DNS法检测灰略红链霉菌JSD1产纤维素酶的CMC酶活条件的优化
2015-06-05罗艳青冯海玮柴晓彤支月娥
罗艳青,张 丹,冯海玮,柴晓彤,支月娥,周 培
(上海交通大学农业与生物学院,上海交通大学陆伯勋食品安全研究中心,农业部都市农业(南方)重点实验室,上海 200240)
罗艳青,张 丹,冯海玮,柴晓彤,支月娥*,周 培
(上海交通大学农业与生物学院,上海交通大学陆伯勋食品安全研究中心,农业部都市农业(南方)重点实验室,上海 200240)
为研究3,5-二硝基水扬酸法(DNS)检测纤维素酶的CMC酶活条件,采用单因素实验探讨了波长、酶促反应温度与时间、pH、粗酶液和底物添加量、底物浓度、显色时间及空白实验对测定结果的影响,并通过响应面(Box-Behnken)实验设计及方差分析确定了一种较优的CMC酶活检测条件组合。结果表明,CMC酶活检测最佳条件为:波长520nm、酶促反应温度和时间分别为45℃和30min、pH7.0、底物浓度12g/L、显色15min。
3,5-二硝基水扬酸法,响应面,CMC酶活检测条件
纤维素(Cellulose)是地球上最丰富的可再生生物高聚物[1],这类物质大多以秸秆的形式存在,占植物秸秆干质量的 40%至50%,通常难以被降解,人类对其合理开发利用极其有限[2-3]。目前公认最合理的途径是纤维素的酶学降解,水解此类生物聚合物,酶活测定是不可或缺的部分[4-5]。纤维素酶是一组能够将纤维素降解成较小糖成分(葡萄糖单元)的酶系的总称,CMC酶活力主要代表外切β-1,4葡萄糖苷酶和内切酶的活力总和,在研究中常作为纤维素酶活性的判断依据。不同来源的纤维素酶组分及其活性存在差异,对底物的作用方式也比较复杂,致使纤维素酶活力的测定方法多样化[6-7],造成同一体系用不同方法或者同一方法在不同条件下测定的结果都会有很大的差异。因此,必须选择适宜的测定方法与条件,提高测定结果的准确性与重复性。
3,5-二硝基水扬酸(DNS)比色法具有操作简便、快速、精确等优点,在国内外被广泛采用。Miller GL等[8]和Tasun K等[9]较早的利用DNS法检测还原糖,Goksyr J等[10]进一步证明了利用DNS法检测纤维素酶活的灵敏性以及方法的可靠性。王琳等[11]和孙盈等[12]都对DNS法检测定纤维素酶活的最适条件进行了探讨,邹水洋等[13]则探讨了DNS法测定CMC酶活的空白实验对检测结果的影响。但以上方法得出的结论不适用于本实验室筛选分离的灰略红链霉菌JSD-1[14-15]产CMC酶活力的研究。因此,本文利用单因素法和响应面实验系统分析DNS检测条件,提出了一种最适于JSD-1产CMC酶活的检测方案,为合理的开发利用高产纤维素酶菌株奠定基础。
1 材料与方法
1.1 材料与仪器
菌种:灰略红链霉菌(Streptomycesgriseorubens),编号为CGMCC No. 5706,由本实验室分离并保藏。
