不同能量水平饲粮对7~10月龄荷斯坦育成牛生长性能和血清指标的影响
2016-01-19曾书秦刁其玉王建芬王俊司丙文杨春涛屠中国农业科学院饲料研究所奶牛营养学北京重点实验室北京0008北京延庆县畜牧技术推广站北京000北京市畜牧总站北京0009
曾书秦刁其玉王建芬王 俊司丙文杨春涛屠 焰∗(.中国农业科学院饲料研究所,奶牛营养学北京重点实验室,北京0008;.北京延庆县畜牧技术推广站,北京000;.北京市畜牧总站,北京0009)
不同能量水平饲粮对7~10月龄荷斯坦育成牛生长性能和血清指标的影响
曾书秦1刁其玉1王建芬2王 俊3司丙文1杨春涛1屠 焰1∗
(1.中国农业科学院饲料研究所,奶牛营养学北京重点实验室,北京100081;2.北京延庆县畜牧技术推广站,北京102100;3.北京市畜牧总站,北京100029)
摘 要:本试验旨在研究不同能量水平饲粮对7~10月龄育成牛生长性能和血清指标的影响。选取(200±3)日龄中国荷斯坦育成母牛24头,随机分为A、B和C 3个处理,对应饲喂产奶净能分别为5.40、5.90和6.40 MJ/kg的试验饲粮,每个处理8个重复,每个重复1头牛。预试期10 d,正试期90 d。于210、240、270和300日龄清晨空腹测定生长性能指标(体重、体高、体斜长、胸围、腹围、腰角宽、乳头长度、体况评分),并采集血液样品用于血清指标(甘油三酯、尿素氮、雌激素、雌二醇、瘦素、生长激素和胰岛素样生长因子-1)分析。结果表明:1)在210~300日龄,A、B、C组育成牛平均日增重分别为0.69、0.81、0.96 kg/d,C组显著高于A组(P<0.05)。2)在240日龄,A组左后乳头长度均显著高于C组(P<0.05);240和270日龄,A、B组右后乳头长度显著高于C组(P<0.05)。3)在240日龄,A和B组血清甘油三酯(TG)含量显著低于C组(P<0.05)。4)在300日龄,A组血清雌二醇(E2)含量显著高于B组(P<0.05),极显著高于C组(P<0.01)。综合以上,在本试验条件下,饲粮产奶净能为5.90 MJ/kg时,能维持育成牛体型和乳腺的正常发育。
关键词:饲粮能量水平;育成牛;生长性能;血清指标;乳头发育
能量是动物生长发育的基础,动物的所有活动,如血液循环、生长发育、合成体内激素等都需要能量。7~10月龄育成牛是奶牛培育的重要阶段,该阶段营养水平的高低会直接影响其生长性能和乳腺组织的发育。饲粮营养成分含量过高,易导致乳腺组织脂肪的堆积,不利于乳腺发育[1];反之则会导致生长发育受阻,发情和配种受胎延迟,对其繁殖性能造成影响[2]。同时,饲粮能量水平也是控制动物采食量最重要的因素,饲粮能量水平发生变化时,动物通过调整采食量来维持相对稳定的能量摄入量。因此选择适宜能量水平饲粮对维持成年奶牛理想体型、繁殖性能和泌乳性能有重要意义[3-4]。陈福音[5]比较了不同能量水平的饲粮对3~5月龄荷斯坦母牛生产性能的影响,结果显示各处理牛的平均日增重(ADG)随饲粮能量水平的升高而提高;崔祥等[6]同样证实在干物质采食量(DMI)一致的情况下,生长母牛的ADG与饲粮能量水平呈正相关。乳腺组织的发育也与血清激素含量密切相关[7],Block等[8]研究了饲粮能量水平对3~12月龄后备牛血清激素含量的影响,其中血清胰岛素样生长因子-1(IGF⁃1)含量随饲粮能量水平升高而升高,生长激素(GH)含量随饲粮能量水平升高而降低,而其中血清激素含量的改变直接影响乳腺组织的发育。后备牛的培养目标是14月龄时,体重达到380~400 kg,体高120~130 cm。育成牛作为成年牛群的基础,该阶段的培育对其成年后的生产性能和繁殖性能至关重要,但其各阶段饲粮营养水平,特别是能量水平的研究与应用未能得到应有的重视。本试验通过研究不同能量水平饲粮对育成牛生长性能、乳腺发育以及血清指标的影响,探讨7~10月龄中国荷斯坦育成牛饲粮适宜的能量水平,为培育优质育成牛提供参考。
1 材料与方法
1.1 时间和地点
