非小细胞肺癌组织中磷酸甘油酸变位酶1表达及其与患者预后的关系
2015-12-02潘威孙倩李舒展张喜英曹水任秀宝
潘威,孙倩,李舒展,张喜英,曹水,任秀宝
(天津医科大学肿瘤医院,国家肿瘤临床医学研究中心,天津市肿瘤免疫与生物治疗重点实验室,天津300060)
有氧糖酵解是肿瘤的重要特征之一。磷酸甘油酸变位酶1(PGAM1)是糖酵解途径中的重要酶之一,催化3-磷酸甘油酸生成2-磷酸甘油酸[1]。在多种肿瘤组织中存在 PGAM1 mRNA 高表达[2~5]。但目前关于PGAM1 mRNA在肺癌中的表达,特别是在非小细胞肺癌(NSCLC)中表达的研究报道较少。本研究探讨了NSCLC组织中PGAM1 mRNA的表达及其与患者预后的关系。
1 资料与方法
1.1 临床资料 选择2004年1月~2006年12月天津医科大学肿瘤医院手术切除的NSCLC组织标本206份,其中28份包含配对癌旁正常组织。患者男136例、女70例,年龄38~84岁、中位年龄61岁,吸烟144例;组织学类型:腺癌74例,鳞癌111例,其他(主要是大细胞肺癌和腺鳞癌)21例;临床分期:Ⅰ期101例,Ⅱ~Ⅳ期105例;病理T分期:pT1期140例,pT2~4期66例;有淋巴结转移91例,远处转移32例。所有患者术前未接受放化疗,术后经组织病理学检查明确诊断。
1.2 PGAM1 mRNA表达检测 所有组织标本手术切除后立即置入液氮中,-80℃超低温冰箱长期冻存备用。TRIzol法提取NSCLC组织中总RNA,分光光度计测RNA的浓度和纯度,应用Oligo(dT)引物和M-MLV逆转录酶提取总RNA,逆转录成cDNA。获得的cDNA产物采用SYBR GreenⅡ染料进行实时荧光定量PCR检测PGAM1 mRNA相对表达量。引物序列:PGAM1:上游引物5'-GGCATTGTCAAGCATCTGGAG-3',下游引物5'-GAATACCAGTCGGCAGGTTCA-3';内参 β-actin:上游引物 5'-TGGCACCCAGCACAATGAA-3',下游引物5'-CTAAGTCATAGTCCGCCTAGAAGCA-3'。所有引物热循环条件相同,反应体系均为20 μL。反应条件:95℃ 5 min,94 ℃ 30 s,56 ℃ 30 s,72 ℃ 30 s,共40 个循环,最后72℃延伸10 min。采用2-ΔΔCt法计算 PGAM1 mRNA的相对表达量。以ROC曲线中约登指数最高值(1.96)作为 PGAM1 mRNA表达水平的分割点,>1.96为 PGAM1 mRNA 高表达,≤1.96为PGAM1 mRNA低表达。ROC曲线见图1。
1.3 随访 所有患者电话随访1~80个月、中位随访34个月,失访10例。采用Kaplan-Meier生存分析法计算不同PGAM1 mRNA表达NSCLC患者的总生存期(OS)、无病生存期(DFS)。

图1 PGAM1 mRNA表达水平的分割点确定
1.4 统计学方法 采用SPSS21.0统计软件。计量资料比较采用t检验,计数资料比较采用χ2检验。采用Cox比例风险回归模型分析不同PGAM1 mRNA表达与NSCLC患者预后的关系;采用Log-rank检验比较不同PGAM1 mRNA表达NSCLC患者OS、DFS。P<0.05为差异有统计学意义。
2 结果
2.1 28例NSCLC患者肿瘤组织及其癌旁正常组织PGAM1 mRNA表达情况 28例NSCLC组织中PGAM1 mRNA相对表达量平均为1.94,癌旁正常组织为1.25,二者比较差异有统计学意义(t=2.586,P<0.05)。其中 NSCLC组织与癌旁正常组织PGAM1 mRNA相对表达最高比值达到5.18,其中比值 >1.5者15例(53.6%)。见插页Ⅰ图1。
2.2 PGAM1 mRNA表达与NSCLC患者临床病理特征的关系 见表1。