试剂:羧甲基纤维素钠(CMC-Na)、3,5-二硝基水杨酸、酒石酸钾钠、苯酚、葡萄糖等 均购自生工生物工程(上海)股份有限公司。
仪器:多功能酶标仪 M200 PRO 瑞士TECAN;双层真彩触摸屏摇床ZWYR-2112B 上海智诚分析仪器制造有限公司;离心机 CT14RD 天美(中国)科学仪器有限公司。
1.2 培养基及菌种培养条件
固体培养基:CMC-Na 10.0g,K2HPO41.0g,MgSO4·7H2O 0.1g,FeSO4·7H2O 0.1g,MnSO41.0×10-4g,蛋白胨10.0g,酵母膏5.0g,琼脂粉15~20g,pH7.2~7.4[16]。
CMC发酵培养基:CMC-Na 10.0g,蛋白胨5.0g,酵母膏1.0g,蒸馏水1000mL,pH7.2~7.4。
培养条件:将灰略红链霉菌(JSD-1)于固体培养基培养3~4d,活化后移一环至灭菌的CMC发酵培养液中,发酵液量为500mL的锥形瓶中装80mL,于32℃,200r/min摇床培养4~5d。
1.3 测定方法
1.3.1 粗酶液的制备 将培养后发酵液于4℃下,8000r/min离心5min,通过0.22μm的一次性针头过滤器收集所得的液体即为粗酶液。
1.3.2 纤维素酶活测定方法 采用DNS法[8,17],以羧甲基纤维素钠(CMC-Na)为底物,取粗酶液0.5mL,加入含0.5%CMC-Na、pH为4.8的柠檬酸缓冲液1.5mL,50℃恒温水浴30min,加入DNS 1.5mL沸水浴显色5min,迅速冷却定容至20mL,摇匀后用多功能酶标仪在特定波长下测定CMC酶活。CMC酶活单位为每分钟每毫升酶液催化纤维素水解生成1.0μg葡萄糖所需的酶量为一个酶活力单位U,即1U/mL。
DNS试剂配制参照高培基方法[18],稍作改进:取酒石酸钾钠91g,溶于500mL蒸馏水中,加热至50℃溶解,再依次加入3,5-二硝基水杨酸3.5g、NaOH 20g、苯酚2.5g、无水亚硫酸钠2.5g,搅拌至完全溶解,冷却后用蒸馏水定容至1L,储于棕色瓶中,放置1周后使用。
1.3.3 葡萄糖标准曲线的绘制 将葡萄糖在80℃恒温干燥烘干至恒重,精确称取0.10g完全溶解于100mL蒸馏水中,具体操作见表1。以葡萄糖含量(mg/mL)为横坐标,以对应的吸光度OD值为纵坐标,绘制葡萄糖标准曲线。

表1 葡萄糖标准曲线的绘制Table 1 Drawing of glucose standard curve
备注:沸水浴中显色5min,冷却定容至20mL,520nm处测定OD值。
1.4 CMC酶活力测定条件