本试验在北京市顺义区望加奶牛养殖中心进行,时间为2013年10月至2014年4月。
1.2 试验动物与试验设计
本试验采用单因素随机分组设计。选用24头健康荷斯坦育成母牛,按日龄、体重相近原则随机分成3个处理,每个处理8个重复,每个重复1头。犊牛试验初始为(200±3)日龄,初始体重为(268.10±7.32)kg。参照文献[9]推荐的育成牛营养需要,按250 kg体重生长母牛预期ADG为900 g/d计算,配制产奶净能(NEL)为5.40、5.90 和6.40 MJ/kg(干物质基础)、粗蛋白质(CP)含量基本一致的3种全混合日粮(TMR)[10]。试验饲粮组成及营养水平见表1。
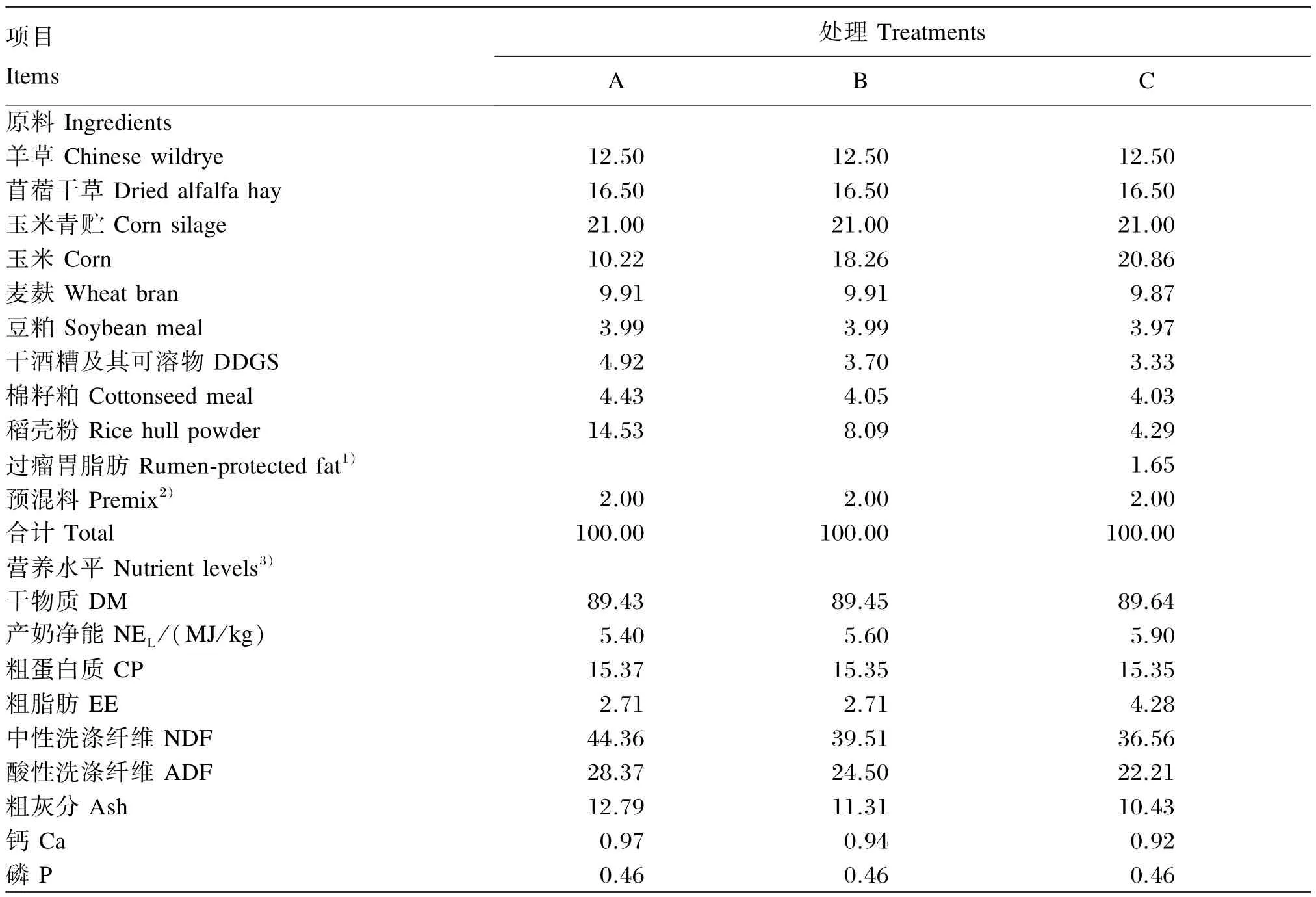
表1 试验饲粮组成及营养水平(干物质基础)Table 1 Composition and nutrient levels of experimental diets(DM basis) %
1.3 饲养管理
试验用育成牛按组分圈饲养,保证牛舍卫生。预试期10 d,预试期内每头牛按体重的2.6%(以干物质计)饲喂试验饲粮。正试期90 d。各头试验牛日饲喂量相同,按B组(饲粮NEL为5.90 MJ/kg组)平均体重的2.6%计算,单独记录每头牛的采食量。每10 d根据采食情况和体重变化进行调整。自由饮水。每天饲喂3次,第1次:06:00,第2次:12:00,第3次:18:00。放足量盐砖供试验牛自由采食。
1.4 测定指标与方法
1.4.1 饲粮营养水平
饲粮中粗蛋白质含量采用凯氏定氮法测定,粗脂肪含量采用石油醚反复抽提法测定,中性洗涤纤维和酸性洗涤纤维含量采用滤袋法测定,钙含量采用原子吸收法测定,磷含量采用钼酸铵分光光度计法测定,粗灰分含量采用灼烧法测定。
1.4.2 生长性能指标
在210、240、270、300日龄晨饲前测定每头牛的体重、体尺(体高、体斜长、胸围、腹围和腰角宽)、前、后乳头长度并进行体况评分(BCS)。根据各生长阶段始、末体重值计算ADG。详细记录每头牛每天的饲喂量和剩料量,每2周采集饲粮和剩料样品,混合后测定干物质含量,计算每头牛的DMI和料重比。乳头长度测定和体况评分分别参照Lammers等[11]和Wildman等[12]的方法。
1.4.3 血清指标
在210、240、270、300日龄晨饲前每处理以体重接近平均体重为原则选取5头牛,采用颈静脉真空管刺穿采血约20 mL,3 000 r/min离心25 min,收集血清于5 mL的离心管中,-20℃冷藏待测。血清样品采用全自动生化仪(日立7160,日本)和酶联免疫吸附测定法(ELISA)以试剂盒测定相关指标。测定指标包括生化指标:甘油三酯(TG)、尿素氮(UN)含量;血清激素及生长因子:雌激素、雌二醇(E2)、瘦素、GH和IGF⁃1含量。
1.5 统计分析
采用SAS 9.2统计软件中的单因素方差分析(one⁃way ANOVA)和关于重复测量数据的Mixed模型进行分析,并采用LSD法进行双重比较,统计模型如下:
Yijk=μ+Ti+Mj+TMij+C(T)ik+εijk。
式中:μ为总平均值;T为处理(i=1、2、3),固定效应;M为日龄(j=1、2、3、4),随机效应;C为犊牛(k=1、2……24),随机效应;ε为残差。
统计分析以P<0.05为差异显著,P<0.01为差异极显著。Mixed模型分析结果中,处理×日龄交互作用0.05≤P<0.10时,说明在不同日龄下各处理间的变化规律有存在差异的趋势,故列出各日龄阶段数值进行分析。
2 结果与分析
2.1 生长性能
2.1.1 增重
由表2可知,育成牛在210日龄初始的体重各组间无显著差异(P>0.05),平均值为268.07 kg;在240、270和300日龄,育成牛体重各组间差异不显著(P>0.05)。随着日龄增加,育成牛的体重出现极显著的变化(P<0.01),处理与日龄交互作用显著(P<0.05)。在210~300日龄,C组ADG显著高于A组(P<0.05),并且C组与A、B组相比,分别提高了39.13%和18.52%。本试验通过控制采食量,各组ADG控制在0.69~0.96 kg/d范围内。
