菊科紫菀属3种植物的核型分析
2015-06-28王梓辛耿胜娟陈三茂黎维平
李 志,王梓辛,耿胜娟,陈三茂,黎维平
(湖南师范大学生命科学学院,长沙410081)
菊科紫菀属3种植物的核型分析
李 志,王梓辛,耿胜娟,陈三茂,黎维平*
(湖南师范大学生命科学学院,长沙410081)
菊科紫菀属密毛系(Aster Series Vestiti Ling)含3个物种:密毛紫菀(A.vestitus Franch.)、灰枝紫菀(A.poliothamnus Diels)和西固紫菀(A.sikuensis W.W.Smith et Farr.),新近的分子系统学研究对密毛系是否单系提出质疑。该文采用常规根尖压片法,对紫菀属密毛系3种植物4个居群的核型进行观察分析。结果显示:3种植物的核型公式均为2n=2x=18=14m+4sm(2SAT);密毛紫菀居群2个的核型属于1A型,灰枝紫菀和西固紫菀为2A型。密毛紫菀和西固紫菀的核型资料为首次报道。灰枝紫菀的染色体数目、基数和不对称性等核型资料与先前唯一的相关报道结果(体细胞染色体数目为32,核型公式2n=2x=32=26sm+6st,属4A型,染色体中未发现随体)迥然不同。3种植物的核型结果不支持保留密毛系,而核型不对称性类型支持该实验室先前提出的亚属划分。3种植物随体的大小属于紫菀亚族型,这支持欧亚紫菀属与北美紫菀类没有密切亲缘关系的观点。
紫菀属;密毛系;密毛紫菀;灰枝紫菀;西固紫菀;核型
紫菀属(Aster L.)是菊科的模式属,其定义近年发生了巨大的改变。DNA序列证据[1-2]显示,广义紫菀属(Aster sensu lato)既是多系类群,又是并系类群。最近的几个研究[2-4]分别重新定义了紫菀属,一方面将一些类群排除出紫菀属,另一方面又建议将一些属归并进来。分子系统学研究与早先的一些细胞学研究在一定程度上相吻合。例如染色体基数的差异和核型特征[3]支持分子系统学研究将北美紫菀类植物从紫菀属中独立出去的建议[1-2]。原马兰属(Kalimeris Cass.)的物种,除长柄马兰[K.longipetiolata(Chang)Ling]外,虽然已并入紫菀属[2,4],但是马兰类均具有小型染色体[5],是其作为一自然类群的有力证据之一,马兰类在分子系统树上也聚为一支[2]。新定义的紫菀属中,有细胞学资料的种类只占很少的一部分[5-14]。新的细胞学资料累积有利于紫菀属系统演化和分类学的深入研究。
林镕等[15]建立的密毛系(Aster Series Vestiti Ling)由密毛紫菀(A.vestitus Franch.)、灰枝紫菀(A.poliothamnus Diels)和西固紫菀(A.sikuensis W.W.Smith et Farr.)3个种组成;其中灰枝紫菀和西固紫菀均为亚灌木,特产于中国西部,而密毛紫菀为多年生草本,为喜马拉雅区系特有植物[4,15]。虽然分子研究支持将这3个种留在紫菀属内,但认为密毛系是多系类群,密毛紫菀被置于一个亚属,而将灰枝紫菀和西固紫菀归入另一亚属,后2种植物的亲缘关系密切[2],但分子系统学的结论还有待更多证据包括细胞学资料的支持。尤其困惑的是有报道[16]灰枝紫菀的染色体数目为2n=32,为二倍体。但是一般认为,染色体基数为9是紫菀属乃至欧亚紫菀族的特征[3,5-14]。紫菀属植物具有染色体基数为16的报道很不寻常,值得验证。本研究首次报道密毛紫菀和西固紫菀的核型,探讨3种植物核型差异的分类学意义,并检验前人对灰枝紫菀细胞学特征的报道,讨论新定义的紫菀属植物染色体的随体特征。
