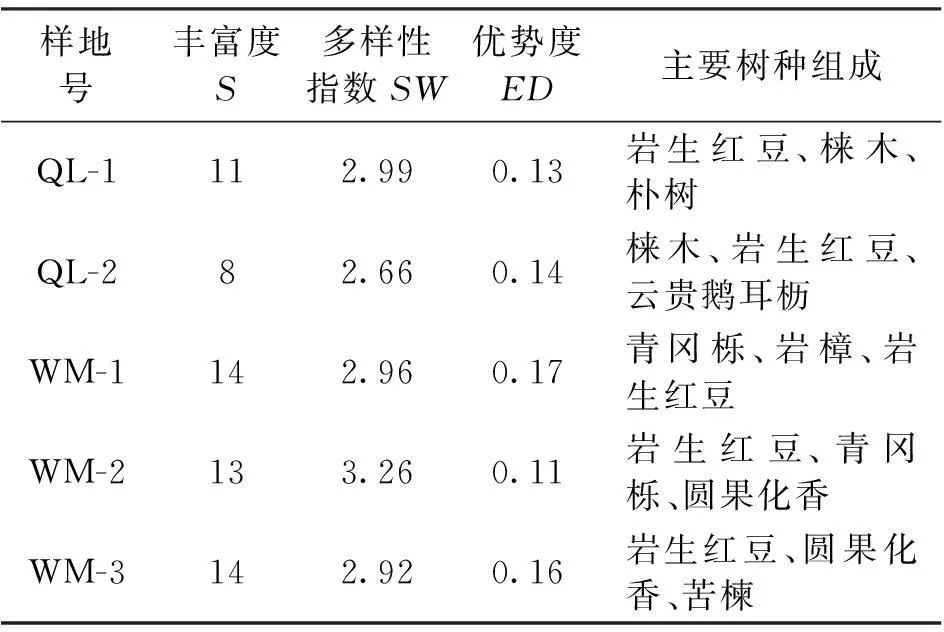
表2 群落建群层物种多样性特征

表3 岩生红豆天然林乔木层(建群层)主要树种的相对重要值
3.2 岩生红豆在群落中的地位分析
3.2.1在所处群落建群层的地位
岩生红豆在贵阳黔灵山和望谟笔架山这两个研究区的生长都能达到所处群落的建群层,在5个群落样地的建群层能成为次优种或共优种(表3)。岩生红豆在QL-1、QL-2、WM-3群落中重要值居第2位,在WM-1、WM-2群落中重要值居第3位,表明该种在研究区具有一定的竞争优势。
3.2.2在所处群落更新层的地位
各群落样地更新层皆不发达,其物种丰富度以及岩生红豆所处的地位差异亦很大(表4)。其中,黔灵山2个样地更新层不发达,且物种单一,OL-1仅有云贵鹅耳枥和岩生红豆共2个种,岩生红豆的相对重要值为0.43,居第二,且数量稀少;OL-2仅有云贵鹅耳枥1个种,缺岩生红豆幼苗和幼树。
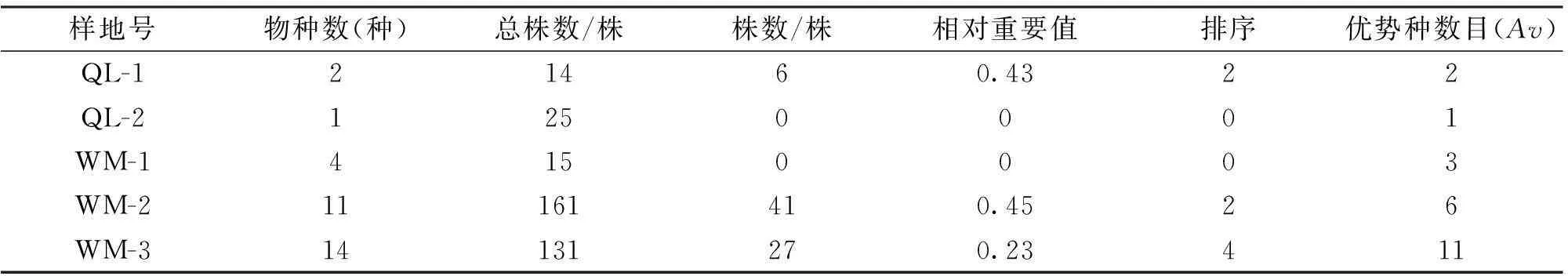
表4 岩生红豆在不同样地灌木层(更新层)所处的地位
3.3 种群径级结构
种群幼龄个体数量是种群更新的基础,不同起源对种群遗传多样性维持意义重大。通过统计贵阳黔灵山和望谟笔架山不同干扰条件下岩生红豆幼苗(Ⅰ级)、小幼树(Ⅱ级)、中幼树(Ⅲ级)、大幼树(Ⅳ级)的数量状况及各阶段岩生红豆的资源状况,发现两个调查点的岩生红豆更新层个体的数量结构均成倒三角分布,其中,幼苗(Ⅰ级)数量仅占8 %和15 %(图1)。
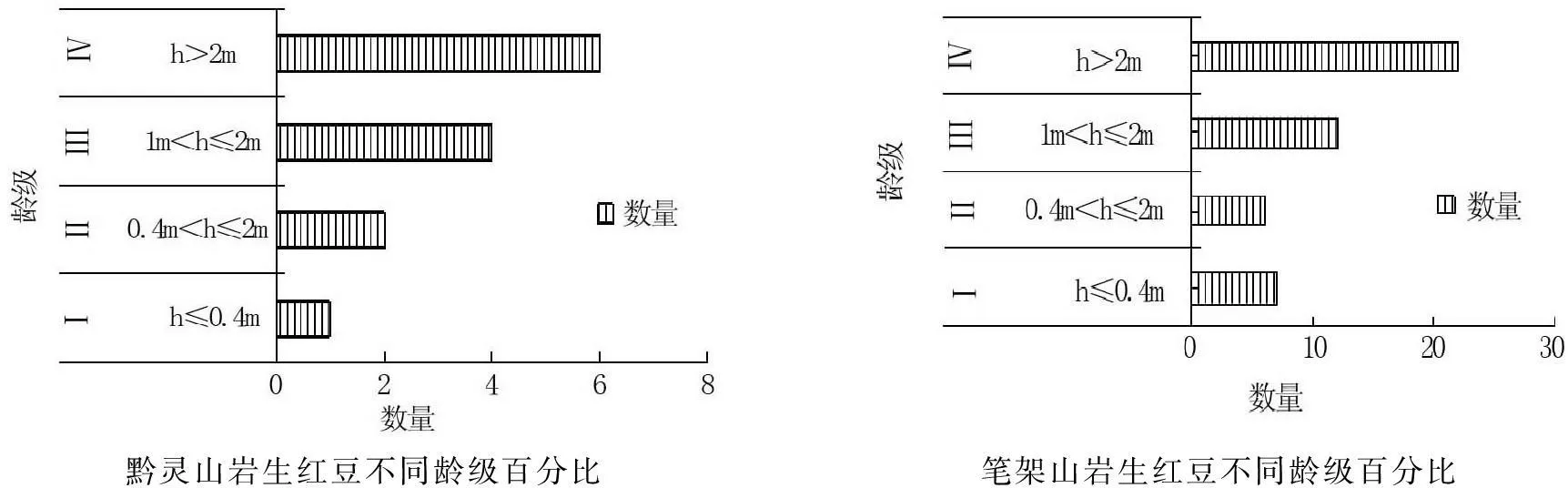
黔灵山岩生红豆不同龄级百分比笔架山岩生红豆不同龄级百分比
图1不同分布区(干扰强度)岩生红豆年龄级百分比
Fig.1Age class ofOrmosiasaxatilispopulation in different distribution areas (disturbance intensity)
3.4 大小级指数
林下干扰较大的黔灵山岩生红豆种群的SDI值为正值(表5),表明在该生境中岩生红豆的更新不连续,小径级个体数量稀缺,种群更新呈现严重的衰退趋势。而林下干扰较少的望谟笔架山岩生红豆种群的SDI为负值(表5),表明岩生红豆种群在该生境连续,小径级数量相对较多,林下更新相对较为正常。但即使在笔架山,岩生红豆种群的总体结构仍旧不合理(图1)。