2.1.2 DMI和料重比
由表2可知,育成牛采食精料、苜蓿干草、羊草、玉米青贮和合计DMI各组间均无显著差异(P>0.05),C组育成牛料重比得到改善,较A、B组分别降低了37.83%和19.24%。
2.1.3 体尺指标
由表3可知,在210~300日龄,育成牛体高、体斜长、胸围、腹围和腰角宽各组间均无显著差异(P>0.05);育成牛各体尺指标均随着日龄的增加极显著升高(P<0.01);对体高、胸围、腹围和腰角宽,处理与日龄间无显著交互作用(P>0.05),对体斜长,处理与日龄间交互作用显著(P<0.05)。
在210~300日龄,育成牛体况评分各组间无显著差异(P>0.05);育成牛体况评分随日龄的增长变化不显著(P>0.05);对体况评分,处理与日龄间的交互作用也不显著(P>0.05)。
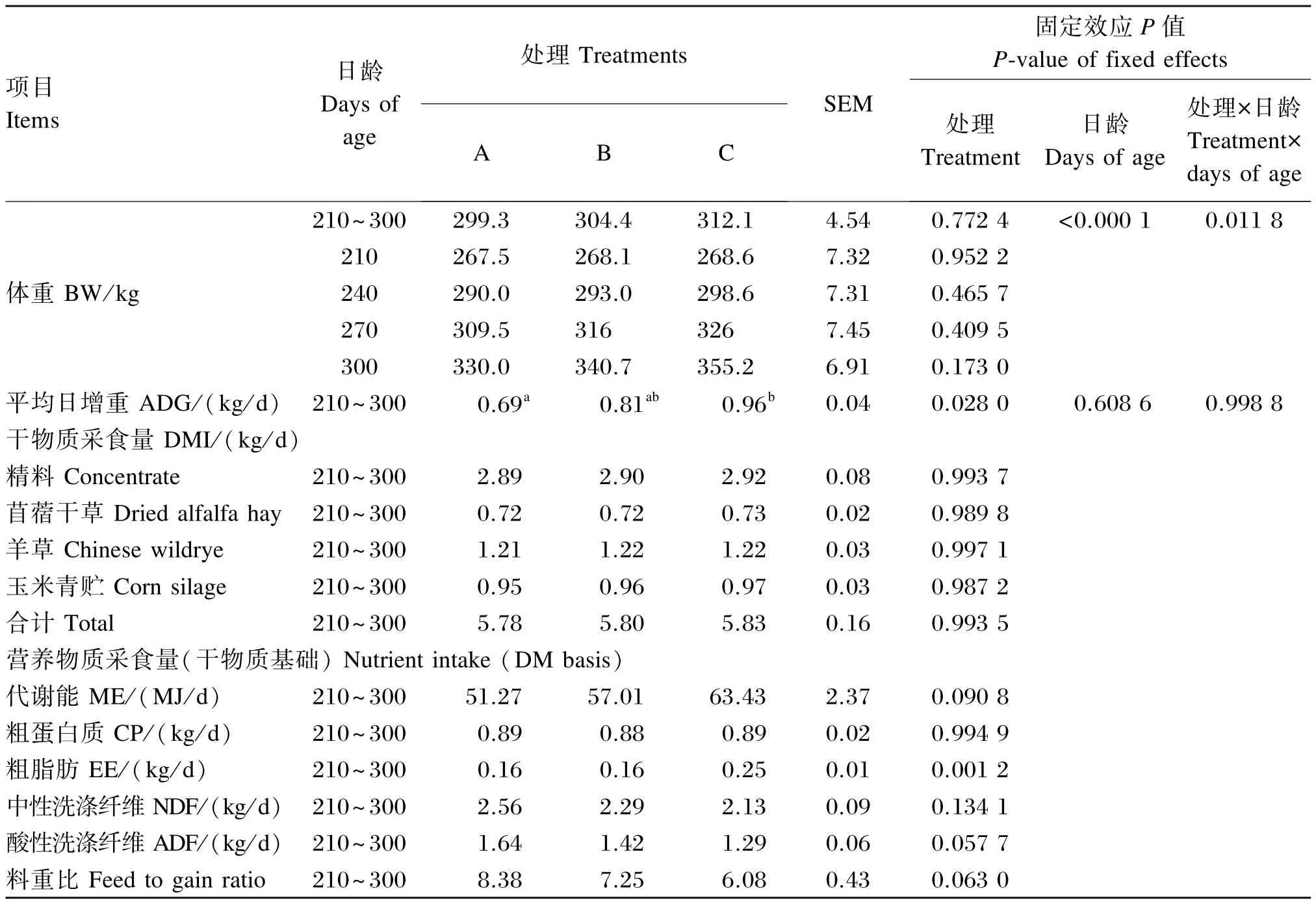
表2 不同能量水平饲粮对育成牛增重、干物质采食量和料重比的影响Table 2 Effects of diets with different energy levels on weight gain,dry matter intake and feed to gain ratio of heifers(n=8)
2.1.5 乳头发育
由表4可知,在210~300日龄,育成牛4个乳头长度受饲粮能量水平影响均不显著(P>0.05),前、后乳头长度分别在31.29~33.16 mm和28.04~30.31 mm之间变化,育成牛前乳头平均长度比后乳头平均长度长3.26 mm;育成牛乳头长度随日龄的增加极显著提高(P<0.01);处理与日龄间交互作用不显著(P>0.05)。在240日龄,A组左后乳头长度显著高于C组(P<0.05),且A组较B、C组分别增加了12.31%和14.15%;在240和270日龄,A、B组右后乳头长度显著高于C组(P<0.05),在240日龄,A、B组较C组分别增加了14.94%、5.75%;在270日龄,A、B组较C组分别增加了11.10%、10.37%。
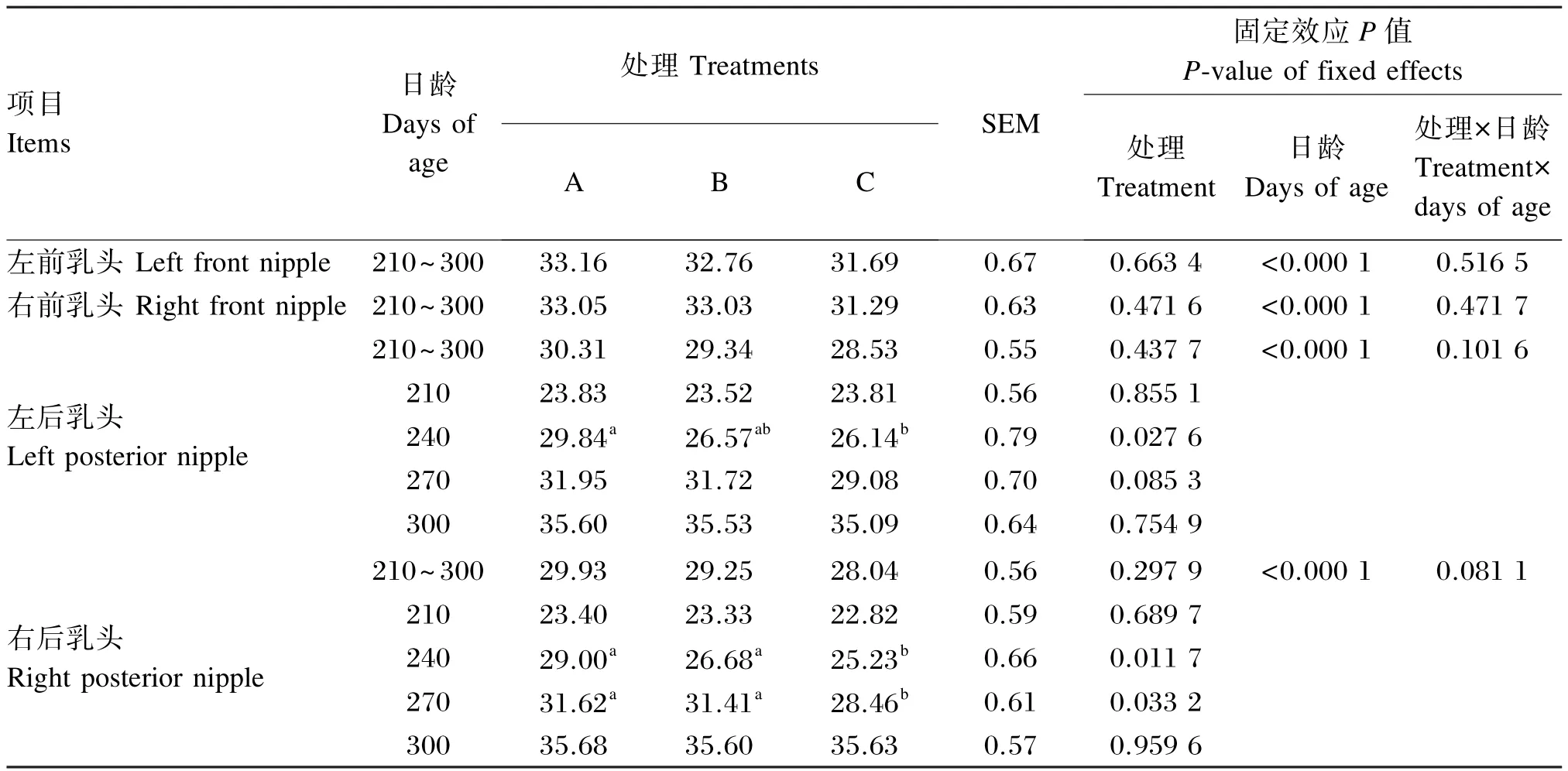
表4 不同能量水平的饲粮对育成牛乳头长度的影响Table 4 Effects of diets with different energy levels on nipple length of heifers(n=8) mm
2.2 血清指标
2.2.1 血清生化指标
由表5可知,在210~300日龄,育成牛血清TG含量各组间差异不显著(P>0.05),但在240日龄,C组血清TG含量显著高于A、B组(P<0.05)。在210~300日龄,育成牛血清UN含量各组间差异不显著(P>0.05);日龄对血清UN含量有极显著影响(P<0.01)。
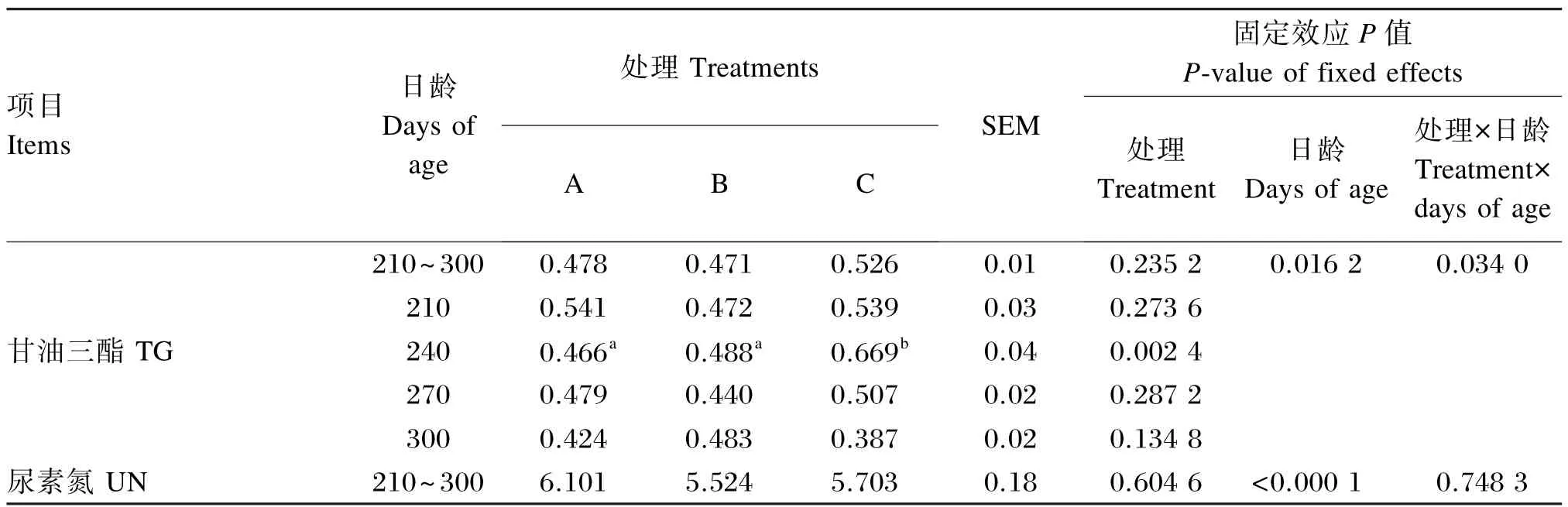
表5 不同能量水平的饲粮对育成牛血清生化指标的影响Table 5 Effects of diets with different energy levels on serum biochemical indices of heifers(n=5) mmol/L
2.2.2 血清激素及生长因子
由表6可知,在210~300日龄,育成牛血清E2含量差异不显著(P>0.05);在300日龄时,C组血清E2含量极显著低于A组(P<0.01),显著低于B组(P<0.05)。在210~300日龄,血清雌激素和瘦素含量各组间差异不显著(P>0.05),日龄对这2个指标有显著或极显著影响(P<0.05或P<0.01)。在210~300日龄,各组间育成牛血清GH 和IGF⁃1含量差异不显著(P>0.05),日龄对这2个指标无显著影响(P>0.05),处理与日龄间交互作用也不显著(P>0.05)。