1 材料和方法
密毛紫菀、灰枝紫菀和西固紫菀的居群取样分别于近年在云南香格里拉的桑那(桑那居群Ps)和小中甸(小中甸居群Px)、陕西旬阳小河镇和甘肃贵清山(表1),凭证标本保存于湖南师范大学植物标本室(HNNU)。
西固紫菀植株被移栽后取根尖;而密毛紫菀和灰枝紫菀则用瘦果培育成幼苗,然后获取根尖。以个体为单位切取根尖,经0.1%秋水仙碱中于室温下预处理4h,卡诺固定液(冰醋酸∶95%乙醇=1∶3)中固定4~18h,用1mol/L盐酸于60℃下解离8min,充分漂洗后,用改良苯酚品红染液浸泡0.5h以上,最后压片、镜检、拍照。
每个居群观察至少5株20个以上细胞有丝分裂中期照片(染色体分散较好且着丝粒清晰),进行染色体记数。核型分析根据李懋学和陈瑞阳的方法[17]。核型不对称系数(As K%)计算公式为:As K%=长臂总长/全组染色体总长×100。核型分类按Stebbins的标准[18]划分。
2 结果与分析
密毛紫菀2个居群,灰枝紫菀和西固紫菀体细胞的染色体数目均为18,各有1对染色体具有随体,核型公式也均为2n=2x=18=14m+4sm(2SAT)(图1;表2)。但是,染色体参数在种间及居群间存在差异(表2),密毛紫菀的核型属于1A型,灰枝紫菀和西固紫菀的核型为2A型,但灰枝紫菀的核型(图1C,D,I;表2)与先前报道[16]的截然不同。
3 讨 论
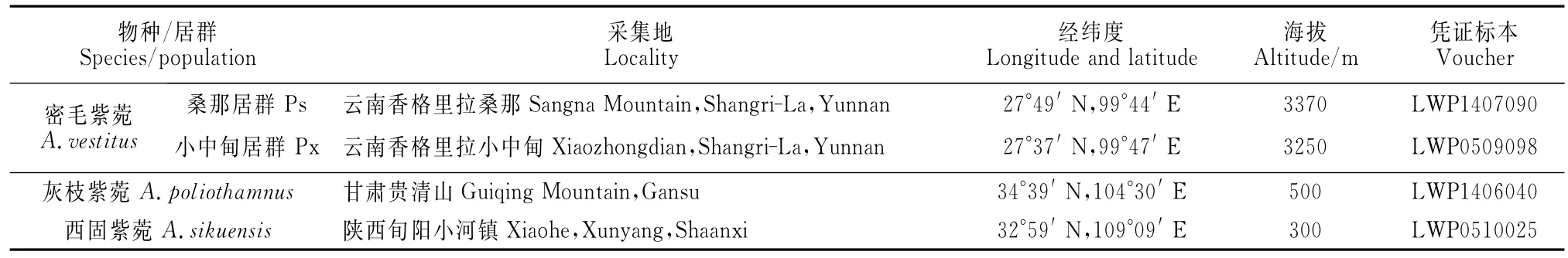
表1 3种紫菀属植物4居群的野外采集信息Table 1 Information of four populations sampled
3.1 灰枝紫菀的核型
先前的报道[16]指出,灰枝紫菀为二倍体,体细胞染色体数目为32,核型公式2n=2x=32=26sm+6st,属4A型,染色体中未发现随体。但本研究结果显示,灰枝紫菀体细胞染色体数为18,属于二倍体,染色体基数为9,这与迄今为止其他有关欧亚紫菀属植物乃至欧亚紫菀族植物的染色体基数[3,5-14]是一致的。先前报道灰枝紫菀的体细胞染色体数目为32,其染色体基数为16(其被当成二倍体时[16])或8(若其被当成四倍体时);但是至今没有研究认为菊科体细胞染色体基数高达16,而且在紫菀族中,x=8不见于中国[2]。紫菀属植物核型不对称性通常较低[8-12],本研究认为灰枝紫菀的核型属于2A型,这在紫菀属中属于进化类型;而关于其核型属于4A型的结论[16]是令人意外的,紫菀族从未有过如此高的核型不对称性的报道。综上所述,先前关于灰枝紫菀核型的研究难以令人置信,有必要检查其凭证标本或做重复实验。
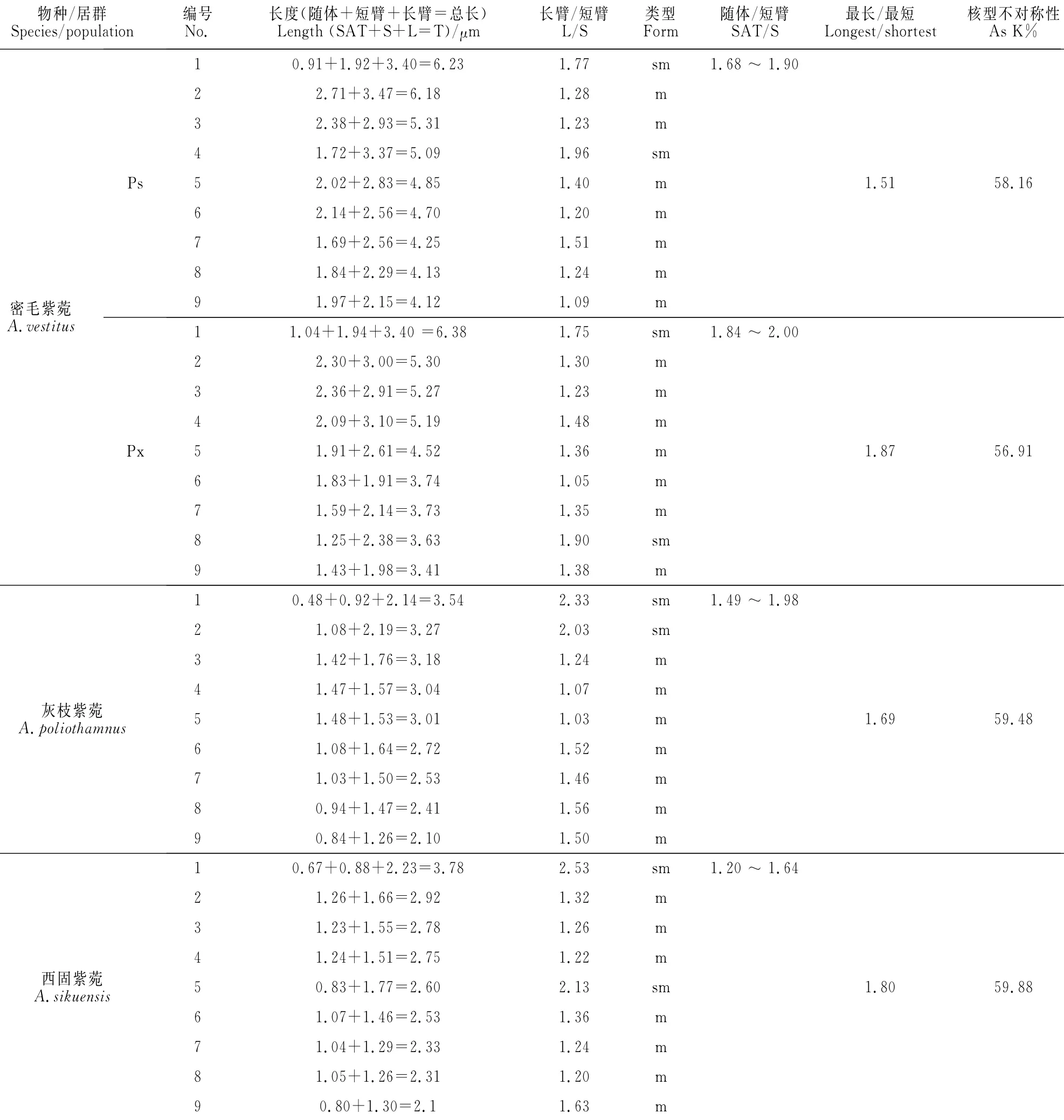
表2 3种紫菀属植物4居群的核型参数Table 2 Karyotypic parameters of four populations of three Aster species
3.2 3个物种间核型差异的系统学意义