表5 不同分布区岩生红豆SDI值
注:SDI表示大小级指数。
3.5 岩生红豆的更新方式及自然更新能力
在望谟笔架山和贵阳黔灵山研究区内皆未发现岩生红豆的种子繁殖的实生苗,只发现桩蘖和根蘖两种更新方式的苗木。从研究区情况看,岩生红豆具有一定的萌生能力,在研究区所有群落中都有不同程度的发生,每个树桩萌蘖少则1~2枝,较大树桩可达8~10株。但桩蘖仅发生在被砍伐或折断的树桩上,需要有活的残桩,这种更新方式是一种建立在破坏基础上的更新,且更新位置只有发生在原有植株生长点,不利于种群的扩散与健康发展。
以望谟笔架山研究区1株地径60 cm的残存岩生红豆树桩为例,该树桩被砍除多年,除残桩上有少数萌条外,在其周围裸露岩石上的根亦有萌条(图 2)。根蘖苗主要集中分布在离伐桩2~4 m的范围内,约占总根蘖苗数的49 %,在4 m以内根蘖苗质量无明显差异。4 m以外的根蘖苗较少,仅占总株数的36.8 %,产生根蘖苗的最远距离为11.5 m。
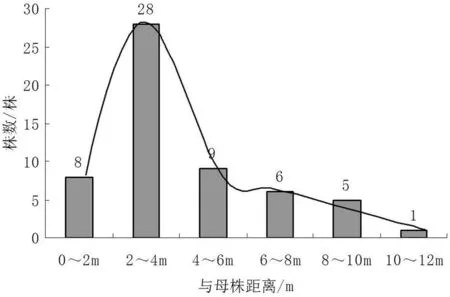
图2 岩生红豆离伐桩距离根蘖苗数量
4结论与讨论
4.1 结论
岩生红豆所处群落的建群层物种具有较高的丰富度和多样性,在各样地中物种数为8~14种,多样性指数(SW)为2.66~3.26;其生态优势度(ED)较低,为0.11~0.17。各群落之间物种丰富度、多样性与生态优势度的差异性不明显;岩生红豆所处群落更新层均不发达,实生苗缺乏,萌生苗数量稀少。
不同干扰强度下的岩生红豆种群均处于衰退趋势,黔灵山岩生红豆种群已出现严重的更新不连续现象;笔架山种群更新相对较为连续,但种群结构不合理,不利于种群结构的健康发展。
岩生红豆根蘖苗株数随着距离伐桩母株的增加呈先增加后减小趋势,以分布2~4 m数量最大,比例达49 %。
4.2 讨论
岩生红豆为贵州喀斯特山地适生特有珍贵乡土树种、贵州省省级重点保护树种,该种已达极危等级[14],资源保护与拯救相关研究工作开展已迫在眉睫。贵阳黔灵山、望谟笔架山是岩生红豆分布最为典型、资源数量相对较大的分布区域,两地因各自所处群落不同所受干扰情况不同,群落结构亦表现明显差异。两地岩生红豆所处群落更新层均不发达,尤其贵阳黔灵山群落更新层仅有1~2种,其物种丰富度显著低于群落建群层;望谟笔架山群落更新层亦只有4~14种,且个体数量很少,不利于整个群落的更新演替。研究结果与相关文献一致[16]。