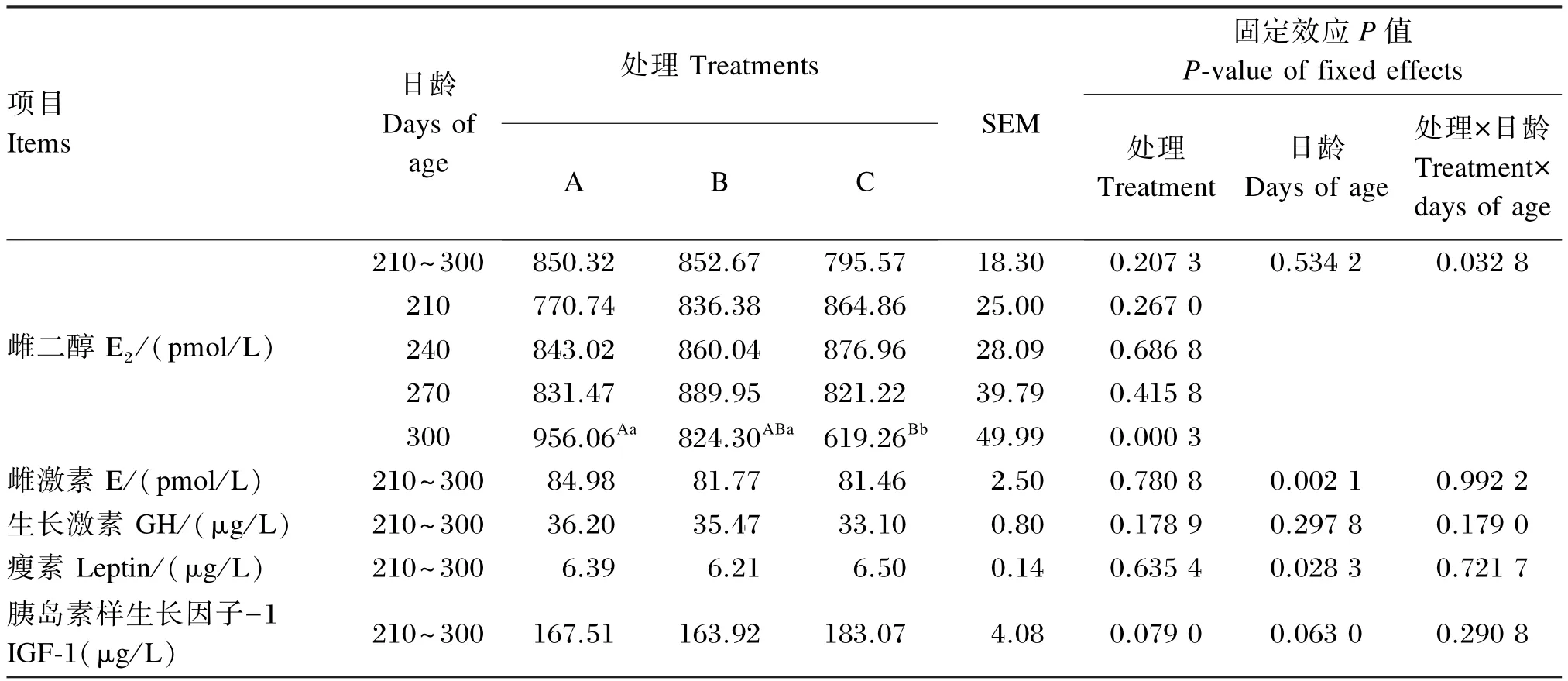
表6 不同能量水平饲粮对育成牛血清激素及生长因子含量的影响Table 6 Effects of diets with different energy levels on hormone and growth factor contents in serum of heifers(n=5)
3 讨 论
3.1 不同能量水平饲粮对育成牛生长性能的影响
奶牛从出生到成年,体重随着年龄的增长绝对量逐渐增长,这与体高、胸围和腹围这3项体尺指标的变化一致,各体尺的生长发育强度随年龄的增长发生转移,在6~14月龄,体重、胸围、体长和体高随年龄呈线性增长[13],本试验也证实了各体尺指标随日龄的增长而极显著增加。
体况评分是衡量奶牛体组织储存状况及监控奶牛能量平衡的一种方法,一般认为奶牛的理想体况是2.5~3.5之间,但不同生理期有一定差异[14]。本试验育成牛的体况评分在2.8~2.9范围内,表明其生长状况良好,未出现营养过剩或营养不良的情况。
崔祥等[6]研究表明,在限饲水平相同的条件下,犊牛NEL水平依次为6.24、7.04、7.53和7.85 MJ/kg,其ADG分别为0.64、0.75、0.78和0.84 kg/d,ADG随饲粮能量水平升高而增加,而料重比呈降低趋势。本试验采用的也是限饲方法,各组的采食量保持一致,育成牛合计DMI各组间差异不显著,保持在5.78~5.83 kg之间,但饲料NEL水平从A到C组依次升高,使得育成牛的NEL采食量依次提高,料重比得到改善,6.40 MJ/kg组较5.40和5.90 MJ/kg组分别降低了37.83%和19.24%,提高了试验牛的ADG。这同崔祥等[6]的研究结果是相一致的。本试验3个处理饲粮中玉米比例逐渐提高,而稻壳粉比例降低,玉米中淀粉含量较高,在瘤胃中降解为葡萄糖被瘤胃微生物摄取,后通过细胞酶的作用又可迅速降解为丙酸。葡萄糖代谢产生的烟酰胺腺嘌呤二核苷酸磷酸(NADP)是脂肪酸生物合成所必需的,丙酸是葡萄糖最重要的前体物。当丙酸的代谢不足以提供足够的还原型烟酰胺腺嘌呤二核苷酸磷酸(NADPH2),瘤胃发酵产生的大量乙酸就会转化为热,阻止更多的代谢。因此,饲料间的组合效应反映到组织代谢层次上,通过改变挥发性脂肪酸含量和比例以及葡萄糖的比例和数量,进而影响饲粮养分和能量的利用效率[15-16]。由于上述原因,本试验料重比随能量水平的升高而降低,说明了其主要受玉米中淀粉代谢的影响。
3.2 不同能量水平饲粮对育成牛血清生化指标的影响
TG是动物机体贮存能量的主要形式,是体内含量最多的脂类,可在肝脏和脂肪等组织中合成与贮存。本试验中高NEL(6.40 MJ/kg)饲粮添加了一定量的过瘤胃脂肪,采食高过瘤胃脂肪饲粮促使机体脂肪转运机制的活化,从而引起TG含量的上升[17],试验牛血清TG含量在240日龄时有所升高。
UN是反映蛋白质和氨基酸之间平衡的重要指标。在210~300日龄,各试验组间血清UN含量差异不显著,但数值上随着饲粮NEL水平的提高先降低后升高。饲粮中碳水化合物缺乏,或是瘤胃微生物生长繁殖受到抑制都会导致体内UN含量升高;瘤胃蛋白质降解减少,或是菌体蛋白合成量增加也会导致UN含量的降低[18-20]。本试验条件下,5.90 MJ/kg组血清UN含量相对另外2组偏低,有可能暗示适宜能量水平的饲粮可以改善育成牛的氮代谢,但这一问题尚需要进一步试验来证实。
3.3 饲粮能量水平对育成牛血清激素及生长因子含量的影响