密毛紫菀、灰枝紫菀和西固紫菀的核型公式一致;三者虽然在最长与最短染色体长度比值和As K%值上有差异,但种间差异小于密毛紫菀二居群间差异。值得注意的是,密毛紫菀的染色体明显大于灰枝紫菀和西固紫菀的染色体,密毛紫菀的核型不对称性(1A型)则小于灰枝紫菀和西固紫菀的(2A型),这些差异具有系统学意义。密毛紫菀属于原亚属(含属的模式种雅美紫菀[A.amellusL.)][2],同属原亚属的植物三脉紫菀(A.ageratoides Turcz.)[56]、川鄂紫菀(Aster moupinensis Hand.-Mazz.)[8]、神农架紫菀(Aster shennongjiaensis)[9]的核型不对称性都较低,平均臂比都小于1.40,核型不对称类型为1A;而与灰枝紫菀和西固紫菀属于另一亚属[2]的东风菜[A.scaber Thunb.=Doellingeria scaber(Thunb.)Nees][19]和莎菀[A.gramineus(L.)Komarov.=Arctogeron gramineum(L.)DC.][14]则均具有2A核型。所以核型证据不支持将密毛紫菀、灰枝紫菀和西固紫菀构成一个系,即密毛系并不是一个单系的自然类群,而倾向于将它们置于2个不同的亚属,这支持本实验室早先基于分子证据的建议[2]。随着紫菀属植物细胞学资料的不断积累,核型不对称性类型的差异可能被确认为紫菀属2亚属的特征。不过,虽然灰枝紫菀和西固紫菀的染色体长度明显小于密毛紫菀,但与密毛紫菀同属于原亚属的马兰类植物和川鄂紫菀具有更小的染色体,所以染色体大小不能作为划分亚属的依据,而在组或系的水平上可能具有分类学价值。灰枝紫菀和西固紫菀均为2A核型,染色体都较小(<4μm),核型不对称系数也接近,在已报道的分子系统树[2]上二者组成一个分支,可见核型资料和分子证据都建议二者具有密切亲缘。

图1 紫菀属3种植物的有丝分裂中期分裂相(A~F)及其核型图(G~J)箭头指随体;A、G.密毛紫菀的桑那居群;B、H.密毛紫菀的小中甸居群;C、D、I.灰枝紫菀;E、F、J.西固紫菀Fig.1 Photomicrographs of somatic metaphase(A-F)and the karyotypes(G-J)of the three Aster species Arrows indicate satellites;A,G.Ps of A.vestitus;B,H.Px of A.vestitus;C,D,I.A.poliothamnus;E,F,J.A.sikuensis
3.3 随体的分类学价值
Semple等[20-21]将具有的随体染色体称为核仁组织者区染色体(The nucleolar organizer region chromosome,NOR chromosome)。他们根据随体的形态,将传统的广义紫菀属植物分为3类:(1)原始型(Primitive type),即随体长度大体等于其所相连的短臂,以Doellingeria umbellate Nees(=Aster umbellatus Mill.)为代表[20-21];(2)真紫菀型(Euaster type),其随体长于短臂,见于Eurybia radula(Ait.)GL Nesom(=Aster radula Ait.)