扫描葡萄糖标准液在波长(480、490、500、510、520、530、540、550、560nm);固定波长设置酶促反应温度(30、35、40、45、50、55、60℃)和酶促反应时间(5、10、20、30、40、50、60min)共49种组合;固定波长、酶促温度和时间,设置两种不同pH的缓冲体系:浓度为0.05mol/L的柠檬酸缓冲液体系(pH=4.0、4.4、4.8、5.2、5.8、6.2、6.6)和浓度为0.1mol/L的磷酸缓冲液体系(pH=5.8、6.6、7.0、7.5、8.0);在波长、酶促温度和时间、缓冲液体系确定后将底物浓度(3、5、8、10、12、15、20g/L)与粗酶和底物添加量(0.5+1.5mL、1+3mL、1+2mL、1+1mL、2+2mL)结合设计共35种组合;固定优化后的酶活条件,设置显色时间(3、5、8、10、12、15、20min),测定各单因素和组合实验对CMC酶活力的影响,每一实验设3个重复。
1.5 空白实验处理
条件设定具体方案见表2。

表2 空白实验处理条件的设定Table 2 Set of processing conditions for blank test
1.6 Box-Behnken实验设计
基于上述单因素及组合实验结果,研究温度、pH、反应时间、底物浓度、显色时间五因素对酶活测定结果的影响,实验设计见表3。

表3 响应面因素水平表Table 3 Factors and levels in RSA
1.7 数据分析方法
采用Statistica 8.0软件,Experimental Design模块中的3**(k-p)与 Box-Behnken Design对实验数据进行回归拟合,结合SAS软件对数据进行方差分析,利用OriginPro 8.0软件作图。
2 结果与讨论
2.1 波长对CMC酶活力的影响
由图1可知,OD值在480nm处有最大吸光值,并随着波长的增加逐渐减小,标准曲线的斜率也逐渐降低,不同浓度的标准品在同一波长下测定值的波动范围也随之缩小,这种变化趋势可能与DNS显色液本身在较低波长下有较高的吸光值有关。此外,随着葡萄糖浓度的增加,不同波长检测条件下的OD值差异逐渐增大。结合表4分析可知,还原糖在480nm 处的R2最小(R2=0.9921),吸光值在此波长下稳定度偏低,线性较差,随着波长的增大吸光值的线性回归系数逐渐上升且在520nm 处有最高值(R2=0.99933)。由于是同一批样品在不同波长下扫描的结果值,单个波长下的R2大小可以排除人为操作误差。实验过程中共扫描了3次,平行组间差异较小,文章中呈现的是三组平行的平均值,基本可以排除系统误差。综合线性变化规律和回归系数可得,最适测定波长为520nm,这一结论与文献[19]报道的结果基本一致。

图1 系列葡萄糖标准品与DNS显色后 在不同波长下的OD值Fig.1 OD values under different wavelength of a series of glucose standards colorized by DNS