良好、适宜的生态环境是珍稀濒危植物赖以生存的根本和基础[21]。人为干扰常被认为是珍稀濒危树种致危因子中最重要的外在因素。结合课题组对贵州特有种岩生红豆资源特征调查结果显示,岩生红豆常因其材质优良,成年树木多遭砍伐,这在其主要分布点贵州省的望谟、修文、惠水、独山、三都、长顺、花溪、清镇、息烽等(未处于保护区内或缺乏有效保护)皆十分严重,加之其分布多见于临时性干旱频繁的岩山环境,土壤瘠薄,生境恶劣,难成大树。即使在不会遭砍伐的贵阳黔灵山(黔灵公园内),由于旅游的影响,尤其引入大量的猕猴频繁攀爬,且喜啃食嫩芽,对幼苗幼树的影响很大。而且,自从黔灵山有了猕猴后,已多年没有看到岩生红豆开花结实。本研究对两种干扰强度下的岩生红豆所处群落分析表明,分布地的岩生红豆种群结构均不合理,甚至呈不连续状态(黔灵山),该种群更新困难已是客观事实。造成这一现状或更严重的情况根本原因与种群所处生境及本省生物生态学特性密切相关,而对其相关的深入研究尚未见报道。
植物根蘖更新是除种子繁殖更新的一种重要方式。植株通过母株根系上的不定芽产生无性繁殖的繁殖体,随后其定居成功后就成为潜在的独立个体(即无性系小株),达到种群更新的目的[23]。根蘖繁育更新作为岩生红豆野外天然群落中种群更新的一种重要方式,在维持自身种群数量、扩大及扩展扮演着极其重要的作用。研究表明,岩生红豆根蘖苗株数随着距离伐桩母株的增加呈先增加后减小趋势,以分布2~4 m数量最大,比例达49 %。据观察,在4 m以外,距伐桩越远,其根蘖苗顶端出现不同程度枯萎现象,生长的苗木质量较差,可能与根系状况及营养供应有关,具体原因有待进一步研究。
喀斯特山地适生乡土珍贵树种岩生红豆因其优良的材用价值、独特的观赏价值,人为不合理采伐破坏严重,资源分布极为零散,种群已处于极危状态。生境破碎化已逐渐成为限制岩生红豆种群更新和扩繁的主要原因,由于每一分布区中岩生红豆种群的消长都将影响到整个种群的稳定性,因而,当前迫切需要采取有效措施对每一分布区的现有种群给予就地保护,并适当进行人工引种繁育及回归保育。总之,深入研究岩生红豆种群的动态和周期性变化,加强保护与管理,对濒危物种岩生红豆的野生资源保护和种群恢复具有重要意义。