等植物[20-21];(3)Virgulus型(Virgulus type),其随体非常微小,远远小于其所连接的短臂,如Symphyotrichum novae-angliae(L.)Nesom(A.novae-angliae L.)和Lasallea Greene的数个种[20-22]。上述3类限于新世界紫菀类植物(北美广义紫菀属Aster sensu lato)。Nesom[3]提出了第4种随体类型,其随体显著短于短臂,但是远远大于Virgulus型随体。此类型限于欧亚紫菀亚族植物,所以称其为紫菀亚族型(Asterinae type)。Nesom定义的紫菀亚族与本实验室最近重新定义的紫菀属[2]相当吻合。所以紫菀亚族型应是紫菀属的细胞学特征。本研究涉及的3个紫菀属植物体细胞均有1对随体染色体,但短臂与其上发随体长度之比在1~2之间,与紫菀亚族型十分相似。先前关于Aster thomsonii Clarke[23]和三脉紫菀核型的研究[5-6,10]也支持紫菀亚族型的概念。因此,认为北美紫菀类与欧亚紫菀属没有密切亲缘而将紫菀属中分离出去[3]是合理的。紫菀属植物的细胞学资料仍十分缺乏,紫菀亚族型是否完全成立还有待进一步研究。
[1] NOYES R D,RIESEBERG L H.ITS sequence data support a single origin for North American Astereae(Asteraceae)and reflect deep geographic divisions in Aster s.l.[J].American Journal of Botany,1999,86(3):398-412.
[2] LI W P,YANG F S,JIVKOVA T,et al.Phylogenetic relationships and generic delimitation of Eurasian Aster(Asteraceae:Astereae)inferred from ITS,ETS and trnL-Fsequence data[J].Annals of Botany,2012,109(7):1 341-1 357.
[3] NESOM G L.Review of the taxonomy of Aster sensu lato(Asteraceae:Astereae),emphasizing the New World species[J].Phytologia,1994,77(3):141-297.
[4] CHEN Y L,BROUILLET L,SEMPLE J C.Flora of China(Volume 20~21)[M].Beijing:Science Press;St.Louis:Missouri Botanical Garden Press,2011:574-632.
[5] LI W P.Natural hybridization between Aster ageratoides var.scaberulus and Kalimeris indica(Asteraceae):evidences from morphology,karyotype and ITS sequences[J].Botanical Studies,2006,47(2):191-197.