波长/nm斜率(Slope)截距(Intercept)葡萄糖标准曲线的R2480048270249309921490045680145909961500041480086809972510035830059609980520031430051009993530027280048709978540023350048009967550020470044209957560017550044109939
2.2 酶促反应温度和时间对CMC酶活力的影响
酶促反应温度和时间对CMC酶活力的影响见图2,不同时间段的温度与酶活的趋势呈钟罩型曲线状,5~20min 时CMC酶活在50℃ 出现最高值,30~60min 酶活最高点在45℃ 出现,其中酶促反应30min、45℃出现最大值23.979U/mL。分析其原因:一方面酶活力随着温度的升高而增大,由于酶是蛋白质,当温度偏高时造成部分酶蛋白逐渐变性而失活,从而引起酶反应速率下降;另一方面,酶活力的表达速率随时间的延长而增大,达到一定时间点后其速率增长曲线逐渐平稳并且下降,这主要是因为随着反应进行,底物浓度降低,产物浓度增加,逆反应从无到有逐渐变得显著,所以酶促反应时间以30min为最适。综合以上分析,检测酶活力最佳反应温度和时间分别为45℃、30min。
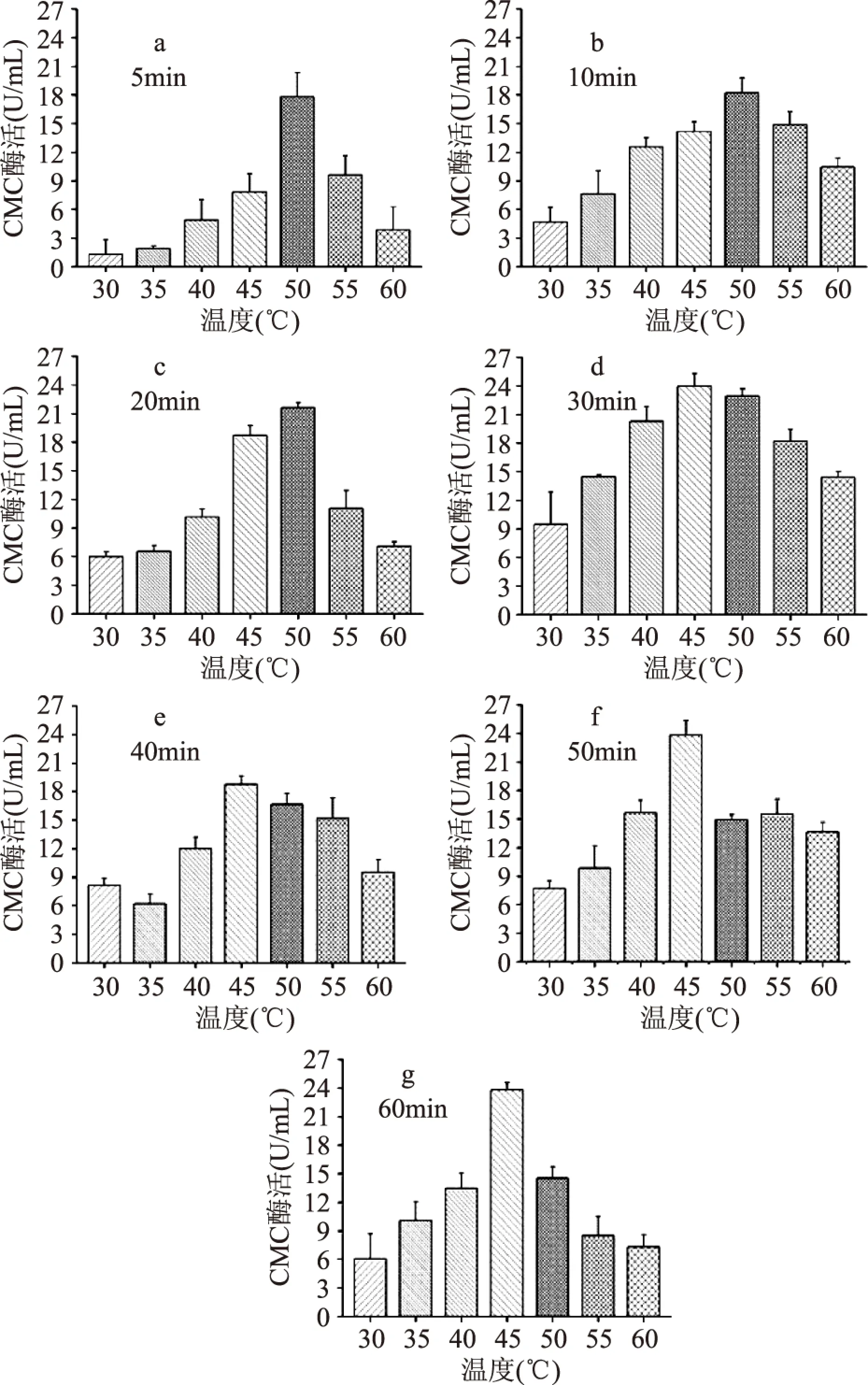
图2 酶促反应时间在不同的温度下对CMC酶活的影响Fig.2 Effect of enzymatic reaction time under different temperature on the determination of CMCase activity
2.3 pH和缓冲液对CMC酶活力的影响
pH对酶活力的影响极为显著,在一定pH的适宜缓冲体系下,酶表现出最大活力,高于或低于此pH,酶活力降低[20]。目前,对于本株灰略红链霉菌的酶学性质尚未有研究报道,在两种缓冲体系下测定的结果如图3所示。

图3 不同缓冲体系的系列pH测定纤维素酶活OD值Fig.3 OD values of cellulose enzyme activity under different buffer systems of a series of pH
从图3可以看出,菌株JSD-1所表达的纤维素酶活力在磷酸缓冲液体系中较高,其中pH为7.0 时测定的酶活力最高,且测定值的平行组之间差异相对较小,高于或低于此条件下测出的酶活力都较低。究其原因可能有两点:不同缓冲体系所含离子对纤维素酶的激活或抑制不同;同一缓冲体系中,不同pH的状态下其底物和分子的带电状态不同,酶的活性部位及底物的解离状态也不同,当酶反应处于最适pH时,酶分子上的活性基团的解离状态最易与底物结合,故在pH7.0的条件下,酶反应速率最高,其他条件下的活性基团解离状态不同程度的发生改变,酶和底物的结合力降低,因而酶反应速度降低。
2.4 底物浓度以及粗酶液和底物添加量对CMC酶活力的影响
根据酶反应动力学原理及参考文献[21-23],同时研究底物浓度以及粗酶液和底物添加量这三个因子的交互作用对酶活力测定的影响,结果如图4所示。