参考文献:
[1]达良俊,杨永川,宋永昌.浙江天童国家森林公园常绿阔叶林主要组成种的种群结构及更新类型[J].植物生态学报,2004,28(3):376-384.
[2]杨旭,杨志玲,雷虓,等.濒危植物凹叶厚朴幼苗更新及环境解释[J].林业科学,2013,49(12):36-42.
[3]王芳,喻理飞.喀斯特山地珍稀濒危植物掌叶木种群结构与分布格局研究[J].林业实用技术,2014(6):6-9.
[4]刘峰,陈伟烈,贺金生.神农架地区锐齿槲栎种群结构与更新的研究[J].植物生态学报,2000(4):396-401.
[5]陈沐,曹敏,林露湘.木本植物萌生更新研究进展[J].生态学杂志,2007(7):1114-1118.
[6]李小双,彭明春,党承林.植物自然更新研究进展[J].生态学杂志,2007(12):2081-2088.
[7]张传余,喻理飞,姬广梅.喀斯特地区不同演替阶段植物群落天然更新能力研究[J].贵州农业科学,2011(6):155-158.
[8]骆争荣,陈德良,杨辉,等.百山祖常绿阔叶林厚叶红淡比的萌蘖繁殖特性[J].生态学报,2015(16):1-10.
[9]Bungard R A,Zipperlen S A,Press M C,etal.The influence of nutrients on growth and photosynthesis of seedlings of two rainforest dipterocarp species[J].Functional Plant Biology,2002,29(4):505-515.
[10]Beckage B,Clark J S,Clinton B D,etal.A long-term study of tree seedling recruitment in southern Appalachian forests:the effects of canopy gaps and shrub understories[J].Canadian Journal of Forest Research,2000,30(10):1617-1631.