[6] CHEN R Y,IRIFUNE K,SONG W Q,et al.Cytogenetics on Plant Correlating between Japan and China[M].Hiroshima:Hiroshima University,1992:89-112.
[7] ITO M,SOEJIMA A.Flora of Japan(Volume IIIb)[M].Tokyo:Kodansha,1995:59-73.
[8] LI W P(黎维平).A study on morphology and cytology of Aster moupinensis(Asteraceae)from the Three Gorge Reservior Area[J].Life Science Research(生命科学研究),2003,7(1):70-72(in Chinese).
[9] LI W P,ZHANG Z G.Aster shennongjiaensis(Asteraceae),a new species from central China[J].Botanical Bulletin of Academia Sinica,2004,45(1):95-99.
[10] LI W P.The Cytogeography of Aster ageratoides var.laticorymbus(Asteraceae),apolyploid complex endemic to China[J].Botanical Bulletin of Academia Sinica,2005,46(4):355-361.
[11] LI W P(黎维平),LIU SH X(刘胜祥).Differentiation between Aster ageratoides var.micranthus and var.gerlachii:evidence from morphology and cytology[J].Acta Phytotaxonomica Sinica(植物分类学报),2005,43(1):31-36(in Chinese).
[12] LI W P(黎维平),LIU SH X(刘胜祥).Cytogeography of Aster ageratoides var.scaberulus[J].Life Science Research(生命科学研究),2005,9(1):68-72(in Chinese).
[13] LI W P(黎维平),ZHANG P(张 平),YIN G SH(殷根深).New evidence for the tribal placement of Sheareria within Astereae(Compositae)[J].Journal of Systematics and Evolution(植物分类学报),2008,46(4):608-613(in Chinese).
[14] 周广明.莎菀属及其近缘属植物(菊科)的系统学研究[D].山东曲阜:曲阜师范大学,2011.