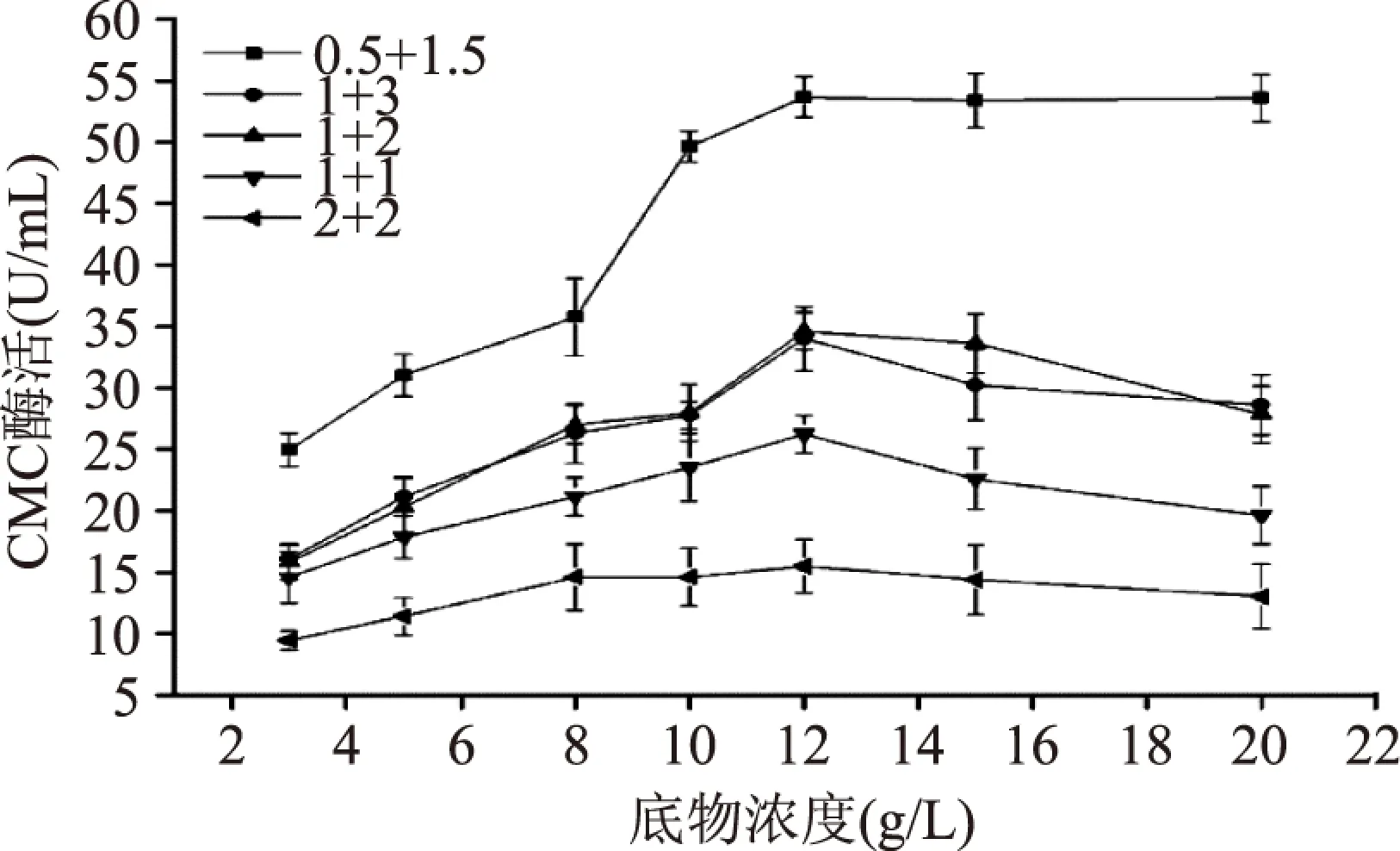
图4 底物浓度以及粗酶量 和底物添加量对CMC酶活的影响Fig.4 The influences on CMCase activity of substrate concentration as well as solvent amount of crude enzyme and substrate
从图4可以看出,纤维素酶活力随着底物浓度的增加而增大,当底物浓度达到12g/L时,酶活表达量达到最大,酶活趋于极限值,底物浓度超过该饱和浓度后酶活反应速率趋于平稳并缓慢下降。原因是底物CMC浓度较低时,酶活反应速率与底物浓度成正相关;当底物浓度恰好被酶所饱和时反应速率达到最大,此后再增加底物浓度酶活力不但不会增加反而会慢慢下降,这是由于高浓度条件下CMC 溶解性较差,反应速率在一定程度上被阻遏,且实验结果的准确性和重复性也较差。
图4还表明粗酶液和底物添加量对酶活测定结果影响显著,可以看出当粗酶液和底物添加量比例为1∶3时酶活力反应速率较高,其中当粗酶液和底物添加量的组合为0.5+1.5mL 时所测得的纤维素酶活力均比其他组合高,当底物浓度为12g/L时其所测的酶活值最高,说明该组合在整个体系反应中饱和率较高。因此,综合分析,酶活检测体系的最适底物浓度为12g/L,最适粗酶液和底物添加量分别为0.5mL和1.5mL。
2.5 DNS显色时间对CMC酶活力的影响
由图5可知,显色时间为1~5min 时,显色反应速率增长较快,在5~12min CMC酶活值基本保持平稳,12min 后反应速率再次上升,并在15min 时出现最大值,比平稳期的平均值提高了14.75%,且其数值稳定性和重复性较高,15min 后呈缓慢下降趋势,这与文献[24]测得的显色时间超过5min 后基本趋于稳定的说法不一致。可能是由于不同来源的纤维素酶酶活不同,产生的还原糖量也不同。因此,采用最高测量值的还原糖标准液来确定显色时间,即显色15min。

图5 系列显色时间对CMC酶活力的影响Fig.5 Influence on CMCase activity of a series of coloration time
2.6 空白(对照)实验对CMC 酶活测定的影响
国际理论和应用化学协会(IU PAC)对纤维素酶酶活测定中的空白实验没有做明确的规定,根据实际测定中选择不同处理方法,这就使得同一酶样品的酶活测定值各异[6]。为了最小程度的降低人为因素造成的误差和提高酶活测定值的可比性,本实验设计了六个不同的空白处理,具体实验设计和结果见图6。
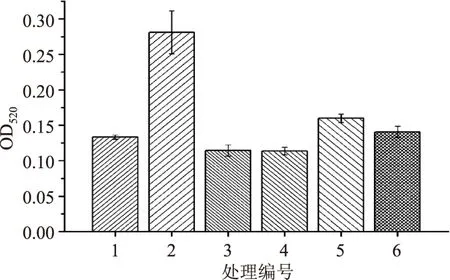
图6 不同空白处理对酶活吸光值的影响Fig.6 Effects on absorbance values of enzyme activity with different blank treatments