2.3 PGAM1 mRNA表达与NSCLC患者预后的关系 见插页Ⅰ图2。PGAM1 mRNA高表达80例、低表达126例。PGAM1 mRNA高表达者OS、DFS中位时间分别为23、14个月,低表达者分别为42、24个月,二者比较P均<0.01。
Cox比例风险回归模型单因素分析显示,患者OS、DFS与PGAM1 mRNA高表达、病理T分级高、有淋巴结转移及远处转移有关(P均<0.05),与患者性别、年龄无关(P均>0.05);多因素分析显示,PGAM1 mRNA高表达、淋巴结转移和远处转移是NSCLC 患者 OS(HR 分别为1.730、2.971、3.641)和DFS(HR 分别为1.698、2.649、2.896)的独立影响因素(P 均 <0.05)。
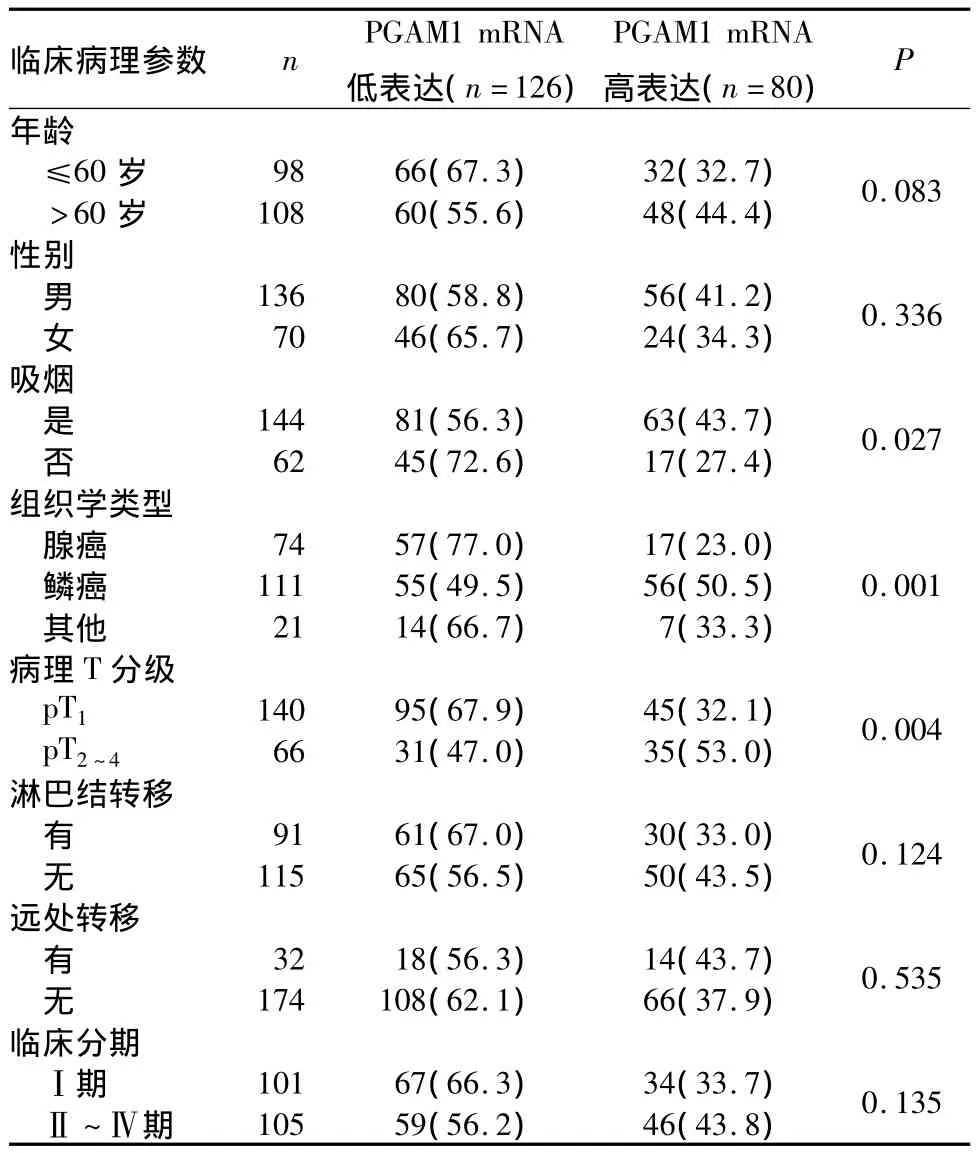
表1 不同PGAM1 mRNA表达与NSCLC患者临床病理参数的关系[例(%)]
3 讨论
上世纪20年代德国科学家Otto Warburg等研究发现,肿瘤细胞无论氧气充足与否,均选择产能效率较低的糖酵解途径产能,并且其葡萄糖消耗、糖酵解速率及乳酸生成均较正常细胞明显增加。这种现象被称为“Warburg效应”或“有氧糖酵解[6]。有氧糖酵解通过消耗大量的葡萄糖来产生肿瘤细胞生物合成所需要的含碳中间产物。研究证实,糖酵解酶的过表达是有氧糖酵解的促进因素之一[5]。PGAM1为糖酵解通量的关键分子,催化用于合成代谢的大部分碳源中间产物的合成;进一步研究发现,PGAM1参与肿瘤的生长调节[2]。质谱分析研究显示,肝癌、结肠癌和肺癌组织中PGAM1 mRNA表达明显增加[2,3,7],并伴随着酶活性增加而增加[4,8]。Chen等[9]研究发现,在肺腺癌组织中PGAM1 mRNA 呈高表达。Kondoh等[10]报道,PGAM1 mRNA 在肿瘤组织中高表达是由肿瘤抑制基因TP53丢失所致,与 Bensaad 等[11,12]报道 TP53 对细胞糖酵解具有负调节作用一致。由此推测,PGAM1 mRNA高表达是维持肿瘤生长和功能的关键因子之一。PGAM1 mRNA高表达可增强乳腺癌细胞系上皮间质转化的能力,增加肺癌细胞对淋巴血管及胸膜的侵袭力[13]。但其在NSCLC组织中的报道较少。
本研究首次系统地评估了PGAM1 mRNA表达与NSCLC患者预后的关系。结果显示,NSCLC组织PGAM1 mRNA表达显著高于癌旁正常组织,与Chen等[9]在肺腺癌组织中的结果一致。PGAM1 mRNA高表达与患者吸烟、组织类型为鳞癌和病理T分级高有关,提示PGAM1参了与NSCLC的发生、发展过程。本研究还发现,PGAM1 mRNA高表达者OS、DFS均低于PGAM1 mRNA低表达者;Cox比例风险回归模型分析显示,PGAM1 mRNA高表达是影响患者OS、DFS的独立危险因素。故认为NSCLC组织中PGAM1 mRNA表达水平与患者预后有关,PGAM1 mRNA可作为判断患者预后的候选肿瘤分子标记物。
综上所述,PGAM1 mRNA在NSCLC组织中表达升高,其高表达是患者预后不良的独立危险因素。
[1]Fothergill-Gilmore LA,Watson HC.The phosphoglycerate mutases[J].Adv Enzymol Rela Areas Mol Biol,1989,(62):227-313.
[2]Li C,Xiao Z,Chen Z,et al.Proteome analysis of human lung squamous carcinoma[J].Proteomics,2006,6(2):547-558.
[3]Shen J,Wang W,Wu J,et al.Comparative proteomic profiling of human bile reveals SSP411 as a novel biomarker of cholangiocarcinoma[J].PLoS One,2012,7(10):e47476.