[15] 林 镕,陈艺林,石 铸.中国植物志(第74卷)[M].北京:科学出版社,1985:70-353.
[16] GONG H D(巩红冬).Study on karyotypes of Aster poliothamnus in the Eastern of Qinghai-Tibet Plateau[J].Guangdong Agricultural Science(广东农业科学),2010,3(3):60-61(in Chinese).
[17] LI M X(李懋学),CHEN R Y(陈瑞阳).A suggestion on the standardization of karyotype analysis in plants[J].Journal of Wuhan Botanical Research(武汉植物学研究),1985,3(4):297-302(in Chinese).
[18] STEBBINS G L.Chromosomal Evolution in Higher Plants[M].London:Edward Arnold,1971:87-123.
[19] WANG B(王 冰),LI N(李 楠),GU X J(顾晓洁),et al.Karyotype analysis of Doellingeria scaber chromosome[J].Chinese Pharmaceutical Journal(中国医药杂志),2006,41(10):732-734(in Chinese).
[20] SEMPLE J C,BROUILLET L.Chromosome numbers and satellite chromosome morphology in Aster and Lasallea[J].American Journal of Botany,1980,67(7):1 027-1 039.
[21] SEMPLE J C,CHMIELEWSKI J G,CHINNAPPA C C.Chromosome number determinations in Aster L.(Compositae)with comments on cytogeography,phylogeny and chromosome morphology[J].American Journal of Botany,1983,70(10):1 432-1 443.
[22] CHMIELEWSKI J G,SEMPLE J C.The biology of Canadian weeds.125.Symphyotrichum ericoides(L.)Nesom(Aster ericoides L.)and S.novae-angliae(L.)Nesom(A.novae-angliae L.)[J].Canadian Journal of Plant Sciences,2003,83(4):1 017-1 037.
[23] MEHRA P N,REMANANDAN P.Cytological investigations on the Indian Compositae.II.Astereae,Heliantheae,Helenieae and Anthemideae[J].Caryologia,1974,27(3):255-284.
(编辑:宋亚珍)
A Karyotypic Study on Three Aster Species(Asteraceae)
LI Zhi,WANG Zixin,GENG Shengjuan,CHEN Sanmao,LI Weiping*
(College of Life Sciences,Hunan Normal University,Changsha 410081,China)
Aster vestitus Franch.,A.poliothamnus Diels and A.sikuensis W.W.Smith et Farr.comprise Aster Series Vestiti Ling(Asteraceae)whose monophyly was not supported by a new molecular phylogenetic investigation.Karyotypes of four populations of the three species were analyzed by conventional root tip squash method.The results showed that all the three species had a karyotypic formula of 2n=2x=18=14m+4sm(2SAT),while the two populations of A.vestitus belonged to the Stebbins’1Akaryotype and the other two species to the Stebbins’2A.The karyotypic data of A.vestitus and A.sikuensis were reported for the first time and that of A.poliothamnus was quite different from that reported by the only previous related study.The results did not suggest maintaining Ser.Vestiti but support the comment of Li et al.on subgeneric classification.Satellite size of the three species belonged to“Asterinae type”,which supported the opinion that there is no close relationship between Eurasian Aster and North American asters.
Aster;Series Vestiti;A.vestitus;A.poliothamnus;A.sikuensis;karyotypes
Q343.2+2
A
10.7606/j.issn.1000-4025.2015.06.1148
1000-4025(2015)06-1148-05
2015-01-27;修改稿收到日期:2015-04-16
国家自然科学基金面上项目(31370265);湖南省生态学重点学科建设项目(0713);湖南省生物发育工程及新产品研发协同创新中心(20134486)
李 志(1989-),男,在读博士研究生,主要从事植物分类学研究。E-mail:442050223@qq.com
*通信作者:黎维平,博士,教授,博士研究生导师,主要从事菊科紫菀族的系统分类研究。E-mail:lwp@hunnu.edu.cn