由结果可知,JSD-1所产CMC酶活耐高温,100℃处理15min难以将其彻底灭活。处理3和处理4酶活样本灭活比较彻底,且样本误差也较低,但处理4的设计更为合理。空白以底物作为对照,可消除酶活测定的系统误差,其结果更具说服力和代表性。处理2和处理5的空白测定值都偏高,原因是由于酶液和底物混合在缓慢升温过程中已经发生部分反应,尤其是在灭菌锅里升温的时间较长,故酶和底物结合后反应生成较多还原糖。处理5和处理6利用常规的100℃沸水浴灭酶活并不彻底,残留酶活形成的空白值导致测定结果产生较大的负偏差。
有文献[13]研究指出总空白等于底物空白与酶液空白吸光度的算术和,依据此算法总和达到0.18左右,比处理3和处理4高出许多。由以上分析可知,将单独酶液1×105Pa 灭菌的空白处理比较彻底,负偏差<1%,方法操作简便,结果可靠,故最终确定处理4为空白实验体系进行纤维素酶酶活力的空白测定。
2.7 响应面设计(Box-Behnken Design)优化CMC酶活测定条件
根据以上单因素实验结果,选取酶促反应温度(X1)、pH(X2)、反应时间(X3)、底物浓度(X4)和显色时间(X5)五个主要影响因素为自变量,以纤维素酶活力为响应指标,采用Box-Behnken方法设计响应面分析实验,实验设计及实施结果见表5。

表5 Box-Behnken优化实验设计与结果Table 5 Optimizationexperimental design and responses of Box-Behnkendesign
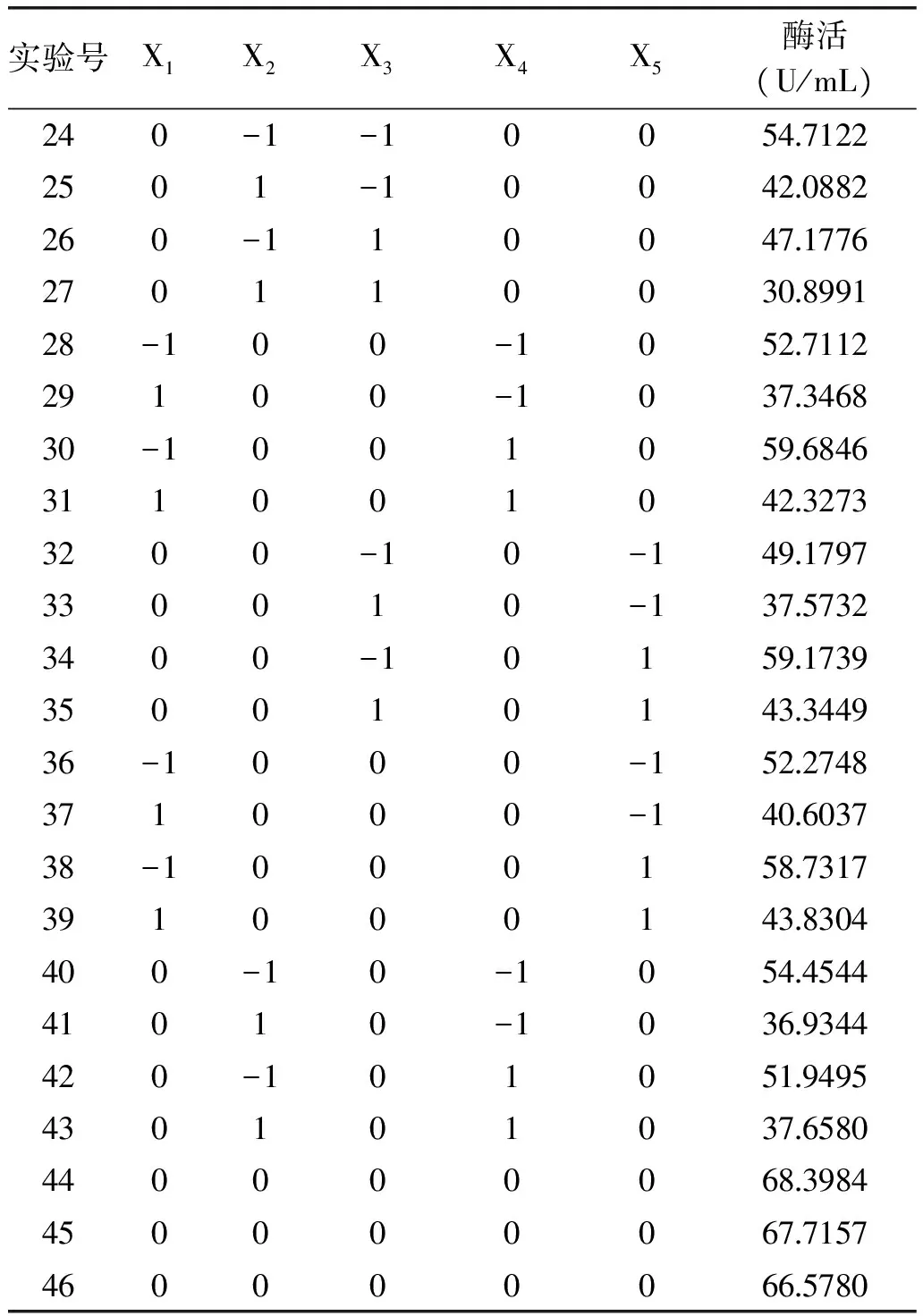
续表
通过采用Statistica 8.0软件分析得到目标响应值与各因素关系的二阶经验模型(如下式所示):Y=67.46240-7.24316X1-7.90046X2-4.88195X3+1.28797X4+2.79076X5-10.05468X12-13.55794X22-10.35951X32-9.66461X42-9.69721X52,该方程系数极显著(Pr>F小于0.01),自动剔除差异非极显著的交叉项。

根据回归模型预测出CMC酶活检测的最高值,即当酶促温度为43℃,pH6.7,酶促时间28min,底物浓度12g/L,显色时间16min时,预测出CMC酶活的最大值为70.7366U/mL,实际测CMC酶活最大值为68.546U/mL,实际值比预测值误差为±3.19%,可见该模型较好的预测了实验结果。
保持其中单一因素最优条件,其他两个因素与响应值关系用响应面立体图如图7所示。
由图7可知,图a、b、c、d 交互作用对酶活影响显著,这与表6中的p值结果一致。
2.8 模型的验证