[4]Durany N,Joseph J,Campo E,et al.Phosphoglycerate mutase,2,3-bisphosphoglycerate phosphatase and enolase activity and isoenzymes in lung,colon and liver carcinomas[J].Br J Cancer,1997,75(7):969-977
[5]Vander Heiden MG,Cantley LC,Thompson CB.Understanding the Warburg effect:the metabolic requirements of cell proliferation[J].Science,2009,324(5930):1029-1033.
[6]Hanahan D,Weinberg RA.Hallmarks of cancer:the next generation[J].Cell,2011,144(5):646-674.
[7]Lei Y,Huang K,Gao C,et al.Proteomics identification of ITGB3 as a key regulator in reactive oxygen species-induced migration and invasion of colorectal cancer cells[J].Mol Cell Proteomics,2011,10(10):M110.005397.
[8]Durany N,Joseph J,Jimenez OM,et al.Phosphoglycerate mutase,2,3-bisphosphoglycerate phosphatase,creatine kinase and enolase activity and isoenzymes in breast carcinoma[J].Br J Cancer,2000,82(1):20-27.
[9]Chen G,Gharib TG,Wang H,et al.Protein profiles associated with survival in lung adenocarcinoma[J].Proc Natl Acad Sci U S A,2003,100(23):13537-13542.
[10]Kondoh H,Lleonart ME,Gil J,et al.Glycolytic enzymes can modulate cellular life span[J].Cancer Res,2005,65(1):177-185.
[11]Bensaad K,Tsuruta A,Selak MA,et al.TIGAR,a p53-inducible regulator of glycolysis and apoptosis[J].Cell,2006,126(1):107-120.
[12]Madan E,Gogna R,Bhatt M,et al.Regulation of glucose metabolism by p53:emerging new roles for the tumor suppressor[J].Oncotarget,2011,2(12):948-957.
[13]Ishikawa M,Inoue T,Shirai T,et al.Simultaneous expression of cancer stem cell-like properties and cancer-associated fibroblastlike properties in a primary culture of breast cancer cells[J].Cancers(Basel),2014,6(3):1570-1578.