产奶量是奶牛业生产的一项重要经济技术指标,产奶量有总产和单产之分,其中单产是衡量牛群品质、饲养管理水平和经济效益的主要依据。从市场需求和资源潜力方面考虑,奶牛业具有很大的发展空间,因此提高奶牛单产是转变奶牛业经济增长的方式、建立质量效益型奶牛业的必由之路,也是奶牛业面临的重大抉择。育成牛作为成年牛群的基础,其乳腺发育是影响成年后奶牛单产量最重要的因素,在奶牛3~9月龄阶段为异速生长阶段,在该阶段乳腺组织细胞的生长速度是体细胞的3~4倍,衡量乳腺发育的指标包括乳腺软组织的表观指标、乳腺实质和脂肪垫质量及DNA含量、血清内与乳腺发育相关的激素水平、乳腺发育相关基因的表达量及体外培养的分泌上皮细胞的增殖速率和数量等,其中脂肪垫质量、脂肪垫质量占整个乳腺质量的比例与ADG呈正相关[21]。奶牛的产奶性能主要由其乳腺合成细胞的数量来决定,这就意味着ADG与产奶性能之间存在负相关关系。当ADG过大,乳腺脂肪沉积过多,乳腺合成细胞数量减少,进而间接导致成年后的泌乳性能下降。Sejrsen等[22-24]认为生长母牛ADG为0.7~0.8 kg/d时,其成年后的产奶性能可达最大。也有学者认为,初情期前母牛的乳头长度能间接反映首次发情前乳腺导管的发育程度[12]。本试验中,210~300日龄饲粮NEL为5.40、5.90和6.40 MJ/kg时,ADG分别为0.69、0.81和0.96 kg/d,而各处理间前乳头长度的差异不显著,但在240和270日龄时,后乳头的长度随饲粮能量水平的变化产生了差异,这可能是受血清激素和ADG的双重的调控。因此可以推测,当饲粮NEL为5.90 MJ/kg时,既可以保持ADG在合理范围内,同时又能最大促进乳腺的发育。
E2是由卵巢成熟滤泡分泌的一种雌激素,能够促使乳腺导管发育增生,降低血液中胆固醇含量,并且能增加钙在骨中的沉积。雌激素在乳腺发育中起关键作用,可刺激乳腺导管生长,显著增加乳腺细胞的孕酮受体数目,而孕酮受体是促进乳腺小叶腺泡发育的基础。在育成牛营养因素对乳腺发育影响的研究中,血清E2和雌激素含量可作用衡量乳腺发育的重要的指标来指示乳腺导管的生长情况。崔祥等[6]研究表明,3~6月龄犊牛血清E2含量不受饲粮能量水平的影响[6],而本试验中,210~300日龄各处理间血清E2含量差异虽不显著,但在300日龄时,NEL为5.40和5.90 MJ/kg组育成牛血清E2含量显著或极显著高于NEL为6.40 MJ/kg组,这可能是由于300日龄时,育成牛处于初情期阶段,机体内激素的调控机制受饲粮营养水平的影响较大。雌激素相关受体α是能量代谢的调节者[25],本试验各处理血清雌激素含量随饲粮能量水平的升高而降低,可能是因为不同能量水平的饲粮影响雌激素相关受体α,进而间接影响雌激素含量的变化。
本试验条件下,血清GH含量随饲粮能量水平的升高逐步降低,而ADG随饲粮能量水平的升高呈增加的趋势。GH作为一种脑垂体激素,可控制动物生长及营养物质的代谢,并且与ADG有一定相关性[26]。随着动物体逐渐发育成熟,体内蛋白质合成量慢慢减少,脂肪沉积增加,而GH的作用是促进蛋白质的合成以及加速脂肪的水解,试验处理ADG越大,动物相对发育成熟些,因此GH的作用相对降低,含量也减少。前人研究认为乳腺发育也与血清GH含量呈正相关[27]。Meyer等[28]认为初情期前采食高能饲粮会降低血清GH含量。Hayden等[29]研究表明,阉犊牛营养限饲期间血浆GH的含量升高,但恢复营养后GH含量又降低。本试验设置了3个能量梯度,血清GH含量随能量水平的升高而有所降低,这可能是由于GH受下丘脑释放因子(GHRF)和促生长激素抑制素(SS)2种激素的调节,其分泌是脉冲式的,试验处理能量水平的差异引起下丘脑分泌的SS对垂体分泌的GH的负反馈作用加强有关,但该作用并未达到显著性水平。
研究表明,给3~6月龄荷斯坦生长母牛饲喂不同能量水平的饲粮,各处理乳头长度不受饲粮能量水平的影响,其血清E2和GH的含量各组间差异也不显著[6]。而本试验,各试验处理血清E2、雌激素和GH含量均随饲粮能量水平的升高而逐渐降低,可见随着奶牛的生长发育,其体内的激素达到一定水平,进而对机体组织的生长发育起到一定的促进作用,这也可能是在240和270日龄时,NEL为5.40、5.90 MJ/kg组的后乳头长度高于6.40 MJ/kg组的原因。