[11]Bruna E M.Effects of forest fragmentation onHeliconiaacuminataseedling recruitment in central Amazonia[J].Oecologia,2002,132(2):235-243.
[12]Howe H F,Smallwood J.Ecology of seed dispersal[J].Annual review of ecology and systematics,1982:201-228.
[13]蓝开敏.贵州省红豆树属一新种[J].南京林业大学学报(自然科学版),1981(1):134-135.
[14]袁丛军,喻理飞,安明态,等.岩生红豆天然林主要树种生态位研究[J].生态科学,2013(4):487-493+528.
[15]方小平,张廷忠,姚淑君,等.贵州省林科院树木园珍稀树种引种研究报告[J].贵州林业科技,1996(4):7-13.
[16]袁丛军,安明态,严令斌,等.两种干扰强度下岩生红豆天然群落特征及物种多样性分析[J].南方农业学报,2013(2):280-284
[17]王伯荪.植物群落学实验手册[M].广州:广东高等教育出版社,1996:36-65.
[18]王海明,李贤伟,陈治谏,等.四川喇叭河自然保护区珙桐群落特征与更新[J].山地学报,2005(3):360-366.
[19]覃林.统计生态学[M].北京:中国林业出版社,2009:81-97.
[20]王中磊,高贤明.锐齿槲栎林的天然更新——坚果、幼苗库和径级结构[J].生态学报,2005,25(5):986-993.
[21]苏维词.贵州喀斯特地区珍稀濒危植物及其保护[J].长江流域资源与环境,2002(2):111-116.
[23]Alpert P.Nutrient Sharing in Natural Clonal Fragments of Fragaria Chiloensis[J].Journal of Ecology,1996,84(3):395-406.
Community Characteristic and Population Structure of
Ormosia saxatilis in Guizhou Province
WANG Mao-shi1,YUAN Cong-jun2,3,AN Ming-tai2,YAN Ling-bin2,LI He2,3,YU De-hui4
(1.Forestry Survey Design Corporation of Guizhou Province,Guiyang Guizhou 550003,P.R.China;2.College of Forestry,Guizhou University,
Guiyang Guizhou 550025,P.R.China;3.Guizhou Academy of Forestry,Guiyang Guizhou 550005,P.R.China;
4.National Nature Reserve of Leigong Mountain,Leishan Guizhou 557100,P.R.China)
Abstract:Population structure and community characteristics ofOrmosiasaxatilisunder different disturbance densities was studies by using classical community sampling approach,and the results shows that:(1)The species diversity of tree layers in the community is rich.The diversity index (SW) is 2.66~3.26,the ecological dominance (ED) is 0.11~0.17,and there is no significant difference between different community samples ;(2)EvenOrmosiasaxatiliscould become subdominant species or co-dominant species of some local communities,but such phenomena is few and it is usually at a disadvantage position in the whole tree layer community;(3) The grade index ofOrmosiasaxatilispopulation size are different under different disturbance densities,theSDIin Bijia Mountain is a negative value (-0.003 7) with continuous population,andSDIin Qianling Mountain is positive (0.001 6) with discrete population;(4)The age structure of the regeneration layer in the communities is an inverted pyramid type.The number of I-grade seedlings is was small,and the most are saplings from II to IV,and natural regeneration ofOrmosiasaxatilisis not obvious.
Key words:different disturbance density;species diversity;population structure;natural regeneration;Ormosiasaxatilis
作者简介:第一王茂师(1988-),男,助理工程师,主要从事植物生态与林业资源保护管理研究。E-mail:1059296795@qq.com通讯 安明态(1975-),男,高级工程师,博士生,主要从事植物分类学与植物生态学研究。E-mail:gdanmingtai@126.com
基金项目:贵州省林业科学技术研究项目(2009-6),贵州省科技支撑项目(黔科合NY字[2010]3058号),贵州大学自然科学青年科研基金项目(贵大自青基合字2009-004)。
*收稿日期:2015-06-17
中图分类号:Q 948;S 718.54
文献标识码:A
文章编号:1672-8246(2016)01-0081-07