为证实预测值与真实值之间的拟合程度,进行3次平行验证实验,即在底物浓度为12g/L、pH7.0的磷酸缓冲液、45℃酶促反应30min的条件下,显色15min后于520nm处检测CMC酶活,分别为64.985、65.156和66.509U/mL,实验值略低于模型预测值,实验值与模型估计值最大相差8.1%,二者非常接近,表明预测值与实验值有较好的拟合性,进一步验证了该模型的可信程度。
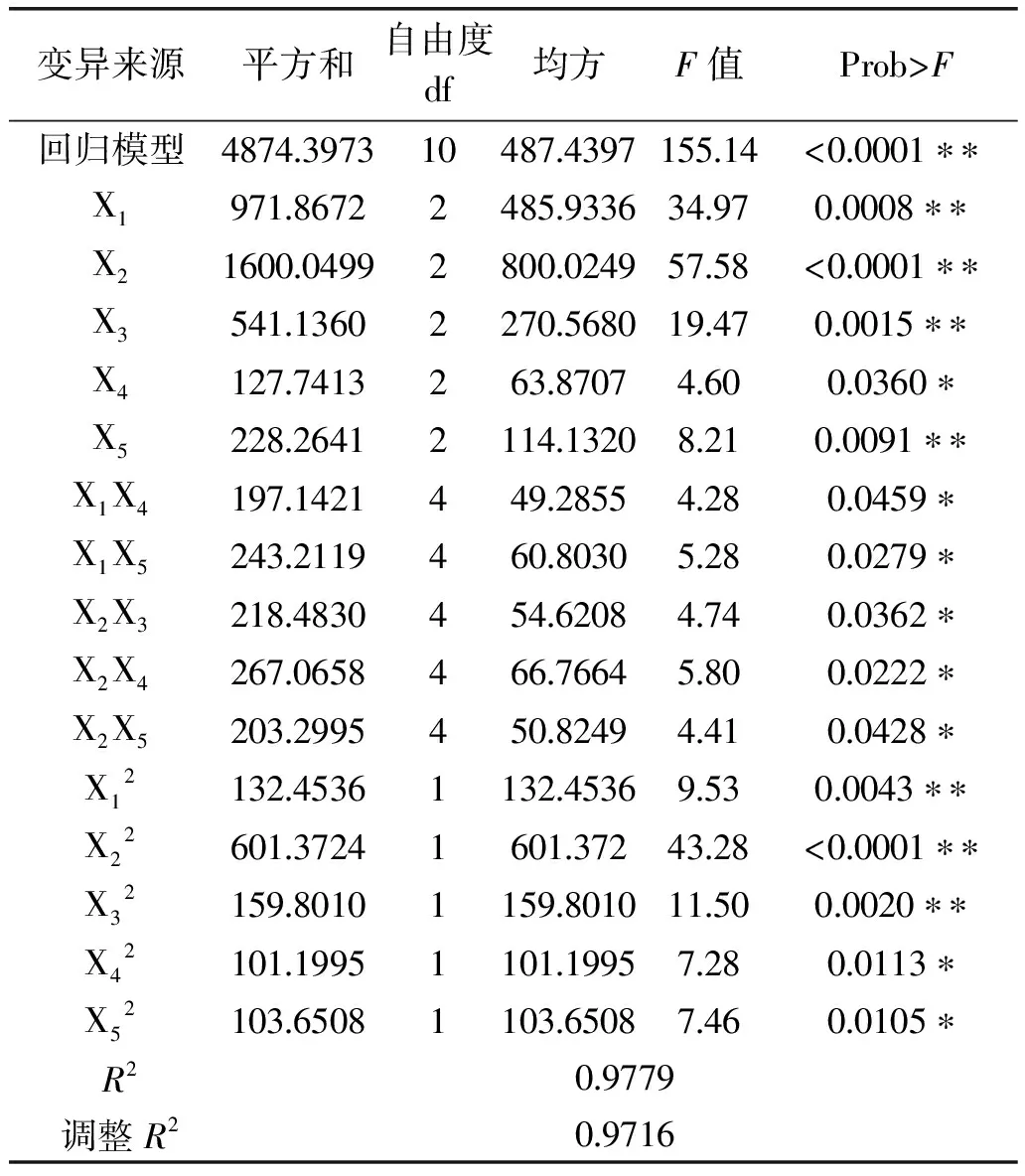
表6 回归与方差分析结果Table 6 Results of regression and variance analysis
注:*差异显著,p<0.05;**差异极显著p<0.01。
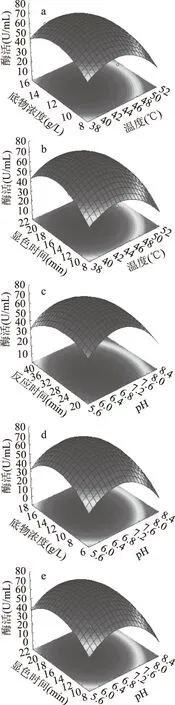
图7 五个因子交互作用对CMC酶活影响的响应面图Fig.7 Response surface of effect of interaction of with five factors on CMCase activity注:a:反应温度和底物浓度交互对CMC酶活的响应; b:温度和显色时间交互对CMC酶活的响应; c:pH和反应时间交互对CMC酶活的响应; d:pH和底物浓度交互对CMC酶活的响应; e:pH和显色时间的交互对CMC酶活的影响。
3 结论
本文通过单因素实验分析了DNS法检测条件对CMC酶活的影响,并结合Box-Behnken 的中心组合设计及响应面分析了DNS法中5个主要影响因子及其交互作用对CMC酶活检测条件的影响。CMC酶活在pH为7.0的磷酸缓冲液、底物浓度为12g/L、45℃酶促反应30min的条件下,显色15min后于520nm处检测最适宜。BBD实验结果显示五个因子的显著性大小依次为pH>反应温度>反应时间>显色时间>底物浓度,pH、反应时间和温度对CMC酶活检测影响较大,在适宜的温度范围内(45~55℃),足够的酶促反应时间可使CMC酶活的表达量较高。因此,在测定不同菌株产纤维素酶活的实验中需对其酶学性质进行分析,然后在一定pH的缓冲体系中进行测定。此外,空白实验酶液1×105Pa处理15min灭活作为纤维素酶活最优空白对照组。
[1]Bhat M K,Bhat S. Cellulose degrading enzymes and their potential industrial applications[J]. Biotechnology advances,1997,15(3):583-620.
[2]王建荣,张曼夫. 绿色木霉纤维素酶CBHII 基因的分子克隆[J]. 真菌学,1994,13(3):235-240
[3]Ragauskas AJ,Williams CK,Davison BH,etal.The path forward for biofuels and biomaterials[J].Science,2006,311(5760):484-489.