瘦素是脂肪细胞分泌的一种蛋白质,主要作用是调节能量平衡,促进动物初情期的启动,并且与乳腺的发育相关。研究发现羊和牛营养不良时,脂肪瘦素mRNA水平降低[30],Chelikani等[31]研究表明,3~12月龄生长母牛血浆瘦素含量随饲粮能量水平的升高而升高。本试验血清瘦素含量在NEL为5.40、5.90和6.40 MJ/kg组间依次升高,代谢能为6.40 MJ/kg组血清瘦素含量高于另外2组,但差异并不显著,而NEL为5.90 MJ/kg组血清中含量低于5.40 MJ/kg组,这可能是因为奶牛血浆及脂肪组织瘦素基因表达不仅与饲粮能量有密切联系,而且与饲粮中粗蛋白质、粗纤维、钙及磷等含量有一定关系[32]。
前人研究表明,饲粮能量水平显著影响血清IGF⁃1的含量[33]。本试验中也发现6.40 MJ/kg组血清IGF⁃1的含量高于另外2组。胰岛素样生长因子(IGF)是由肝脏分泌的一种多功能性细胞增殖调控因子,通过受体介导作用于靶组织,发挥生物学功能。IGF的合成与分泌受GH的调节和机体营养状态的影响,并且GH与IGF⁃1在机体内形成一个GH⁃IGF的轴,共同调控机体大多数组织生长[34]。高能饲粮饲喂后,打破了GH⁃IGF轴,导致6.40 MJ/kg组血清GH含量下降,IGF⁃1含量上升。
4 结 论
本试验条件下,给7~10月龄荷斯坦育成牛供给NEL分别为5.40、5.90和6.40 MJ/kg,粗蛋白质含量基本一致的饲粮,得出如下结论:
①随着饲粮能量水平的提高,育成牛ADG显著升高,NEL为5.90和6.40 MJ/kg时ADG分别达到了0.81和0.96 kg/d。
②较高饲粮能量水平可降低血清E2含量,影响育成牛乳头长度,饲喂NEL为6.40 MJ/kg的饲粮,育成牛270日龄的右后乳头长度显著低于饲喂其他2种饲粮。
③240日龄,饲喂NEL为6.40 MJ/kg的饲粮,育成牛血清TG含量显著高于饲喂其他2种饲粮。
综上,饲粮NEL为5.90 MJ/kg时,既可维持育成牛800 g/d的ADG,又能较有效地促进乳腺的发育。
参考文献:
[1] RADCLIFF R P,VANDEHAAR M J,SKIDMORE A L,et al.Effects of diet and bovine somatotropin on heifer growth and mammary development[J].Journal of Dairy Science,1997,80(9):1996-2003.
[2] 王根林.养牛学[M].北京:中国农业出版社,2006:290.
[3] COLE D J A,DUCKWORTH E,HOLMES W,et al.Factors affecting voluntary feed intake in pigs.3.The effect of a period of feed restriction,nutrient density of the diet and sex on intake,performance and carcass characteristics[J].Animal Production,1968,10(4):345-357.
[4] CHIBA L I,LEWIS A J,PEO E R,Jr.Amino acid and energy interrelationships in pigs weighing 20 to 50 kil⁃ograms:Ⅰ.Rate and efficiency of weight gain[J].Journal of Animal Science,1991,69(2):694-707.
[5] 陈福音.育成奶牛能量和蛋白质代谢规律及其需要量研究[D].硕士学位论文.保定:河北农业大学,2012.
[6] 崔祥,刁其玉,张乃锋,等.不同能量水平的饲粮对3~6月龄犊牛生长发育及血清指标的影响[J].动物营养学报,2014,26(4):947-961.