[4]王骥. 福寿螺多功能纤维素酶EGX的研究[D],上海:中国科学院研究生院:上海生命科学研究院,2004.
[5]Dashtban M,Maki M,Leung K T,etal. Cellulase activities in biomass conversion:measurement methods and comparison[J]. Critical reviews in biotechnology,2010,30(4):302-309.
[6]Ghose T K. Measurement of cellulase activities[J]. Pure and applied Chemistry,1987,59(2):257-268.
[7]Goksyr J,Eriksen J. Microbial Enzymes and Bioconversions(e d.Hose A. H.),London,NewYork,Torouto,Sydney,San Francisco[J]. 1980,5:301~306
[8]Miller GL,Blum R,Glennon WE. Measurement of carboxymethylcellulase activity[J]. Analytical biochemistry 1960;2:127-32.
[9]Tasun K,Chose P,Ghen K. Sugar determination of DNS method[J]. Biotechnology and Bioengineering,1970,12:921.
[10]Breuil C,Saddler J N. Comparison of the 3,5-dinitrosalicylic acid and Nelson-Somogyi methods of assaying for reducing sugars and determining cellulase activity[J]. Enzyme and microbial technology,1985,7(7):327-332.
[11]王琳,刘国生,王林嵩,等. DNS 法测定纤维素酶活力最适条件研究[J]. 河南师范大学学报:自然科学版,1998,26(3):66-69.
[12]孙盈,田永强,赵丽坤. 纤维素酶的 CMC 酶活测定条件的研究[J]. 食品工业科技,2013,34(2):68-71.
[13]邹水洋,郭祀远. 空白实验及测试条件对纤维素酶活测定的影响[J]. 食品工业科技,2010(6):344-346.
[14]冯海玮,周培,毛亮,等. 一株高效纤维素降解菌的筛选及其产酶条件优化[J]. 上海交通大学学报:农业科学版,2013,31(2):24-29.
[15]Feng H W,Zhi Y E,Shi W W,etal. Isolation,identification and characterization of a straw degrading Streptomyces griseorubens JSD-1[J]. African Journal of Microbiology Research,2013,7(22):2730-2735.
[16]张丽青,吴海龙,姜红霞,等. 纤维素降解细菌的筛选及其产酶条件优化[J]. 环境科学与管理,2007,10(33):110-113.
[17]赵玉萍. 稻壳对绿色木霉产纤维素酶的影响[J]. 中国粮油学报,2006,21(6):18-21.
[18]高培基. 纤维素酶活力测定方法研究进展[J]. 工业微生物,1985,6(5):5-8.
[19]王俊丽,聂国兴,李素贞,等. DNS 法测定还原糖含量时最适波长的确定[J]. 河南农业科学,2010,4(4):115-118.
[20]刘宗林,彭义交,胡阿伟. 一株纤维素酶酶学特性的研究[J]. 食品科学,2003(3):32-35.
[21]B施特马赫. 酶的测定方法[M].钱嘉渊译,北京:中国轻工业出版社,1992:103-107.
[22]姜心,陈伟,周波,等. 纤维素酶活测定影响因素的研究[J]. 食品工业科技,2010(5):65-68.
[23]夏服宝,邱雁临,孙宪迅. 纤维素酶活力测定条件研究[J]. 饲料工业,2006,26(16):23-26.
[24]李兰晓,杜金华,李军训,等. CMC 糖化力法测定纤维素酶活性条件的研究[J]. 饲料工业,2007,27(24):49-52.
Optimization of conditions for CMCase activity of cellulase detected withDNS method forStreptomycesgriseorubens(JSD-1)
LUO Yan-qing,ZHANG Dan,FENG Hai-wei,CHAI Xiao-tong,ZHI Yue-e*,ZHOU Pei
(School of Agriculture and Biology&Bor S. Luh Food Safety Research Center,Shanghai Jiaotong University,Key Laboratory of Urban Agriculture(South),Ministry of Agriculture,Shanghai 200240,China)
In order to study the experimental conditions of 3,5-Dinitrosalicylic acid(DNS)to measure the activity of carboxymethyl cellulase(CMCase),factors including wavelength,enzymatic reaction temperature and time,pH,solvent amount of crude enzyme and substrate,substrate concentration,coloration time and blank experiment were used by a single factor test to explore the influence on the results of CMCase activity. A relatively optimal solution which was applicable to CMCase activity was established by using the method of response surface analysis(RSA)and variance analysis. Results showed that the best conditions for detecting the activity of CMCase was at 520nm wavelength,45℃ and 30min for enzymatic reaction,pH7.0,substrate concentration 12g/L,coloration 15min.
3,5-Dinitrosalicylic acid;response surface;detection conditions for CMCase activity
2014-04-21
罗艳青(1988-),女,硕士研究生,研究方向:农业微生物。
*通讯作者:支月娥(1961-),女,本科,副教授,研究方向:农业微生物。
国家高技术研究发展计划(863计划)课题(2012AA101405);公益性行业(农业)科研专项经费项目(200903056);国家自然科学基金 (31071860)。
TS201.3
A
1002-0306(2015)03-0156-07
10.13386/j.issn1002-0306.2015.03.024