[7] PLAUT K,IKEDA M,VONDERHAAR B K.Role of growth hormone and insulin⁃like growth factor⁃Ⅰin mammary development[J].Endocrinology,1993,133(4):1843-1848.
[8] BLOCK S S,SMITH J M,EHRHARDT R A,et al.Nutritional and developmental regulation of plasma leptin in dairy cattle[J].Journal of Dairy Science,2003,86(10):3206-3214.
[9] 国家科学研究委员会组织.奶牛营养需要[M].孟庆翔,译.北京:中国农业大学出版社,2002.
[10] 冯仰廉.反刍动物营养学[M].北京:科学出版社,2004.
[11] LAMMERS B P,HEINRICHS A J,KENSINGER R S.The effects of accelerated growth rates and estrogen implants in prepubertal Holstein heifers on estimates of mammary development and subsequent reproduction and milk production[J].Journal of Dairy Science,1999,82(8):1753-1764.
[12] WILDMAN E E,JONES G M,WAGNER P E,et al.A dairy cow body condition scoring system and its re⁃lationship to selected production characteristics[J].Journal of Dairy Science,1982,65(3):495-501
[13] 李文,刘小林,文利侠,等.中国荷斯坦后备奶牛生长发育规律的研究[J].畜牧兽医杂志,2007,26(5):19-21,24.
[14] 李鹏,杨延周,王晓峰,等.奶牛体况评分(BCS)影响奶牛繁殖力的研究进展[J].上海畜牧兽医通讯,2009(2):16-17.
[15] 段智勇.反刍动物日粮中淀粉与纤维的组合效应及其机理的研究[D].博士学位论文.杭州:浙江大学,2006.
[16] 陈伟健,段智勇,阎伟杰,等.反刍动物饲料组合效应在养分吸收和能量利用上的研究进展[J].中国畜牧兽医文摘,2007,43(6):32-33.
[17] SHANNON A D,LASCELLES A K.Changes in the concentration of lipids and some other constituents in the blood plasma of calves from birth to 6 months of age[J].Australian Journal of Biological Sciences,1966,19(5):831-839.
[18] STANLEY C C,WILLIAMS C C,JENNY B F,et al.Effects of feeding milk replacer once versus twice dai⁃ly on glucose metabolism in Holstein and Jersey calves[J].Journal of Dairy Science,2002,85(9):2335-2343.
[19] 门小明.三种精粗比日粮条件下空怀小尾寒羊母羊的消化代谢及部分血液指标的研究[D].硕士学位论文.乌鲁木齐:新疆农业大学,2006.
[20] 董志岩,叶鼎承,李忠荣,等.理想蛋白质氨基酸模式对生长猪生产性能、血清尿素氮及游离氨基酸的影响[J].家畜生态学报,2010,31(5):30-34.
[21] SEJRSEN K,HUBER J T,TUCKER H A,et al.Influ⁃ence of nutrition on mammary development in pre⁃and postpubertal heifers[J].Journal of Dairy Science,1982,65(5):793-800.
[22] SEJRSEN K,PURUP S.Influence of prepubertal feed⁃ing level on milk yield potential of dairy heifers:a re⁃view[J].Journal of Animal Science,1997,75(3):828-835.
[23] SEJRSEN K.Mammary development and milk yield in relation to growth rate in dairy and dual⁃purpose heif⁃ers[J].Acta Agriculturae Scandinavica,1978,28(1):41-46.
[24] SEJRSEN K,PURUP S,VESTERGAARD M,et al.High body weight gain and reduced bovine mammary growth:physiological basis and implications for milk yield potential[J].Domestic Animal Endocrinology,2000,19(2):93-104.
[25] 钱云霞,钱凯先.雌激素相关受体ERR的功能及其调控[J].生物化学与生物物理进展,2005,32(6):495-500.
[26] PURCHAS R W,MACMILLAN K L,HAFS H D.Pi⁃tuitary and plasma growth hormone levels in bulls from birth to one year of age[J].Journal of Animal Science,1970,31(2):358-363.
[27] SEJRSEN K,HUBER J T,TUCKER H A.Influence of amount fed on hormone concentrations and their re⁃lationship to mammary growth in heifers[J].Journal of Dairy Science,1983,66(4):845-855.
[28] MEYER M J,CAPUCO A V,ROSS D A,et al.De⁃velopmental and nutritional regulation of the prepuber⁃tal bovine mammary gland:Ⅱ.Epithelial cell prolifera⁃tion,parenchymal accretion rate,and allometric growth [J].Journal of Dairy Science,2006,89(11):4298-4304.
[29] HAYDEN J M,WILLIAMS J E,COLLIER R J.Plas⁃ma growth hormone,insulin⁃like growth factor,insu⁃lin,and thyroid hormone association with body protein and fat accretion in steers undergoing compensatory gain after dietary energy restriction[J].Journal of Ani⁃mal Science,1993,71(12):3327-3338.
[30] EHRHARDT R A,BELL A W,BOISCLAIR Y R.A⁃nalysis of leptin and leptin receptor mRNA expression in the sheep placenta[J].Journal of Animal Science,1999,77(Suppl.1):159.
[31] CHELIKANI P K,AMBROSE D J,KEISLER D H,et al.Effects of dietary energy and protein density on plasma concentrations of leptin and metabolic hor⁃mones in dairy heifers[J].Journal of Dairy Science,2009,92(4):1430-1441.
[32] 牛淑玲,张才,张国才,等.饲粮处理对奶牛产奶量及脂肪组织瘦素基因表达的影响[J].中国农业科学,2007,40(10):2382-2386.
[33] 栗丽.能量限制通过调节IGF⁃1水平影响成年雌性大鼠生殖潜能[D].硕士学位论文.汕头:汕头大学,2012.
(责任编辑 王智航)
[34] YU H,ROHAN T.Role of the insulin⁃like growth fac⁃tor family in cancer development and progression[J].Journal of the National Cancer Institute,2000,92 (18):1472-1489.
Effects of Diets with Different Energy Levels on Growth Performance and Serum Indices in Chinese Holstein Heifers Aged 7 to 10 Months
ZENG Shuqin1DIAO Qiyu1WANG Jianfen2WANG Jun3SI Bingwen1YANG Chuntao1TU Yan1∗
(1.Feed Research Institute,Chinese Academy of Agricultural Sciences,Beijing Key Laboratory of Dairy Nutrition,Beijing 100081,China;2.Yanqing County Animal Husbandry Technical Extension Station of Beijing,Beijing 102100,China;3.The Animal Husbandry Station of Beijing,Beijing 100029,China)
∗Corresponding author,professor,E⁃mail:tuyan@caas.cn
Abstract:The objective of this study was to investigate the effects of diets with different energy levels on growth performance and serum indices in Chinese Holstein heifers aged 7 to 10 months.Twenty four heifers aged(200±3)days were randomly divided into 3 dietary treatments with 8 replicates and 1 heifer per repli⁃cate,and heifers were fed diets with net energy for lactation(NEL)of 5.40(group A),5.90(group B)and 6.40(group C)MJ/kg,respectively.The pre⁃experiment lasted for 10 days,and the experiment lasted for 90 days.At 210,240,270 and 300 days of age,growth performance indices including body weight,withers height,body length,heart girth,abdominal girth,hip width,nipple length and body condition scoring were determined before morning feeding,and blood samples were collected for the analysis of serum indices inclu⁃ding triglyceride,urea nitrogen,estrogen,estradiol,leptin,growth hormone and insulin⁃like growth factor 1.The results showed as follows:1)at 210 to 300 days of age,the average daily gain of heifers in groups A,B and C was 0.69,0.81,0.96 kg/d,respectively,and group C was significantly higher than group A(P<0.05).2)At 240 days of age,the length of left posterior nipple of heifers in group A was significantly higher than that in group C(P<0.05),while at 240 and 270 days of age,the length of right posterior nipple of heif⁃ers in both groups A and B was significantly higher than that in group C(P<0.05).3)At 240 days of age,the content of serum triglyceride of heifers in groups A and B was significantly lower than that in group C(P<0.05).4)At 300 days of age,the content of serum estradiol of heifers in group C was significantly higher than that in groups A(P<0.05)and C(P<0.01).In conclusion,the body figure and development of mam⁃mary gland of heifers can be maintained when dietary NELis 5.90 MJ/kg under the conditions in the present study.[Chinese Journal of Animal Nutrition,2015,27(2):606⁃615]
Key words:dietary energy level;heifers;growth performance;serum index;nipple development
通信作者:∗屠 焰,研究员,硕士生导师,E⁃mail:tuyan@caas.cn
作者简介:曾书秦(1988—),女,重庆人,硕士研究生,动物营养与饲料科学专业。E⁃mail:zengshuqin0719@sina.com
基金项目:奶牛产业技术体系北京市创新团队;国家科技支撑计划项目课题“华北农区及北方大城市奶牛健康养殖生产技术集成及产业化示范”(2012BAD12B06)
收稿日期:2014-08-26
doi:10.3969/j.issn.1006⁃267x.2015.02.033
文章编号:1006⁃267X(2015)02⁃0606⁃10
文献标识码:A
中图分类号:S823
